

Impact of Maternal Environment and Inflammation on Fetal Neurodevelopment

Abstract
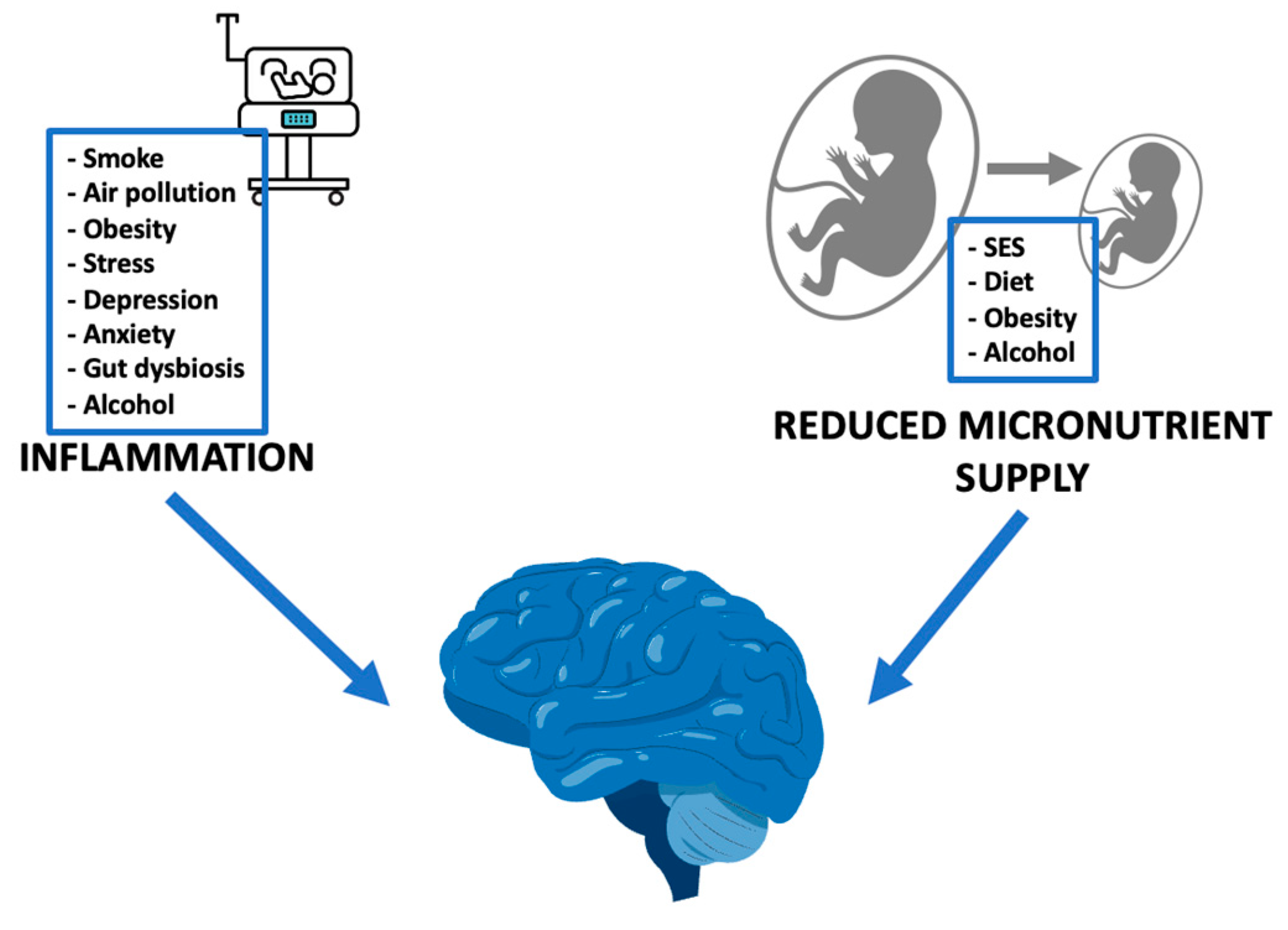
1. Introduction
2. Environment, Lifestyle, and Specific Maternal Phenotypes and Risk Factors
2.1. Maternal Nutrition
2.2. Obesity
2.3. Depression, Anxiety, and Stress
2.4. Smoking
2.5. Alcohol
2.6. Air Pollution
2.7. Socioeconomic Status
2.8. Gut Microbiota
3. Inflammation and Altered Nutrient Supply: Models of Intrauterine Health Programming
3.1. Fetal Growth Restriction: Model of Reduced Nutrient Supply
3.2. Preterm Born Children: Model of Intrauterine Inflammation
3.3. Inflammation
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heland, S.; Fields, N.; Ellery, S.J.; Fahey, M.; Palmer, K.R. The role of nutrients in human neurodevelopment and their potential to prevent neurodevelopmental adversity. Front. Nutr. 2022, 9, 992120. [Google Scholar] [CrossRef]
- Pardi, G.; Cetin, I. Human fetal growth and organ development: 50 years of discoveries. Am. J. Obstet. Gynecol. 2006, 194, 1088–1099. [Google Scholar] [CrossRef]
- Diaz, A.L.; Gleeson, J.G. The Molecular and Genetic Mechanisms of Neocortex Development. Clin. Perinatol. 2009, 36, 503–512. [Google Scholar] [CrossRef]
- Barker, D.J.P.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in england and wales. Lancet 1986, 327, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; et al. Origins of lifetime health around the time of conception: Causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Vasistha, N.A.; Khodosevich, K. The impact of (ab)normal maternal environment on cortical development. Prog. Neurobiol. 2021, 202, 102054. [Google Scholar] [CrossRef] [PubMed]
- Hertz-Picciotto, I.; Korrick, S.A.; Ladd-Acosta, C.; Karagas, M.R.; Lyall, K.; Schmidt, R.J.; Dunlop, A.L.; Croen, L.A.; Dabelea, D.; Daniels, J.L.; et al. Maternal tobacco smoking and offspring autism spectrum disorder or traits in ECHO cohorts. Autism Res. 2022, 15, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Nakaki, A.; Crovetto, F.; Urru, A.; Piella, G.; Borras, R.; Comte, V.; Vellvé, K.; Paules, C.; Segalés, L.; Dacal, M.; et al. Effects of Mediterranean diet or mindfulness-based stress reduction on fetal and neonatal brain development: A secondary analysis of a randomized clinical trial. Am. J. Obstet. Gynecol. MFM 2023, 5, 101188. [Google Scholar] [CrossRef]
- Ogundipe, E.; Tusor, N.; Wang, Y.; Johnson, M.; Edwards, A.; Crawford, M. Randomized controlled trial of brain specific fatty acid supplementation in pregnant women increases brain volumes on MRI scans of their newborn infants. Prostaglandins, Leukot. Essent. Fat. Acids 2018, 138, 6–13. [Google Scholar] [CrossRef]
- Rifkin-Graboi, A.; Bai, J.; Chen, H.; Hameed, W.B.; Sim, L.W.; Tint, M.T.; Leutscher-Broekman, B.; Chong, Y.-S.; Gluckman, P.D.; Fortier, M.V.; et al. Prenatal Maternal Depression Associates with Microstructure of Right Amygdala in Neonates at Birth. Biol. Psychiatry 2013, 74, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Lebel, C.; Walton, M.; Letourneau, N.; Giesbrecht, G.F.; Kaplan, B.J.; Dewey, D. Prepartum and Postpartum Maternal Depressive Symptoms Are Related to Children’s Brain Structure in Preschool. Biol. Psychiatry 2016, 80, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lu, Y.-C.; Jacobs, M.; Pradhan, S.; Kapse, K.; Zhao, L.; Niforatos-Andescavage, N.; Vezina, G.; du Plessis, A.J.; Limperopoulos, C. Association of Prenatal Maternal Psychological Distress with Fetal Brain Growth, Metabolism, and Cortical Maturation. JAMA Netw. Open 2020, 3, e1919940. [Google Scholar] [CrossRef]
- Jacobson, S.W.; Jacobson, J.L.; Molteno, C.D.; Warton, C.M.; Wintermark, P.; Hoyme, H.E.; De Jong, G.; Taylor, P.; Warton, F.; Lindinger, N.M.; et al. Heavy Prenatal Alcohol Exposure is Related to Smaller Corpus Callosum in Newborn MRI Scans. Alcohol. Clin. Exp. Res. 2017, 41, 965–975. [Google Scholar] [CrossRef]
- Roos, A.; Wedderburn, C.J.; Fouche, J.-P.; Subramoney, S.; Joshi, S.H.; Woods, R.P.; Zar, H.J.; Narr, K.L.; Stein, D.J.; Donald, K.A. Central white matter integrity alterations in 2-3-year-old children following prenatal alcohol exposure. Drug Alcohol Depend. 2021, 225, 108826. [Google Scholar] [CrossRef] [PubMed]
- Çetin, Ç.; Bakar, R.Z.; Takmaz, T.; Pasin, Ö.; Kütük, M.S. The effects of maternal smoking on fetal cranial development. Findings from routine midtrimester sonographic anomaly screening. J. Obstet. Gynaecol. 2023, 43, 2176205. [Google Scholar] [CrossRef]
