Unveiling the Significance of Peroxiredoxin 6 in Central Nervous System Disorders

Abstract
:1. Introduction
2. Structure of Prdx6
3. Enzyme Activities and Function of Prdx6
4. Prdx6 Expression in the CNS
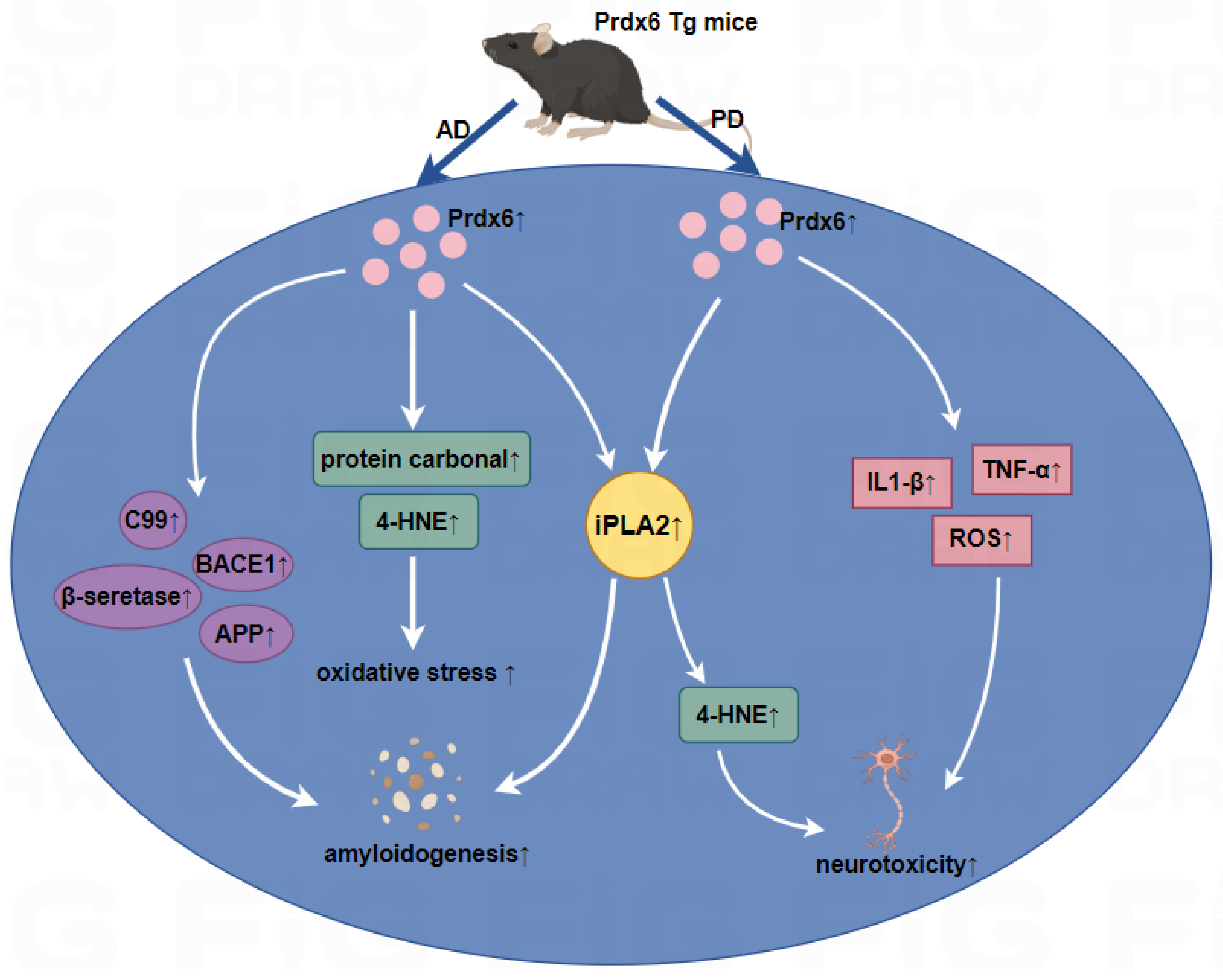
5. Prdx6 and CNS Diseases
5.1. Alzheimer’s Disease (AD)

{kind=link}
{kind=link}
| Tissues | Species | Expression Level | Expression Tissue | Function | Reference |
|---|---|---|---|---|---|
| Postmortem brain | Human | ↑ | midfrontal cortex, cingulate, hippocampus, and amygdala | Prdx6 plays an anti-oxidant role in AD | [43] |
| Postmortem brain (female) | Human | ↑ | superior frontal gyrus | N/A | [46] |
| rpAD postmortem brain | Human | ↓ | hippocampus | N/A | [47] |
| Postmortem brain | Human | no change | frontal cortex and cerebellum | Prdx6 did not show significant changes in the brains of AD patients and possibly has no critical role in cellular defense against oxidative stress. | [50] |
| Postmortem brain | Human | ↑ | hippocampus | The increased expression of Prdx6 in AD was closely related to the degree of oxidative stress. | [51] |
| Tissues | Species | Expression Level | Expression Tissue | Function | Reference |
|---|---|---|---|---|---|
| APP/PS1 Prdx6 Tg female mice | Mice | ↑ | cortex and hippocampus | The upregulation of Prdx6 in AD mice can attenuate Aβ pathology. | [6] |
| 3xTg mice | Mice | no change | hippocampus | Prdx6 was not associated with cumulative oxidative stress in animal models of neurodegenerative disease. | [49] |
| PS2 (N141I) Tg mice | Mice | ↓ | lung | The PS2 mutation inhibits the aiPLA2 activity of Prdx6 through the γ-secretase cleavage mechanism to suppress lung-tumor development. | [53] |
| Aβ1–42-infused Prdx6 transgenic mice | Mice | ↑ | cortex and hippocampus | The overexpression of Prdx6 in AD mice promotes amyloidosis and increases oxidative stress, thereby expediting the progression of AD. | [54] |
| Tau (P301S) Tg mice | Mice | ↑ | anterior horn of the spinal cord | Prdx6 functions as a neuroprotective mechanism against tau toxicity. | [56] |
| Aβ25–35-treated BV2 cells | Mice | ↑ | N/A | Prdx6 is protective against oxidative stress in microglia and synergistically maintains the transition to a chronic neuroinflammatory phenotype, reinforcing the role of Prdx6 in AD | [57] |
| Aβ1–42-infused rat | Rat | ↑ | hippocampus | N/A | [48] |
| Aβ1–42-induced rat primary neuron | Rat | ↓ | N/A | Thiacremonone influences Prdx6 expression levels and oxidative stress, thereby protecting against amyloidosis and memory dysfunction and inhibiting the development and progression of AD. | [55] |
| Aβ25–35-treated rat PC12 cells | Rat | ↑ | N/A | Prdx6 can slow the progression of AD and limit the extent of AD-induced neuronal cell death. | [58] |
5.2. Parkinson’s Disease (PD)
5.3. Cerebral Ischemia
5.4. Spinal Cord Injury (SCI)
5.5. Traumatic Brain Injury (TBI)
5.6. Prion Disease
5.7. Multiple Sclerosis (MS)
5.8. Amyotrophic Lateral Sclerosis (ALS)
5.9. Gliomas
5.10. Epilepsy
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Peshenko, I.V.; Novoselov, V.I.; Evdokimov, V.A.; Nikolaev Yu, V.; Shuvaeva, T.M.; Lipkin, V.M.; Fesenko, E.E. Novel 28-kDa secretory protein from rat olfactory epithelium. FEBS. Lett. 1996, 381, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Shichi, H.; Demar, J.C. Non-selenium glutathione peroxidase without glutathione S-transferase activity from bovine ciliary body. Exp. Eye. Res. 1990, 50, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.W.; Dodia, C.; Feinstein, S.I.; Jain, M.K.; Fisher, A.B. 1-Cys peroxiredoxin, a bifunctional enzyme with glutathione peroxidase and phospholipase A2 activities. J. Biol. Chem. 2000, 275, 28421–28427. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B. Peroxiredoxin 6: A bifunctional enzyme with glutathione peroxidase and phospholipase A2 activities. Antioxid. Redox Signal. 2011, 15, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B.; Dodia, C.; Sorokina, E.M.; Li, H.; Zhou, S.; Raabe, T.; Feinstein, S.I. A novel lysophosphatidylcholine acyl transferase activity is expressed by peroxiredoxin 6. J. Lipid Res. 2016, 57, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Pankiewicz, J.E.; Diaz, J.R.; Martá-Ariza, M.; Lizińczyk, A.M.; Franco, L.A.; Sadowski, M.J. Peroxiredoxin 6 mediates protective function of astrocytes in Aβ proteostasis. Mol. Neurodegener. 2020, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Yeo, I.J.; Park, M.H.; Son, D.J.; Kim, J.Y.; Nam, K.T.; Hyun, B.K.; Kim, S.Y.; Jung, M.H.; Song, M.J.; Chun, H.O.; et al. PRDX6 Inhibits Neurogenesis through Downregulation of WDFY1-Mediated TLR4 Signal. Mol. Neurobiol. 2019, 56, 3132–3144. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, H.; Herojit, K.; Singh, L.R.; Haobam, R.; Fisher, A.B. Structural and Functional Diversity of the Peroxiredoxin 6 Enzyme Family. Antioxid. Redox Signal. 2023. [Google Scholar] [CrossRef] [PubMed]