- Peterson, B.S.; Rauh, V.A.; Bansal, R.; Hao, X.; Toth, Z.; Nati, G.; Walsh, K.; Miller, R.L.; Arias, F.; Semanek, D.; et al. Effects of Prenatal Exposure to Air Pollutants (Polycyclic Aromatic Hydrocarbons) on the Development of Brain White Matter, Cognition, and Behavior in Later Childhood. JAMA Psychiatry 2015, 72, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Triplett, R.L.; Lean, R.E.; Parikh, A.; Miller, J.P.; Alexopoulos, D.; Kaplan, S.; Meyer, D.; Adamson, C.; Smyser, T.A.; Rogers, C.E.; et al. Association of Prenatal Exposure to Early-Life Adversity with Neonatal Brain Volumes at Birth. JAMA Netw. Open 2022, 5, e227045. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Kapse, K.; Andersen, N.; Quistorff, J.; Lopez, C.; Fry, A.; Cheng, J.; Andescavage, N.; Wu, Y.; Espinosa, K.; et al. Association Between Socioeconomic Status and In Utero Fetal Brain Development. JAMA Netw. Open 2021, 4, e213526. [Google Scholar] [CrossRef] [PubMed]
- Herzberg, M.P.; Triplett, R.; McCarthy, R.; Kaplan, S.; Alexopoulos, D.; Meyer, D.; Arora, J.; Miller, J.P.; Smyser, T.A.; Herzog, E.D.; et al. The Association Between Maternal Cortisol and Infant Amygdala Volume Is Moderated by Socioeconomic Status. Biol. Psychiatry Glob. Open Sci. 2023, 3, 837–846. [Google Scholar] [CrossRef]
- Belluscio, L.M.; Berardino, B.G.; Ferroni, N.M.; Ceruti, J.M.; Cánepa, E.T. Early protein malnutrition negatively impacts physical growth and neurological reflexes and evokes anxiety and depressive-like behaviors. Physiol. Behav. 2014, 129, 237–254. [Google Scholar] [CrossRef]
- Fragoso, J.; Santos, G.C.J.; da Silva, H.T.; Loizon, E.; Souza, V.d.O.N.; Vidal, H.; Guedes, R.C.A.; Costa-Silva, J.H.; Aragão, R.d.S.; Pirola, L.; et al. Effects of maternal low-protein diet and spontaneous physical activity on the transcription of neurotrophic factors in the placenta and the brains of mothers and offspring rats. J. Dev. Orig. Health Dis. 2020, 12, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jenkins, S.; Mattern, V.; Comuzzie, A.G.; Cox, L.A.; Huber, H.F.; Nathanielsz, P.W. Effect of moderate, 30 percent global maternal nutrient reduction on fetal and postnatal baboon phenotype. J. Med Primatol. 2017, 46, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Rajarethnem, H.T.; Megur Ramakrishna Bhat, K.; Jc, M.; Kumar Gopalkrishnan, S.; Mugundhu Gopalram, R.B.; Rai, K.S. Combined Supplementation of Choline and Docosahexaenoic Acid during Pregnancy Enhances Neurodevelopment of Fetal Hippocampus. Neurol. Res. Int. 2017, 2017, 8748706. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, I.; Hussein, N.; Baumann, M.H.; Di Martino, C.; Salem, N. An n-3 fatty acid deficiency impairs rat spatial learning in the Barnes maze. Behav. Neurosci. 2009, 123, 196–205. [Google Scholar] [CrossRef]
- Liu, D.; Teng, W.; Shan, Z.; Yu, X.; Gao, Y.; Wang, S.; Fan, C.; Wang, H.; Zhang, H. The Effect of Maternal Subclinical Hypothyroidism During Pregnancy on Brain Development in Rat Offspring. Thyroid 2010, 20, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, Y.; Yu, X.; Wang, X.; Bao, S.; Li, J.; Fan, C.; Shan, Z.; Teng, W. Maternal Subclinical Hypothyroidism Impairs Neurodevelopment in Rat Offspring by Inhibiting the CREB Signaling Pathway. Mol. Neurobiol. 2014, 52, 432–441. [Google Scholar] [CrossRef]
- Tran, P.V.; Kennedy, B.C.; Lien, Y.-C.; Simmons, R.A.; Georgieff, M.K. Fetal iron deficiency induces chromatin remodeling at the Bdnf locus in adult rat hippocampus. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R276–R282. [Google Scholar] [CrossRef]
- Lien, Y.-C.; E Condon, D.; Georgieff, M.K.; A Simmons, R.; Tran, P.V. Dysregulation of Neuronal Genes by Fetal-Neonatal Iron Deficiency Anemia Is Associated with Altered DNA Methylation in the Rat Hippocampus. Nutrients 2019, 11, 1191. [Google Scholar] [CrossRef]
- Barks, A.K.; Beeson, M.M.; Hallstrom, T.C.; Georgieff, M.K.; Tran, P.V. Developmental Iron Deficiency Dysregulates TET Activity and DNA Hydroxymethylation in the Rat Hippocampus and Cerebellum. Dev. Neurosci. 2022, 44, 80–90. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, G.; Liu, H.; Chang, H.; Wilson, J.X. Folic acid enhances Notch signaling, hippocampal neurogenesis, and cognitive function in a rat model of cerebral ischemia. Nutr. Neurosci. 2012, 15, 55–61. [Google Scholar] [CrossRef]
- Araki, R.; Nishida, S.; Nakajima, Y.; Iwakumo, A.; Tachioka, H.; Kita, A.; Yabe, T. Low folate induces abnormal neuronal maturation and DNA hypomethylation of neuronal differentiation-related genes in cultured mouse neural stem and progenitor cells. Heliyon 2021, 7, e08071. [Google Scholar] [CrossRef] [PubMed]