- Chowhan, R.K.; Rahaman, H.; Singh, L.R. Structural basis of peroxidase catalytic cycle of human Prdx6. Sci. Rep. 2020, 10, 17416. [Google Scholar] [CrossRef] [PubMed]
- Sharapov, M.G.; Goncharov, R.G.; Parfenyuk, S.B.; Glushkova, O.V.; Novoselov, V.I. The Role of Phospholipase Activity of Peroxiredoxin 6 in Its Transmembrane Transport and Protective Properties. Int. J. Mol. Sci. 2022, 23, 15265. [Google Scholar] [CrossRef] [PubMed]
- Shahnaj, S.; Potshangbam, A.M.; Chowhan, R.K.; Parray, Z.A.; Kakchingtabam, P.; Kumari, A.; Islam, A.; Khan, A.; Singh, L.R.; Rahaman, H. The anti-oxidant enzyme, Prdx6 might have cis-acting regulatory sequence(s). Int. J. Biol. Macromol. 2020, 149, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Sureshan, M.; Prabhu, D.; Aruldoss, I.; Saraboji, K. Potential inhibitors for peroxiredoxin 6 of W. bancrofti: A combined study of modelling, structure-based drug design and MD simulation. J. Mol. Graph. Model. 2022, 112, 108115. [Google Scholar] [CrossRef] [PubMed]
- Chowhan, R.K.; Hotumalani, S.; Rahaman, H.; Singh, L.R. pH induced conformational alteration in human peroxiredoxin 6 might be responsible for its resistance against lysosomal pH or high temperature. Sci. Rep. 2021, 11, 9657. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B. Peroxiredoxin 6 in the repair of peroxidized cell membranes and cell signaling. Arch. Biochem. Biophys. 2017, 617, 68–83. [Google Scholar] [CrossRef]
- Li, Y.; Yu, H.; Chen, C.; Li, S.; Zhang, Z.; Xu, H.; Zhu, F.; Liu, J.; Spencer, P.S.; Dai, Z.; et al. Proteomic Profile of Mouse Brain Aging Contributions to Mitochondrial Dysfunction, DNA Oxidative Damage, Loss of Neurotrophic Factor, and Synaptic and Ribosomal Proteins. Oxidative Med. Cell. Longev. 2020, 2020, 5408452. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Chhunchha, B.; Fatma, N.; Kubo, E.; Singh, S.P.; Singh, D.P. Delivery of a protein transduction domain-mediated Prdx6 protein ameliorates oxidative stress-induced injury in human and mouse neuronal cells. Am. J. Physiol. Cell Physiol. 2016, 310, C1–C16. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B. Antioxidants Special Issue: Peroxiredoxin 6 as a Unique Member of the Peroxiredoxin Family. Antioxidants 2019, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Derewenda, Z.S.; Sharp, A.M. News from the interface: The molecular structures of triacylglyceride lipases. Trends Biochem. Sci. 1993, 18, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Paluchova, V.; Cajka, T.; Durand, T.; Vigor, C.; Dodia, C.; Chatterjee, S.; Fisher, A.B.; Kuda, O. The role of peroxiredoxin 6 in biosynthesis of FAHFAs. Free Radic. Biol. Med. 2022, 193, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Lapenna, D. Glutathione and glutathione-dependent enzymes: From biochemistry to gerontology and successful aging. Ageing Res. Rev. 2023, 92, 102066. [Google Scholar] [CrossRef] [PubMed]
- Shahnaj, S.; Chowhan, R.K.; Meetei, P.A.; Kakchingtabam, P.; Herojit Singh, K.; Rajendrakumar Singh, L.; Nongdam, P.; Fisher, A.B.; Rahaman, H. Hyperoxidation of Peroxiredoxin 6 Induces Alteration from Dimeric to Oligomeric State. Antioxidants 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Bidooki, S.H.; Sánchez-Marco, J.; Martínez-Beamonte, R.; Herrero-Continente, T.; Navarro, M.A.; Rodríguez-Yoldi, M.J.; Osada, J. Endoplasmic Reticulum Protein TXNDC5 Interacts with PRDX6 and HSPA9 to Regulate Glutathione Metabolism and Lipid Peroxidation in the Hepatic AML12 Cell Line. Int. J. Mol. Sci. 2023, 24, 17131. [Google Scholar] [CrossRef] [PubMed]
- Villar, S.F.; Ferrer-Sueta, G.; Denicola, A. The multifaceted nature of peroxiredoxins in chemical biology. Curr. Opin. Chem. Biol. 2023, 76, 102355. [Google Scholar] [CrossRef] [PubMed]