- Gawlińska, K.; Gawliński, D.; Kowal-Wiśniewska, E.; Jarmuż-Szymczak, M.; Filip, M. Alteration of the Early Development Environment by Maternal Diet and the Occurrence of Autistic-like Phenotypes in Rat Offspring. Int. J. Mol. Sci. 2021, 22, 9662. [Google Scholar] [CrossRef] [PubMed]
- Gawlińska, K.; Gawliński, D.; Borczyk, M.; Korostyński, M.; Przegaliński, E.; Filip, M. A Maternal High-Fat Diet during Early Development Provokes Molecular Changes Related to Autism Spectrum Disorder in the Rat Offspring Brain. Nutrients 2021, 13, 3212. [Google Scholar] [CrossRef]
- Czarzasta, K.; Makowska-Zubrycka, M.; Kasarello, K.; Skital, V.M.; Tyszkowska, K.; Matusik, K.; Jesion, A.; Wojciechowska, M.; Segiet, A.; Wrzesien, R.; et al. A rat model to study maternal depression during pregnancy and postpartum periods, its comorbidity with cardiovascular diseases and neurodevelopmental impact in the offspring. Physiol. Behav. 2018, 199, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Smith, R.M.; Edwards, K.S.; Givens, B.; Tilley, M.R.; Beversdorf, D.Q. Combined effect of maternal serotonin transporter genotype and prenatal stress in modulating offspring social interaction in mice. Int. J. Dev. Neurosci. 2010, 28, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Matsui, F.; Hecht, P.; Yoshimoto, K.; Watanabe, Y.; Morimoto, M.; Fritsche, K.; Will, M.; Beversdorf, D. DHA Mitigates Autistic Behaviors Accompanied by Dopaminergic Change in a Gene/Prenatal Stress Mouse Model. Neuroscience 2018, 371, 407–419. [Google Scholar] [CrossRef]
- Chan, Y.L.; Saad, S.; Pollock, C.; Oliver, B.; Al-Odat, I.; Zaky, A.A.; Jones, N.; Chen, H. Impact of maternal cigarette smoke exposure on brain inflammation and oxidative stress in male mice offspring. Sci. Rep. 2016, 6, 25881. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-Y.; Kim, J.; Bin Kwon, O.; Jung, J.H.; An, K.; Jeong, A.Y.; Lee, C.J.; Choi, Y.-B.; Bailey, C.H.; Kandel, E.R.; et al. Input-specific synaptic plasticity in the amygdala is regulated by neuroligin-1 via postsynaptic NMDA receptors. Proc. Natl. Acad. Sci. 2010, 107, 4710–4715. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.-F.; Xu, J.-C.; Tereshchenko, Y.; Novak, D.; Schachner, M.; Kleene, R. Neural Cell Adhesion Molecule Modulates Dopaminergic Signaling and Behavior by Regulating Dopamine D2Receptor Internalization. J. Neurosci. 2009, 29, 14752–14763. [Google Scholar] [CrossRef]
- Niedzwiedz-Massey, V.M.; Douglas, J.C.; Rafferty, T.; Wight, P.A.; Kane, C.J.; Drew, P.D. Ethanol modulation of hippocampal neuroinflammation, myelination, and neurodevelopment in a postnatal mouse model of fetal alcohol spectrum disorders. Neurotoxicol. Teratol. 2021, 87, 107015. [Google Scholar] [CrossRef]
- Cantacorps, L.; Alfonso-Loeches, S.; Guerri, C.; Valverde, O. Long-term epigenetic changes in offspring mice exposed to alcohol during gestation and lactation. J. Psychopharmacol. 2019, 33, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; LeBlanc, G.L.; Brengel, E.K.; Hekmatyar, K.S.; Kulkarni, P.; Ferris, C.F.; Klintsova, A.Y. Reduced and delayed myelination and volume of corpus callosum in an animal model of Fetal Alcohol Spectrum Disorders partially benefit from voluntary exercise. Sci. Rep. 2022, 12, 10653. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Fu, Y.; Lu, R.; Bi, Y.; Zhang, L.; Zhang, C.; Aschner, M.; Li, X.; Chen, R. Metabolomics analysis explores the rescue to neurobehavioral disorder induced by maternal PM2.5 exposure in mice. Ecotoxicol. Environ. Saf. 2018, 169, 687–695. [Google Scholar] [CrossRef]
- Ehsanifar, M.; Jafari, A.J.; Nikzad, H.; Zavareh, M.S.; Atlasi, M.A.; Mohammadi, H.; Tameh, A.A. Prenatal exposure to diesel exhaust particles causes anxiety, spatial memory disorders with alters expression of hippocampal pro-inflammatory cytokines and NMDA receptor subunits in adult male mice offspring. Ecotoxicol. Environ. Saf. 2019, 176, 34–41. [Google Scholar] [CrossRef]
- Bolton, J.L.; Marinero, S.; Hassanzadeh, T.; Natesan, D.; Le, D.; Belliveau, C.; Mason, S.N.; Auten, R.L.; Bilbo, S.D. Gestational Exposure to Air Pollution Alters Cortical Volume, Microglial Morphology, and Microglia-Neuron Interactions in a Sex-Specific Manner. Front. Synaptic Neurosci. 2017, 9, 10. [Google Scholar] [CrossRef]
- Pol, H.E.H.; Hoek, H.W.; Susser, E.; Brown, A.S.; Dingemans, A.; Schnack, H.G.; van Haren, N.E.; Ramos, L.M.P.; Wied, C.C.G.-D.; Kahn, R.S. Prenatal Exposure to Famine and Brain Morphology in Schizophrenia. Am. J. Psychiatry 2000, 157, 1170–1172. [Google Scholar] [CrossRef]