- Howie, J.; Tulloch, L.B.; Brown, E.; Reilly, L.; Ashford, F.B.; Kennedy, J.; Wypijewski, K.J.; Aughton, K.L.; Mak, J.K.C.; Shattock, M.J.; et al. Glutathione-dependent depalmitoylation of phospholemman by peroxiredoxin 6. Cell Rep. 2024, 43, 113679. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Sundaresh, C.S.; Feinstein, S.I.; Dodia, C.; Skach, W.R.; Jain, M.K.; Nagase, T.; Seki, N.; Ishikawa, K.; Nomura, N.; et al. Identification of a human cDNA clone for lysosomal type Ca2+-independent phospholipase A2 and properties of the expressed protein. J. Biol. Chem. 1997, 272, 2542–2550. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Dong, C.; Li, B. Anti-Oxidant and Pro-Oxidant Effects of Peroxiredoxin 6: A Potential Target in Respiratory Diseases. Cells 2023, 12, 181. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Dodia, C.; Feinstein, S.I.; Harper, S.; Forman, H.J.; Speicher, D.W.; Fisher, A.B. Oxidation of Peroxiredoxin 6 in the Presence of GSH Increases its Phospholipase A₂ Activity at Cytoplasmic pH. Antioxidants 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B.; Dodia, C.; Chatterjee, S. A Peptide Inhibitor of Peroxiredoxin 6 Phospholipase A(2) Activity Significantly Protects against Lung Injury in a Mouse Model of Ventilator Induced Lung Injury (VILI). Antioxidants 2021, 10, 925. [Google Scholar] [CrossRef] [PubMed]
- Qausain, S.; Khan, F.I.; Khan, M.K.A. Conserved acidic second shell residue modulates the structure, stability and activity of non-seleno human peroxiredoxin 6. Int. J. Biol. Macromol. 2023, 242, 124796. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Benipal, B.; Zhou, S.; Dodia, C.; Chatterjee, S.; Tao, J.Q.; Sorokina, E.M.; Raabe, T.; Feinstein, S.I.; Fisher, A.B. Critical role of peroxiredoxin 6 in the repair of peroxidized cell membranes following oxidative stress. Free Radic. Biol. Med. 2015, 87, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Wahlig, S.; Lovatt, M.; Mehta, J.S. Functional role of peroxiredoxin 6 in the eye. Free Radic. Biol. Med. 2018, 126, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Shi, Y.; Zhang, H.; Zhangyuan, G.; Wang, F.; Li, S.; Chen, C.; Zhang, J.; Wang, H.; Zhang, W.; et al. A TNFα/Miz1-positive feedback loop inhibits mitophagy in hepatocytes and propagates non-alcoholic steatohepatitis. J. Hepatol. 2023, 79, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Shi, J.X.; Shi, S.L.; Liu, F.; Rui, G.; Li, X.; Gao, L.B.; Deng, X.L.; Li, Q.F. Nucleophosmin Regulates Intracellular Oxidative Stress Homeostasis via Antioxidant PRDX6. J. Cell. Biochem. 2017, 118, 4697–4707. [Google Scholar] [CrossRef] [PubMed]
- Daverey, A.; Agrawal, S.K. Curcumin alleviates oxidative stress and mitochondrial dysfunction in astrocytes. Neuroscience 2016, 333, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Chhunchha, B.; Kubo, E.; Singh, D.P. Clock Protein Bmal1 and Nrf2 Cooperatively Control Aging or Oxidative Response and Redox Homeostasis by Regulating Rhythmic Expression of Prdx6. Cells 2020, 9, 1861. [Google Scholar] [CrossRef] [PubMed]
- Chhunchha, B.; Kubo, E.; Singh, D.P. Sulforaphane-Induced Klf9/Prdx6 Axis Acts as a Molecular Switch to Control Redox Signaling and Determines Fate of Cells. Cells 2019, 8, 1159. [Google Scholar] [CrossRef] [PubMed]
- Kubo, E.; Chhunchha, B.; Singh, P.; Sasaki, H.; Singh, D.P. Sulforaphane reactivates cellular antioxidant defense by inducing Nrf2/ARE/Prdx6 activity during aging and oxidative stress. Sci. Rep. 2017, 7, 14130. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.B. The phospholipase A(2) activity of peroxiredoxin 6. J. Lipid Res. 2018, 59, 1132–1147. [Google Scholar] [CrossRef]
- Patel, P.; Chatterjee, S. Peroxiredoxin6 in Endothelial Signaling. Antioxidants 2019, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Goemaere, J.; Knoops, B. Peroxiredoxin distribution in the mouse brain with emphasis on neuronal populations affected in neurodegenerative disorders. J. Comp. Neurol. 2012, 520, 258–280. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.Y.; Kim, H.S.; Kim, E.K.; Choi, J.H. Expression of peroxiredoxin 1, 2, and 6 in the rat brain during perinatal development and in response to dexamethasone. Free Radic. Res. 2012, 46, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, Q.; Wang, J.; Guo, X.; Song, L. Expressions of peroxiredoxin 1, peroxiredoxin 6 and GFAP in human brain astrocytoma and their clinical significance. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med. Univ. 2012, 32, 1255–1259. [Google Scholar]
- Power, J.H.; Asad, S.; Chataway, T.K.; Chegini, F.; Manavis, J.; Temlett, J.A.; Jensen, P.H.; Blumbergs, P.C.; Gai, W.P. Peroxiredoxin 6 in human brain: Molecular forms, cellular distribution and association with Alzheimer’s disease pathology. Acta Neuropathol. 2008, 115, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Park, K.R.; Kim, E.C.; Hong, J.T. PRDX6 controls multiple sclerosis by suppressing inflammation and blood brain barrier disruption. Oncotarget 2015, 6, 20875–20884. [Google Scholar] [CrossRef] [PubMed]
- Ossenkoppele, R.; van der Kant, R.; Hansson, O. Tau biomarkers in Alzheimer’s disease: Towards implementation in clinical practice and trials. Lancet Neurol. 2022, 21, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.T.; Lu, A.; Craessaerts, K.; Pavie, B.; Sala Frigerio, C.; Corthout, N.; Qian, X.; Laláková, J.; Kühnemund, M.; Voytyuk, I.; et al. Spatial Transcriptomics and In Situ Sequencing to Study Alzheimer’s Disease. Cell 2020, 182, 976–991.e919. [Google Scholar] [CrossRef] [PubMed]
- Drummond, E.; Nayak, S.; Faustin, A.; Pires, G.; Hickman, R.A.; Askenazi, M.; Cohen, M.; Haldiman, T.; Kim, C.; Han, X.; et al. Proteomic differences in amyloid plaques in rapidly progressive and sporadic Alzheimer’s disease. Acta Neuropathol. 2017, 133, 933–954. [Google Scholar] [CrossRef] [PubMed]
- Panahzadeh, F.; Mirnasuri, R.; Rahmati, M. Exercise and Syzygium aromaticum reverse memory deficits, apoptosis and mitochondrial dysfunction of the hippocampus in Alzheimer’s disease. J. Ethnopharmacol. 2022, 286, 114871. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, L.; Simon, S.L.R.; Waitt, B.; Knox, J.D. Proteomic Screen of Brain Glycoproteome Reveals Prion Specific Marker of Pathogenesis. Proteomics 2018, 18, 1700296. [Google Scholar] [CrossRef] [PubMed]
- Krapfenbauer, K.; Engidawork, E.; Cairns, N.; Fountoulakis, M.; Lubec, G. Aberrant expression of peroxiredoxin subtypes in neurodegenerative disorders. Brain Res. 2003, 967, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Shults, N.V.; Gychka, S.G.; Harris, B.T.; Suzuki, Y.J. Protein Expression of Angiotensin-Converting Enzyme 2 (ACE2) is Upregulated in Brains with Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 1687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Lai, Y.; Shelat, P.B.; Hu, C.; Sun, G.Y.; Lee, J.C. Phospholipases A2 mediate amyloid-beta peptide-induced mitochondrial dysfunction. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 11111–11119. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Yun, H.M.; Hwang, C.J.; Park, S.I.; Han, S.B.; Hwang, D.Y.; Yoon, D.Y.; Kim, S.; Hong, J.T. Presenilin Mutation Suppresses Lung Tumorigenesis via Inhibition of Peroxiredoxin 6 Activity and Expression. Theranostics 2017, 7, 3624–3637. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Jin, P.; Han, J.Y.; Lee, M.S.; Han, S.B.; Oh, K.W.; Hong, S.H.; Jung, E.Y.; Hong, J.T. Acceleration of the development of Alzheimer’s disease in amyloid beta-infused peroxiredoxin 6 overexpression transgenic mice. Mol. Neurobiol. 2013, 48, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Jin, P.; Park, K.R.; Hwang, J.; Jeong, H.S.; Kim, E.C.; Jung, J.K.; Oh, K.W.; Hwang, B.Y.; Han, S.B.; et al. Thiacremonone Potentiates Anti-Oxidant Effects to Improve Memory Dysfunction in an APP/PS1 Transgenic Mice Model. Mol. Neurobiol. 2016, 53, 2409–2420. [Google Scholar] [CrossRef] [PubMed]
- Yata, K.; Oikawa, S.; Sasaki, R.; Shindo, A.; Yang, R.; Murata, M.; Kanamaru, K.; Tomimoto, H. Astrocytic neuroprotection through induction of cytoprotective molecules; a proteomic analysis of mutant P301S tau-transgenic mouse. Brain Res. 2011, 1410, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Correani, V.; Di Francesco, L.; Cera, I.; Mignogna, G.; Giorgi, A.; Mazzanti, M.; Fumagalli, L.; Fabrizi, C.; Maras, B.; Schininà, M.E. Reversible redox modifications in the microglial proteome challenged by beta amyloid. Mol. Biosyst. 2015, 11, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.K.; Lee, K.J.; Rhee, S.; Seo, S.B.; Pak, J.H. Protective effects of peroxiredoxin 6 overexpression on amyloid β-induced apoptosis in PC12 cells. Free Radic. Res. 2013, 47, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.T. Parkinson’s Disease and Parkinsonism. Am. J. Med. 2019, 132, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J.; Tan, E.K. Parkinson’s disease: Etiopathogenesis and treatment. J. Neurol. Neurosurg. Psychiatry 2020, 91, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, R.P.; Cheng, W.H.; Zhu, J.H. Prioritized brain selenium retention and selenoprotein expression: Nutritional insights into Parkinson’s disease. Mech. Ageing Dev. 2019, 180, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Power, J.H.; Shannon, J.M.; Blumbergs, P.C.; Gai, W.P. Nonselenium glutathione peroxidase in human brain: Elevated levels in Parkinson’s disease and dementia with lewy bodies. Am. J. Pathol. 2002, 161, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Connelly, E.M.; Frankel, K.S.; Shaw, G.S. Parkin and mitochondrial signalling. Cell. Signal. 2023, 106, 110631. [Google Scholar] [CrossRef]