- Cortés-Albornoz, M.C.; García-Guáqueta, D.P.; Velez-Van-Meerbeke, A.; Talero-Gutiérrez, C. Maternal Nutrition and Neurodevelopment: A Scoping Review. Nutrients 2021, 13, 3530. [Google Scholar] [CrossRef]
- Peter, C.J.; Fischer, L.K.; Kundakovic, M.; Garg, P.; Jakovcevski, M.; Dincer, A.; Amaral, A.C.; Ginns, E.I.; Galdzicka, M.; Bryce, C.P.; et al. DNA Methylation Signatures of Early Childhood Malnutrition Associated with Impairments in Attention and Cognition. Biol. Psychiatry 2016, 80, 765–774. [Google Scholar] [CrossRef]
- Prado, E.L.; Dewey, K.G. Nutrition and brain development in early life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef]
- Staubo, S.C.; Aakre, J.A.; Vemuri, P.; Syrjanen, J.A.; Mielke, M.M.; Geda, Y.E.; Kremers, W.K.; Machulda, M.M.; Knopman, D.S.; Petersen, R.C.; et al. Mediterranean diet, micronutrients and macronutrients, and MRI measures of cortical thickness. Alzheimer’s Dement. 2016, 13, 168–177. [Google Scholar] [CrossRef]
- Gu, Y.; Brickman, A.M.; Stern, Y.; Habeck, C.G.; Razlighi, Q.R.; Luchsinger, J.A.; Manly, J.J.; Schupf, N.; Mayeux, R.; Scarmeas, N. Mediterranean diet and brain structure in a multiethnic elderly cohort. Neurology 2015, 85, 1744–1751. [Google Scholar] [CrossRef]
- Rodrigues, B.; Coelho, A.; Portugal-Nunes, C.; Magalhães, R.; Moreira, P.S.; Castanho, T.C.; Amorim, L.; Marques, P.; Soares, J.M.; Sousa, N.; et al. Higher Adherence to the Mediterranean Diet Is Associated with Preserved White Matter Integrity and Altered Structural Connectivity. Front. Neurosci. 2020, 14, 786. [Google Scholar] [CrossRef]
- House, J.S.; Mendez, M.; Maguire, R.L.; Gonzalez-Nahm, S.; Huang, Z.; Daniels, J.; Murphy, S.K.; Fuemmeler, B.F.; Wright, F.A.; Hoyo, C. Periconceptional Maternal Mediterranean Diet Is Associated with Favorable Offspring Behaviors and Altered CpG Methylation of Imprinted Genes. Front. Cell Dev. Biol. 2018, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Barry, D.; Ellul, S.; Watters, L.; Lee, D.; Haluska, R.; White, R. The ketogenic diet in disease and development. Int. J. Dev. Neurosci. 2018, 68, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Sussman, D.; Ellegood, J.; Henkelman, M. A gestational ketogenic diet alters maternal metabolic status as well as offspring physiological growth and brain structure in the neonatal mouse. BMC Pregnancy Childbirth 2013, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- Sussman, D.; Germann, J.; Henkelman, M. Gestational ketogenic diet programs brain structure and susceptibility to depression & anxiety in the adult mouse offspring. Brain Behav. 2014, 5, e00300. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, T.; Metzger, B.E.; Burns, W.J.; Burns, K. Correlations between Antepartum Maternal Metabolism and Intelligence of Offspring. New Engl. J. Med. 1991, 325, 911–916. [Google Scholar] [CrossRef]
- Bronisz, A.; Ozorowski, M.; Hagner-Derengowska, M. Pregnancy Ketonemia and Development of the Fetal Central Nervous System. Int. J. Endocrinol. 2018, 2018, 1242901. [Google Scholar] [CrossRef]
- Hanson, M.A.; Bardsley, A.; De-Regil, L.M.; Moore, S.E.; Oken, E.; Poston, L.; Ma, R.C.; McAuliffe, F.M.; Maleta, K.; Purandare, C.N.; et al. The International Federation of Gynecology and Obstetrics (FIGO) recommendations on adolescent, preconception, and maternal nutrition: “Think Nutrition First”. Int. J. Gynecol. Obstet. 2015, 131, S213–S253. [Google Scholar] [CrossRef]
- Cetin, I.; Koletzko, B. Long-chain ω-3 fatty acid supply in pregnancy and lactation. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 297–302. [Google Scholar] [CrossRef]
- Makrides, M.; Neumann, M.; Byard, R.; Simmer, K.; Gibson, R. Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am. J. Clin. Nutr. 1994, 60, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Dietary omega 3 fatty acids and the developing brain. Brain Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef]
- Simon, P.; Dupuis, R.; Costentin, J. Thigmotaxis as an index of anxiety in mice. Influence of dopaminergic transmissions. Behav. Brain Res. 1994, 61, 59–64. [Google Scholar] [CrossRef]
- Martinat, M.; Rossitto, M.; Di Miceli, M.; Layé, S. Perinatal Dietary Polyunsaturated Fatty Acids in Brain Development, Role in Neurodevelopmental Disorders. Nutrients 2021, 13, 1185. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, K.; Matsumura, K.; Tsuchida, A.; Kasamatsu, H.; Tanaka, T.; Ito, M.; Inadera, H. Maternal dietary intake of fish and PUFAs and child neurodevelopment at 6 months and 1 year of age: A nationwide birth cohort—The Japan Environment and Children’s Study (JECS). Am. J. Clin. Nutr. 2020, 112, 1295–1303. [Google Scholar] [CrossRef]