- Tariq, M.; Khan, H.A.; Al Moutaery, K.; Al Deeb, S. Protective effect of quinacrine on striatal dopamine levels in 6-OHDA and MPTP models of Parkinsonism in rodents. Brain Res. Bull. 2001, 54, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Choi, D.Y.; Oh, K.W.; Hong, J.T. PRDX6 Exacerbates Dopaminergic Neurodegeneration in a MPTP Mouse Model of Parkinson’s Disease. Mol. Neurobiol. 2015, 52, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase A2 in the central nervous system: Implications for neurodegenerative diseases. J. Lipid Res. 2004, 45, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Zhang, X.; Zheng, L.; Li, Z.; Zhao, X.; Lai, W.; Shen, H.; Lv, J.; Yang, G.; Wang, Q.; et al. Peroxiredoxin 6 Is a Crucial Factor in the Initial Step of Mitochondrial Clearance and Is Upstream of the PINK1-Parkin Pathway. Antioxid. Redox Signal. 2016, 24, 486–501. [Google Scholar] [CrossRef]
- Davison, E.J.; Pennington, K.; Hung, C.C.; Peng, J.; Rafiq, R.; Ostareck-Lederer, A.; Ostareck, D.H.; Ardley, H.C.; Banks, R.E.; Robinson, P.A. Proteomic analysis of increased Parkin expression and its interactants provides evidence for a role in modulation of mitochondrial function. Proteomics 2009, 9, 4284–4297. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, P.B. The global burden of stroke: Persistent and disabling. Lancet Neurol. 2019, 18, 417–418. [Google Scholar] [CrossRef]
- Ma, R.; Xie, Q.; Li, Y.; Chen, Z.; Ren, M.; Chen, H.; Li, H.; Li, J.; Wang, J. Animal models of cerebral ischemia: A review. Biomed. Pharmacother. 2020, 131, 110686. [Google Scholar] [CrossRef]
- Choi, T.M.; Yun, M.; Lee, J.K.; Park, J.T.; Park, M.S.; Kim, H.S. Proteomic Analysis of a Rat Cerebral Ischemic Injury Model after Human Cerebral Endothelial Cell Transplantation. J. Korean Neurosurg. Soc. 2016, 59, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Sidyakin, A.A.; Kaysheva, A.L.; Kopylov, A.T.; Lobanov, A.V.; Morozov, S.G. Proteomic Analysis of Cerebral Cortex Extracts from Sus scrofa with Induced Hemorrhagic Stroke. J. Mol. Neurosci. MN 2018, 65, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yeung, P.K.; McAlonan, G.M.; Chung, S.S.; Chung, S.K. Transgenic mice over-expressing endothelial endothelin-1 show cognitive deficit with blood-brain barrier breakdown after transient ischemia with long-term reperfusion. Neurobiol. Learn. Mem. 2013, 101, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Ni, W.; Zhou, J.; Ling, Y.; Niu, D.; Lu, X.; Chen, T.; Ramalingam, M.; Hu, J. Peroxiredoxin 6 secreted by Schwann-like cells protects neuron against ischemic stroke in rats via PTEN/PI3K/AKT pathway. Tissue Cell 2021, 73, 101635. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Liu, H.; Zhou, J.; Liu, Z.; Yang, Y.; Peng, Y.; You, H.; Yang, D.; Xie, P. Ipsilateral hippocampal proteomics reveals mitochondrial antioxidative stress impairment in cortical-lesioned chronic mild stressed rats. Curr. Mol. Med. 2014, 14, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Tan, B.; Ma, J.; Zhang, L.; Jin, X.; Li, C. Prdx6 Upregulation by Curcumin Attenuates Ischemic Oxidative Damage via SP1 in Rats after Stroke. BioMed Res. Int. 2017, 2017, 6597401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ma, Y.; Zhao, Y.; Guo, N.; Han, C.; Wu, Q.; Mu, C.; Zhang, Y.; Tan, S.; Zhang, J.; et al. Systematic review of melatonin in cerebral ischemia-reperfusion injury: Critical role and therapeutic opportunities. Front. Pharmacol. 2024, 15, 1356112. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.H.; Cho, E.H.; Kim, M.O.; Koh, P.O. Identification of proteins differentially expressed by melatonin treatment in cerebral ischemic injury—A proteomics approach. J. Pineal Res. 2009, 46, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Ji, Y.; Li, Y.; You, Y.; Zhou, Y. PRDX6-iPLA2 aggravates neuroinflammation after ischemic stroke via regulating astrocytes-induced M1 microglia. Cell Commun. Signal. CCS 2024, 22, 76. [Google Scholar] [CrossRef] [PubMed]
- Shanshan, Y.; Beibei, J.; Li, T.; Minna, G.; Shipeng, L.; Li, P.; Yong, Z. Phospholipase A2 of Peroxiredoxin 6 Plays a Critical Role in Cerebral Ischemia/Reperfusion Inflammatory Injury. Front. Cell. Neurosci. 2017, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Zhou, Y.; Peng, L.; Wu, X.; Li, Y.; Li, Y.; Zhao, Y. Knocking Down Peroxiredoxin 6 Aggravates Cerebral Ischemia-Reperfusion Injury by Enhancing Mitophagy. Neuroscience 2022, 482, 30–42. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Dai, R.; Zhou, X.; Li, X.; Song, X.; Yan, H.; Meng, Q.; Yang, C.; Lin, Q. Protective effect of 4-Methoxy benzyl alcohol on the neurovascular unit after cerebral ischemia reperfusion injury. Biomed. Pharmacother. 2019, 118, 109260. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, J.; Wang, X.; Lei, S.; Wu, X.; Chen, Y.; Wu, J.; Zhao, Y. 4-Hydroxybenzyl alcohol confers neuroprotection through up-regulation of antioxidant protein expression. Neurochem. Res. 2013, 38, 1501–1516. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Tian, L.; Liu, J.; Wu, Q.; Wang, N.; Wang, G.; Wang, Y.; Seto, S. Ligustilide ameliorates hippocampal neuronal injury after cerebral ischemia reperfusion through activating PINK1/Parkin-dependent mitophagy. Phytomed. Int. J. Phytother. Phytopharm. 2022, 101, 154111. [Google Scholar] [CrossRef] [PubMed]