- Abel, M.H.; Caspersen, I.H.; Meltzer, H.M.; Haugen, M.; E Brandlistuen, R.; Aase, H.; Alexander, J.; E Torheim, L.; Brantsæter, A.-L. Suboptimal Maternal Iodine Intake Is Associated with Impaired Child Neurodevelopment at 3 Years of Age in the Norwegian Mother and Child Cohort Study. J. Nutr. 2017, 147, 1314–1324. [Google Scholar] [CrossRef]
- Abel, M.H.; Ystrom, E.; Caspersen, I.H.; Meltzer, H.M.; Aase, H.; Torheim, L.E.; Askeland, R.B.; Reichborn-Kjennerud, T.; Brantsæter, A.L. Maternal Iodine Intake and Offspring Attention-Deficit/Hyperactivity Disorder: Results from a Large Prospective Cohort Study. Nutrients 2017, 9, 1239. [Google Scholar] [CrossRef] [PubMed]
- Dineva, M.; Fishpool, H.; Rayman, M.P.; Mendis, J.; Bath, S.C. Systematic review and meta-analysis of the effects of iodine supplementation on thyroid function and child neurodevelopment in mildly-to-moderately iodine-deficient pregnant women. Am. J. Clin. Nutr. 2020, 112, 389–412. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.N.; Lazarus, J.H.; Moreno-Reyes, R.; Zimmermann, M.B. Consequences of iodine deficiency and excess in pregnant women: An overview of current knowns and unknowns. Am. J. Clin. Nutr. 2016, 104, 918S–923S. [Google Scholar] [CrossRef] [PubMed]
- Ballestín, S.S.; Campos, M.I.G.; Ballestín, J.B.; Bartolomé, M.J.L. Is Supplementation with Micronutrients Still Necessary during Pregnancy? A Review. Nutrients 2021, 13, 3134. [Google Scholar] [CrossRef]
- Georgieff, M.K. The importance of iron deficiency in pregnancy on fetal, neonatal, and infant neurodevelopmental outcomes. Int. J. Gynecol. Obstet. 2023, 162, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Maternal gestational iron status and infant haematological and neurodevelopmental outcomes. BJOG: Int. J. Obstet. Gynaecol. 2023, 130, 92–98. [Google Scholar] [CrossRef]
- Carlson, E.S.; Tkac, I.; Magid, R.; O’connor, M.B.; Andrews, N.C.; Schallert, T.; Gunshin, H.; Georgieff, M.K.; Petryk, A. Iron Is Essential for Neuron Development and Memory Function in Mouse Hippocampus. J. Nutr. 2009, 139, 672–679. [Google Scholar] [CrossRef]
- Tran, P.V.; Kennedy, B.C.; Pisansky, M.T.; Won, K.-J.; Gewirtz, J.C.; A Simmons, R.; Georgieff, M.K. Prenatal Choline Supplementation Diminishes Early-Life Iron Deficiency–Induced Reprogramming of Molecular Networks Associated with Behavioral Abnormalities in the Adult Rat Hippocampus. J. Nutr. 2016, 146, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Kubant, R.; Cho, C.E.; Pannia, E.; Hammoud, R.; Yang, N.V.; Simonian, R.; Anderson, G.H. Methyl donor micronutrients, hypothalamic development and programming for metabolic disease. Neurosci. Biobehav. Rev. 2024, 157, 105512. [Google Scholar] [CrossRef] [PubMed]
- Berti, C.; Fekete, K.; Dullemeijer, C.; Trovato, M.; Souverein, O.; Cavelaars, A.A.; Dhonukshe-Rutten, R.; Massari, M.; Decsi, T.; Veer, P.V.; et al. Folate Intake and Markers of Folate Status in Women of Reproductive Age, Pregnant and Lactating Women: A Meta-Analysis. J. Nutr. Metab. 2012, 2012, 470656. [Google Scholar] [CrossRef]
- Parisi, F.; di Bartolo, I.; Savasi, V.; Cetin, I. Micronutrient supplementation in pregnancy: Who, what and how much? Obstet. Med. 2018, 12, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, K.; Zhao, D.; Li, L. The association between maternal use of folic acid supplements during pregnancy and risk of autism spectrum disorders in children: A meta-analysis. Mol. Autism 2017, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Iosif, A.-M.; Angel, E.G.; Ozonoff, S. Association of Maternal Prenatal Vitamin Use With Risk for Autism Spectrum Disorder Recurrence in Young Siblings. JAMA Psychiatry 2019, 76, 391–398. [Google Scholar] [CrossRef]
- Levine, S.Z.; Kodesh, A.; Viktorin, A.; Smith, L.; Uher, R.; Reichenberg, A.; Sandin, S. Association of Maternal Use of Folic Acid and Multivitamin Supplements in the Periods Before and During Pregnancy With the Risk of Autism Spectrum Disorder in Offspring. JAMA Psychiatry 2018, 75, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.; Riley, A.W.; Volk, H.; Caruso, D.; Hironaka, L.; Sices, L.; Hong, X.; Wang, G.; Ji, Y.; Brucato, M.; et al. Maternal Multivitamin Intake, Plasma Folate and Vitamin B12 Levels and Autism Spectrum Disorder Risk in Offspring. Paediatr. Périnat. Epidemiol. 2017, 32, 100–111. [Google Scholar] [CrossRef]