- Shichita, T.; Hasegawa, E.; Kimura, A.; Morita, R.; Sakaguchi, R.; Takada, I.; Sekiya, T.; Ooboshi, H.; Kitazono, T.; Yanagawa, T.; et al. Peroxiredoxin family proteins are key initiators of post-ischemic inflammation in the brain. Nat. Med. 2012, 18, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Tang, P.; Wang, J.; Ye, W.; Ge, X.; Rong, Y.; Ji, C.; Wang, Z.; Bai, J.; Fan, J.; et al. Extracellular vesicles derived from melatonin-preconditioned mesenchymal stem cells containing USP29 repair traumatic spinal cord injury by stabilizing NRF2. J. Pineal Res. 2021, 71, e12769. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, M.E.; Filous, A.R.; Vadala, C.P.; Webb, A.; Pietrzak, M.; Sahenk, Z.; Prüss, H.; Reiser, P.J.; Popovich, P.G.; Arnold, W.D.; et al. Lesion level-dependent systemic muscle wasting after spinal cord injury is mediated by glucocorticoid signaling in mice. Sci. Transl. Med. 2023, 15, eadh2156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.L.; Gao, X.; Jiang, D.; Zhong, Z.Q.; Zeng, X.; Rao, Y.; Hu, X.; Li, T.Z.; Li, X.J.; et al. Lentivirus-mediated inhibition of tumour necrosis factor-α improves motor function associated with PRDX6 in spinal cord contusion rats. Sci. Rep. 2015, 5, 8486. [Google Scholar] [CrossRef] [PubMed]
- Daverey, A.; Agrawal, S.K. Regulation of Prdx6 by Nrf2 Mediated Through aiPLA2 in White Matter Reperfusion Injury. Mol. Neurobiol. 2021, 58, 1275–1289. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Sharma, S. Recent Advances in Pathophysiology of Traumatic Brain Injury. Curr. Neuropharmacol. 2018, 16, 1224–1238. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pacheco, V.; Vargas-Medrano, J.; Tran, E.; Nicolas, M.; Price, D.; Patel, R.; Tonarelli, S.; Gadad, B.S. Prognosis and Diagnostic Biomarkers of Mild Traumatic Brain Injury: Current Status and Future Prospects. J. Alzheimer’s Dis. JAD 2022, 86, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Di Battista, A.P.; Buonora, J.E.; Rhind, S.G.; Hutchison, M.G.; Baker, A.J.; Rizoli, S.B.; Diaz-Arrastia, R.; Mueller, G.P. Blood Biomarkers in Moderate-To-Severe Traumatic Brain Injury: Potential Utility of a Multi-Marker Approach in Characterizing Outcome. Front. Neurol. 2015, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Buonora, J.E.; Yarnell, A.M.; Lazarus, R.C.; Mousseau, M.; Latour, L.L.; Rizoli, S.B.; Baker, A.J.; Rhind, S.G.; Diaz-Arrastia, R.; Mueller, G.P. Multivariate analysis of traumatic brain injury: Development of an assessment score. Front. Neurol. 2015, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Manevich, Y.; Hutchens, S.; Halushka, P.V.; Tew, K.D.; Townsend, D.M.; Jauch, E.C.; Borg, K. Peroxiredoxin VI oxidation in cerebrospinal fluid correlates with traumatic brain injury outcome. Free Radic. Biol. Med. 2014, 72, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Abu Hamdeh, S.; Shevchenko, G.; Mi, J.; Musunuri, S.; Bergquist, J.; Marklund, N. Proteomic differences between focal and diffuse traumatic brain injury in human brain tissue. Sci. Rep. 2018, 8, 6807. [Google Scholar] [CrossRef] [PubMed]
- Di Battista, A.P.; Churchill, N.; Schweizer, T.A.; Rhind, S.G.; Richards, D.; Baker, A.J.; Hutchison, M.G. Blood biomarkers are associated with brain function and blood flow following sport concussion. J. Neuroimmunol. 2018, 319, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Buonora, J.E.; Mousseau, M.; Jacobowitz, D.M.; Lazarus, R.C.; Yarnell, A.M.; Olsen, C.H.; Pollard, H.B.; Diaz-Arrastia, R.; Latour, L.; Mueller, G.P. Autoimmune Profiling Reveals Peroxiredoxin 6 as a Candidate Traumatic Brain Injury Biomarker. J. Neurotrauma 2015, 32, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Orge, L.; Lima, C.; Machado, C.; Tavares, P.; Mendonça, P.; Carvalho, P.; Silva, J.; Pinto, M.L.; Bastos, E.; Pereira, J.C.; et al. Neuropathology of Animal Prion Diseases. Biomolecules 2021, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Piñar-Morales, R.; Barrero-Hernández, F.; Aliaga-Martínez, L. Human prion diseases: An overview. Med. Clin. 2023, 160, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Ramosaço, E.; Bajrami, N.; Vyshka, G. A Theoretical Framework on the Biology of Prion Diseases. Acta Inform. Med. 2023, 31, 141–145. [Google Scholar] [PubMed]
- López-Pérez, Ó.; Badiola, J.J.; Bolea, R.; Ferrer, I.; Llorens, F.; Martín-Burriel, I. An Update on Autophagy in Prion Diseases. Front. Bioeng. Biotechnol. 2020, 8, 975. [Google Scholar] [CrossRef] [PubMed]
- Piconi, G.; Peden, A.H.; Barria, M.A.; Green, A.J.E. Epitope mapping of the protease resistant products of RT-QuIC does not allow the discrimination of sCJD subtypes. PLoS ONE 2019, 14, e0218509. [Google Scholar] [CrossRef] [PubMed]
- Asuni, A.A.; Guridi, M.; Sanchez, S.; Sadowski, M.J. Antioxidant peroxiredoxin 6 protein rescues toxicity due to oxidative stress and cellular hypoxia in vitro, and attenuates prion-related pathology in vivo. Neurochem. Int. 2015, 90, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Reuter, A.; Hüller, P.; Löwer, J.; Wessler, S. Peroxiredoxin 6 promotes upregulation of the prion protein (PrP) in neuronal cells of prion-infected mice. Cell Commun. Signal. CCS 2012, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.; Antel, J.; Kuhlmann, T. Inflammation in multiple sclerosis: Consequences for remyelination and disease progression. Nat. Rev. Neurol. 2023, 19, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, J.F.; Hoffman, B.M.; Tyor, W.R. CNS inflammatory demyelinating disorders: MS, NMOSD and MOG antibody associated disease. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2020, 68, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Uzawa, A.; Mori, M.; Masuda, H.; Ohtani, R.; Uchida, T.; Aoki, R.; Kuwabara, S. Peroxiredoxins are involved in the pathogenesis of multiple sclerosis and neuromyelitis optica spectrum disorder. Clin. Exp. Immunol. 2020, 202, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Feldman, E.L.; Goutman, S.A.; Petri, S.; Mazzini, L.; Savelieff, M.G.; Shaw, P.J.; Sobue, G. Amyotrophic lateral sclerosis. Lancet 2022, 400, 1363–1380. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Jagaraj, C.J.; Parakh, S.; Atkin, J.D. Emerging Evidence Highlighting the Importance of Redox Dysregulation in the Pathogenesis of Amyotrophic Lateral Sclerosis (ALS). Front. Cell. Neurosci. 2020, 14, 581950. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, S.; Äijö, T.; Vickovic, S.; Braine, C.; Kang, K.; Mollbrink, A.; Fagegaltier, D.; Andrusivová, Ž.; Saarenpää, S.; Saiz-Castro, G.; et al. Spatiotemporal dynamics of molecular pathology in amyotrophic lateral sclerosis. Science 2019, 364, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Tang, L.; Li, X.; Fan, F.; Liu, Z. Immunotherapy for glioma: Current management and future application. Cancer Lett. 2020, 476, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Kim, J.H.; Moon, M.J.; Jung, J.H.; Lim, S.Y.; Park, S.H.; Kim, J.H.; Kim, D.G.; Jung, H.W.; Cho, B.K.; et al. Investigation of molecular factors associated with malignant transformation of oligodendroglioma by proteomic study of a single case of rapid tumor progression. J. Cancer Res. Clin. Oncol. 2008, 134, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Gollapalli, K.; Ghantasala, S.; Atak, A.; Rapole, S.; Moiyadi, A.; Epari, S.; Srivastava, S. Tissue Proteome Analysis of Different Grades of Human Gliomas Provides Major Cues for Glioma Pathogenesis. Omics J. Integr. Biol. 2017, 21, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zhang, Y.; Chen, X.; Zhang, J. The Roles of Peroxiredoxin 6 in Brain Diseases. Mol. Neurobiol. 2021, 58, 4348–4364. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, M. Comprehensive analysis of the expression levels and prognostic values of PRDX family genes in glioma. Neurochem. Int. 2022, 153, 105256. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Czech, T.; Felizardo, M.; Baumgartner, C.; Lubec, G. Aberrant expression of cytoskeleton proteins in hippocampus from patients with mesial temporal lobe epilepsy. Amino Acids 2006, 30, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, X.; Liu, S.; Liu, Y.; Yang, Y.; Yang, H.; Chen, Y.; Chen, L. Differentially expressed proteins underlying childhood cortical dysplasia with epilepsy identified by iTRAQ proteomic profiling. PLoS ONE 2017, 12, e0172214. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.J.; Lee, C.; Kim, J.; Shin, H.S.; Yu, M.H. Proteomic analysis of stargazer mutant mouse neuronal proteins involved in absence seizure. J. Neurochem. 2008, 104, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, D.S.; Kang, T.C. Sp1-Mediated Prdx6 Upregulation Leads to Clasmatodendrosis by Increasing Its aiPLA2 Activity in the CA1 Astrocytes in Chronic Epilepsy Rats. Antioxidants 2022, 11, 1883. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Park, H.; Kang, T.C. Peroxiredoxin 6 Regulates Glutathione Peroxidase 1-Medited Glutamine Synthase Preservation in the Hippocampus of Chronic Epilepsy Rats. Antioxidants 2023, 12, 156. [Google Scholar] [CrossRef]