- Valera-Gran, D.; Navarrete-Muñoz, E.M.; de la Hera, M.G.; Fernández-Somoano, A.; Tardón, A.; Ibarluzea, J.; Balluerka, N.; Murcia, M.; González-Safont, L.; Romaguera, D.; et al. Effect of maternal high dosages of folic acid supplements on neurocognitive development in children at 4–5 y of age: The prospective birth cohort Infancia y Medio Ambiente (INMA) study. Am. J. Clin. Nutr. 2017, 106, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhai, Z.; Wang, S.; Song, X.; Qiu, T.; Yu, L.; Zhai, Q.; Zhang, H. The role and impact of abnormal vitamin levels in autism spectrum disorders. Food Funct. 2024, 15, 1099–1115. [Google Scholar] [CrossRef] [PubMed]
- Larqué, E.; Morales, E.; Leis, R.; Blanco-Carnero, J.E. Maternal and Foetal Health Implications of Vitamin D Status during Pregnancy. Ann. Nutr. Metab. 2018, 72, 179–192. [Google Scholar] [CrossRef]
- Eyles, D.W.; Smith, S.; Kinobe, R.; Hewison, M.; McGrath, J.J. Distribution of the Vitamin D receptor and 1α-hydroxylase in human brain. J. Chem. Neuroanat. 2005, 29, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Vinkhuyzen, A.A.E.; Eyles, D.W.; Burne, T.H.J.; Blanken, L.M.E.; Kruithof, C.J.; Verhulst, F.; White, T.; Jaddoe, V.W.; Tiemeier, H.; McGrath, J.J. Gestational vitamin D deficiency and autism spectrum disorder. BJPsych Open 2017, 3, 85–90. [Google Scholar] [CrossRef]
- A E Vinkhuyzen, A.; Eyles, D.W.; Burne, T.H.J.; E Blanken, L.M.; Kruithof, C.J.; Verhulst, F.; Jaddoe, V.W.; Tiemeier, H.; McGrath, J.J. Gestational vitamin D deficiency and autism-related traits: The Generation R Study. Mol. Psychiatry 2016, 23, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Cui, X.; Eyles, D. Developmental vitamin D deficiency and autism: Putative pathogenic mechanisms. J. Steroid Biochem. Mol. Biol. 2018, 175, 108–118. [Google Scholar] [CrossRef]
- Edlow, A.G. Maternal obesity and neurodevelopmental and psychiatric disorders in offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef]
- van der Burg, J.W.; Sen, S.; Chomitz, V.R.; Seidell, J.C.; Leviton, A.; Dammann, O. The role of systemic inflammation linking maternal BMI to neurodevelopment in children. Pediatr. Res. 2015, 79, 3–12. [Google Scholar] [CrossRef]
- Brion, M.-J.; Zeegers, M.; Jaddoe, V.; Verhulst, F.; Tiemeier, H.; Lawlor, D.A.; Smith, G.D. Intrauterine Effects of Maternal Prepregnancy Overweight on Child Cognition and Behavior in 2 Cohorts. Pediatrics 2011, 127, e202–e211. [Google Scholar] [CrossRef] [PubMed]
- Van De Maele, K.; Bogaerts, A.; De Schepper, J.; Provyn, S.; Ceulemans, D.; Guelinckx, I.; Gies, I.; Devlieger, R. Adiposity, psychomotor and behaviour outcomes of children born after maternal bariatric surgery. Pediatr. Obes. 2020, 16, e12749. [Google Scholar] [CrossRef]
- Dell’agnolo, C.M.; Cyr, C.; De Montigny, F.; Carvalho, M.D.D.B.; Pelloso, S.M. Pregnancy after Bariatric Surgery: Obstetric and Perinatal Outcomes and the Growth and Development of Children. Obes. Surg. 2015, 25, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.A.; Machado, B.M.; da Rosa, R.R.; Rigoni, M.d.S.; Casagrande, D.S.; Mottin, C.C.; Schaan, B.D. Association of Maternal Roux-en-Y Gastric Bypass with Obstetric Outcomes and Fluid Intelligence in Offspring. Obes. Surg. 2018, 28, 3611–3620. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lyo, V.; Groth, S.W. The impact of maternal bariatric surgery on long-term health of offspring: A scoping review. Pediatr. Res. 2023, 94, 1619–1630. [Google Scholar] [CrossRef]
- Lebel, C.; MacKinnon, A.; Bagshawe, M.; Tomfohr-Madsen, L.; Giesbrecht, G. Elevated depression and anxiety symptoms among pregnant individuals during the COVID-19 pandemic. J. Affect. Disord. 2020, 277, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Marcus, S.M.; Heringhausen, J.E. Depression in Childbearing Women: When Depression Complicates Pregnancy. Prim. Care Clin. Off. Pract. 2009, 36, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Heuvel, M.v.D.; Johannes, M.; Henrichs, J.; Bergh, B.V.D. Maternal mindfulness during pregnancy and infant socio-emotional development and temperament: The mediating role of maternal anxiety. Early Hum. Dev. 2015, 91, 103–108. [Google Scholar] [CrossRef]
- Field, T.; Diego, M.; Hernandez-Reif, M.; Figueiredo, B.; Schanberg, S.; Kuhn, C. Sleep disturbances in depressed pregnant women and their newborns. Infant Behav. Dev. 2007, 30, 127–133. [Google Scholar] [CrossRef]
- Deave, T.; Heron, J.; Evans, J.; Emond, A. The impact of maternal depression in pregnancy on early child development. BJOG Int. J. Obstet. Gynaecol. 2008, 115, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Weikum, W.M.; Oberlander, T.F.; Hensch, T.K.; Werker, J.F. Prenatal exposure to antidepressants and depressed maternal mood alter trajectory of infant speech perception. Proc. Natl. Acad. Sci. USA 2012, 109, 17221–17227. [Google Scholar] [CrossRef] [PubMed]
- Wolford, E.; Lahti, M.; Tuovinen, S.; Lahti, J.; Lipsanen, J.; Savolainen, K.; Heinonen, K.; Hämäläinen, E.; Kajantie, E.; Pesonen, A.-K.; et al. Maternal depressive symptoms during and after pregnancy are associated with attention-deficit/hyperactivity disorder symptoms in their 3- to 6-year-old children. PLoS ONE 2017, 12, e0190248. [Google Scholar] [CrossRef]
- Croen, L.A.; Grether, J.K.; Yoshida, C.K.; Odouli, R.; Hendrick, V. Antidepressant Use During Pregnancy and Childhood Autism Spectrum Disorders. Arch. Gen. Psychiatry 2011, 68, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, M.; Guillaume, B.; Chong, Y.-S.; Chen, H.; Fortier, M.V.; Meaney, M.J.; Qiu, A. FKBP5 Moderates the Association between Antenatal Maternal Depressive Symptoms and Neonatal Brain Morphology. Neuropsychopharmacology 2017, 43, 564–570. [Google Scholar] [CrossRef]
- Antonelli, M.C.; Pallarés, M.E.; Ceccatelli, S.; Spulber, S. Long-term consequences of prenatal stress and neurotoxicants exposure on neurodevelopment. Prog. Neurobiol. 2017, 155, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Class, Q.A.; Abel, K.M.; Khashan, A.S.; Rickert, M.E.; Dalman, C.; Larsson, H.; Hultman, C.M.; Långström, N.; Lichtenstein, P.; D‘onofrio, B.M. Offspring psychopathology following preconception, prenatal and postnatal maternal bereavement stress. Psychol. Med. 2013, 44, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.L.; Lyall, K.; Rich-Edwards, J.W.; Ascherio, A.; Weisskopf, M.G. Maternal exposure to intimate partner abuse before birth is associated with autism spectrum disorder in offspring. Autism 2015, 20, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Manzari, N.; Matvienko-Sikar, K.; Baldoni, F.; O’Keeffe, G.W.; Khashan, A.S. Prenatal Maternal Stress and Risk of Neurode-velopmental Disorders in the Offspring: A Systematic Review and Meta-Analysis. Soc. Psychiatry Psychiatr. Epidemiol. 2019, 54, 1299–1309. [Google Scholar] [CrossRef]
- Shamsuddin, K.; Fadzil, F.; Ismail, W.S.W.; Shah, S.A.; Omar, K.; Muhammad, N.A.; Jaffar, A.; Ismail, A.; Mahadevan, R. Correlates of depression, anxiety and stress among Malaysian university students. Asian J. Psychiatry 2013, 6, 318–323. [Google Scholar] [CrossRef]
- Suter, M.A.; Aagaard, K.M. The impact of tobacco chemicals and nicotine on placental development. Prenat. Diagn. 2020, 40, 1193–1200. [Google Scholar] [CrossRef]
- Archie, S.R.; Sifat, A.E.; Mara, D.; Ahn, Y.; Akter, K.A.; Zhang, Y.; Cucullo, L.; Abbruscato, T.J. Impact of in-utero electronic cigarette exposure on neonatal neuroinflammation, oxidative stress and mitochondrial function. Front. Pharmacol. 2023, 14, 1227145. [Google Scholar] [CrossRef]
- Buck, J.M.; Yu, L.; Knopik, V.S.; A Stitzel, J. DNA methylome perturbations: An epigenetic basis for the emergingly heritable neurodevelopmental abnormalities associated with maternal smoking and maternal nicotine exposure. Biol. Reprod. 2021, 105, 644–666. [Google Scholar] [CrossRef]
- Kozieł, S.; Żądzińska, E.; Gomula, A. Parental smoking during pregnancy and head shape and size in school children. Ann. Hum. Biol. 2018, 45, 401–405. [Google Scholar] [CrossRef]
- Rose-Jacobs, R.; Richardson, M.A.; Buchanan-Howland, K.; Chen, C.A.; Cabral, H.; Heeren, T.C.; Liebschutz, J.; Forman, L.; Frank, D.A. Intrauterine exposure to tobacco and executive functioning in high school. Drug Alcohol Depend. 2017, 176, 169–175. [Google Scholar] [CrossRef]
- Thapar, A.; Fowler, T.; Rice, F.; Scourfield, J.; Bree, M.v.D.; Thomas, H.; Harold, G.; Hay, D. Maternal Smoking During Pregnancy and Attention Deficit Hyperactivity Disorder Symptoms in Offspring. Am. J. Psychiatry 2003, 160, 1985–1989. [Google Scholar] [CrossRef] [PubMed]
- Linnet, K.M.; Wisborg, K.; Obel, C.; Secher, N.J.; Thomsen, P.H.; Agerbo, E.; Henriksen, T.B. Smoking During Pregnancy and the Risk for Hyperkinetic Disorder in Offspring. Pediatrics 2005, 116, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Froggatt, S.; Reissland, N.; Covey, J. The effects of prenatal cigarette and e-cigarette exposure on infant neurobehaviour: A comparison to a control group. E Clin. Med. 2020, 28, 100602. [Google Scholar] [CrossRef]
- World Health Organisation. Global Status Report on Alcohol and Health 2018; World Health Organisation: Geneva, Switzerland, 2018; Available online: https://www.who.int/publications/i/item/9789241565639 (accessed on 22 January 2024).