| Tissues | Species | Expression Level | Expression Tissue | Function | Reference |
|---|---|---|---|---|---|
| Postmortem brain | Human | ↑ | gray and white matter | Prdx6 was upregulated in certain cells to respond to oxidative stress in PD. | [62] |
| HeLa cells with CCCP-induced GFP-Parkin overexpression | Human | ↑ | N/A | Prdx6 controls ROS homeostasis during the initial phase of PINK1-Parkin-mediated mitotic phagocytosis. | [67] |
| HEK293 cells with inducible Parkin expression | Human | ↓ | N/A | Prdx6 is a potential substrate of the Parkin. | [68] |
| Tissues | Species | Expression Level | Expression Tissue | Function | Reference |
|---|---|---|---|---|---|
| Parkin−/− mice | mice | ↓ | ventral midbrain | N/A | [63] |
| MPTP-induced mice and 6-OHDA-induced rats | mice/rats | ↑ | striatum | aiPLA2 inhibitor QNC protects against 6-OHDA and MPTP-induced dopaminergic neurotoxicity. | [64] |
| Prdx6 Tg mice in MPTP administration | mice | ↑ | substantia nigra and striatum | The aiPLA2 activity of Prdx6 was increased after MPTP administration in the Prdx6 transgenic mice, which results in a greater loss of dopaminergic neurons and increased behavioral damage. | [65] |
| Models | Species | Expression Level | Expression Tissue | Function | Reference |
|---|---|---|---|---|---|
| TET-1 MCAO | mouse | ↑ | hippocampus | In TET-1 mice perfused for 7 days after 30 min of MCAO, an increased expression of perivascular Prdx6 in the hippocampus may lead to neuronal apoptosis, glial activation, and blood–brain barrier disruption. | [73] |
| Tlr2−/−; Tlr4−/− MCAO | mouse | ↑ | N/A | Prdx6 promotes neuronal cell death by activating Toll-like receptor 2 (TLR2) and TLR4 and inducing macrophages to express inflammatory cytokines, including IL-23. | [80] |
| MCAO | rat | ↑ | N/A | The MCAO model induced an abnormal increased expression of Prdx6, but after human brain endothelial cell transplantation, Prdx6 levels in the MCAO models decreased. | [71] |
| Stroke with heat-induced brain injury in the left anterior cortex | rat | ↑ | hippocampus | A sustained upregulation of Prdx6 expression may help protect hippocampal neurons from oxidative stress in a rat model of stroke (localized heat-induced brain injury in the left anterior cortical tectum). | [75] |
| MCAO | rat | ↑ | peri-infarct cortex | Prdx6 is involved in the inhibition of curcumin-induced oxidative stress during I/R, and the upregulation of Prdx6 by curcumin attenuates ischemic oxidative damage via SP1 in post-stroke rats. | [76] |
| MCAO | rat | ↓ | cerebral cortex | In ischemic brain injury, Prdx6 was increased with a melatonin treatment to protect neuronal cells from ischemic damage. | [78] |
| MCAO | rat | ↑ | cortex | Prdx6 may be an important target for immunomodulation and neuroinflammation after ischemic stroke. Prdx6 released from ischemic cells acts as an endogenous ligand for TLR4 and initiates destructive immune responses in ischemic brains. | [79] |
| OGD/R and MCAO | rat | ↑ | cortex | The aiPLA2 activity of Prdx6 may play a key role in cerebral ischemia/reperfusion injury by regulating TLR2/4, which induces the production of NF-κB, iNOS, and COX-2, and promote neuroinflammation. | [80] |
| MCAO | rat | ↑ | cerebral cortex | Prdx6 knockdown exacerbates cerebral ischemia-reperfusion injury by enhancing PINK1/PARKIN pathway-mediated mitophagy, an effect that increases neuronal apoptosis. | [81] |
| OGD/R and MCAO | rat | ↑ | cortex | Prdx6 is upregulated by 4-HBA to protect neurons from cerebral ischemic injury, possibly through the PI3K/Akt pathway. | [83] |
| Cutting the inner capsule of pig tissue | pig | ↑ | N/A | Prdx6 is involved in neuroprotective mechanisms after stroke, such as compensating for oxidative stress. | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, M.; Huang, X.; Zhu, T.; Zhang, L.; Yang, H.; Shen, Y.; Feng, L. Unveiling the Significance of Peroxiredoxin 6 in Central Nervous System Disorders. Antioxidants 2024, 13, 449. https://doi.org/10.3390/antiox13040449
Xue M, Huang X, Zhu T, Zhang L, Yang H, Shen Y, Feng L. Unveiling the Significance of Peroxiredoxin 6 in Central Nervous System Disorders. Antioxidants. 2024; 13(4):449. https://doi.org/10.3390/antiox13040449
Chicago/Turabian StyleXue, Min, Xiaojie Huang, Tong Zhu, Lijun Zhang, Hao Yang, Yuxian Shen, and Lijie Feng. 2024. "Unveiling the Significance of Peroxiredoxin 6 in Central Nervous System Disorders" Antioxidants 13, no. 4: 449. https://doi.org/10.3390/antiox13040449