- Sharif, S.; Lakshmanan, N.; Sharif, F.; Ryan, S. Spectrum of MRI findings of foetal alcohol syndrome disorders—What we know and what we need to know! BJR Open 2023, 5, 20210063. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Blankenship, J.; Marais, A.-S.; Gossage, J.P.; Kalberg, W.O.; Joubert, B.; Cloete, M.; Barnard, R.; De Vries, M.; Hasken, J.; et al. Maternal alcohol consumption producing fetal alcohol spectrum disorders (FASD): Quantity, frequency, and timing of drinking. Drug Alcohol Depend. 2013, 133, 502–512. [Google Scholar] [CrossRef]
- Subramoney, S.; Eastman, E.; Adnams, C.; Stein, D.J.; Donald, K.A. The Early Developmental Outcomes of Prenatal Alcohol Exposure: A Review. Front. Neurol. 2018, 9, 1108. [Google Scholar] [CrossRef] [PubMed]
- Mews, P.; Egervari, G.; Nativio, R.; Sidoli, S.; Donahue, G.; Lombroso, S.I.; Alexander, D.C.; Riesche, S.L.; Heller, E.A.; Nestler, E.J.; et al. Alcohol metabolism contributes to brain histone acetylation. Nature 2019, 574, 717–721. [Google Scholar] [CrossRef]
- Popova, S.; Charness, M.E.; Burd, L.; Crawford, A.; Hoyme, H.E.; Mukherjee, R.A.S.; Riley, E.P.; Elliott, E.J. Fetal alcohol spectrum disorders. Nat. Rev. Dis. Prim. 2023, 9, 11. [Google Scholar] [CrossRef]
- Wang, X.; Carlson, V.C.C.; Studholme, C.; Newman, N.; Ford, M.M.; Grant, K.A.; Kroenke, C.D. In utero MRI identifies consequences of early-gestation alcohol drinking on fetal brain development in rhesus macaques. Proc. Natl. Acad. Sci. USA 2020, 117, 10035–10044. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.J.; Richards, T.; Aylward, E.H.; Olson, H.C.; Kerns, K.; Brooks, A.; Coggins, T.E.; Davies, J.; Dorn, S.; Gendler, B.; et al. Magnetic resonance spectroscopy outcomes from a comprehensive magnetic resonance study of children with fetal alcohol spectrum disorders. Magn. Reson. Imaging 2009, 27, 760–778. [Google Scholar] [CrossRef]
- Seagull, F.N.; Mowery, J.L.; Simpson, P.M.; Robinson, T.R.; Martier, S.S.; Sokol, R.J.; McCarver-May, D.G. Maternal Assessment of Infant Development: Associations with Alcohol and Drug Use in Pregnancy. Clin. Pediatr. 1996, 35, 621–628. [Google Scholar] [CrossRef]
- Yi, C.; Wang, Q.; Qu, Y.; Niu, J.; Oliver, B.G.; Chen, H. In-utero exposure to air pollution and early-life neural development and cognition. Ecotoxicol. Environ. Saf. 2022, 238, 113589. [Google Scholar] [CrossRef]
- Zhou, G.; He, T.; Huang, H.; Feng, F.; Liu, X.; Li, Z.; Zhang, Y.; Ba, Y. Prenatal ambient air pollution exposure and SOD2 promoter methylation in maternal and cord blood. Ecotoxicol. Environ. Saf. 2019, 181, 428–434. [Google Scholar] [CrossRef]
- Grevendonk, L.; Janssen, B.G.; Vanpoucke, C.; Lefebvre, W.; Hoxha, M.; Bollati, V.; Nawrot, T.S. Mitochondrial oxidative DNA damage and exposure to particulate air pollution in mother-newborn pairs. Environ. Health 2016, 15, 10. [Google Scholar] [CrossRef]
- Lee, A.; Hsu, H.-H.L.; Chiu, Y.-H.M.; Bose, S.; Rosa, M.J.; Kloog, I.; Wilson, A.; Schwartz, J.; Cohen, S.; Coull, B.A.; et al. Prenatal fine particulate exposure and early childhood asthma: Effect of maternal stress and fetal sex. J. Allergy Clin. Immunol. 2017, 141, 1880–1886. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Xu, X.; Wang, Q.; Zhang, Z.; Meng, P.; Huo, X. Maternal exposure to atmospheric PM2.5 and fetal brain development: Associations with BAI1 methylation and thyroid hormones. Environ. Pollut. 2022, 308, 119665. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Li, J.; Cheng, L.; Deng, Y.; Li, Y.; Yan, Z.; Duan, L.; Niu, Q.; Tang, D. Prenatal polycyclic aromatic hydrocarbons metabolites, cord blood telomere length, and neonatal neurobehavioral development. Environ. Res. 2019, 174, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Perera, F.P.; Wheelock, K.; Wang, Y.; Tang, D.; Margolis, A.E.; Badia, G.; Cowell, W.; Miller, R.L.; Rauh, V.; Wang, S.; et al. Combined effects of prenatal exposure to polycyclic aromatic hydrocarbons and material hardship on child ADHD behavior problems. Environ. Res. 2017, 160, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Shu, Y.-H.; Chow, T.; Lurmann, F.W.; Yu, X.; Martinez, M.P.; Carter, S.A.; Eckel, S.P.; Chen, J.-C.; Chen, Z.; et al. Prenatal Exposure to Air Pollution and Autism Spectrum Disorder: Sensitive Windows of Exposure and Sex Differences. Environ. Health Perspect. 2022, 130, 17008. [Google Scholar] [CrossRef]
- Yu, X.; Rahman, M.; Carter, S.A.; Lin, J.C.; Zhuang, Z.; Chow, T.; Lurmann, F.W.; Kleeman, M.J.; Martinez, M.P.; van Donkelaar, A.; et al. Prenatal air pollution, maternal immune activation, and autism spectrum disorder. Environ. Int. 2023, 179, 108148. [Google Scholar] [CrossRef] [PubMed]
- Raz, R.; Roberts, A.L.; Lyall, K.; Hart, J.E.; Just, A.C.; Laden, F.; Weisskopf, M.G. Autism Spectrum Disorder and Particulate Matter Air Pollution before, during, and after Pregnancy: A Nested Case–Control Analysis within the Nurses’ Health Study II Cohort. Environ. Health Perspect. 2015, 123, 264–270. [Google Scholar] [CrossRef]
- Anekwe, C.V.; Jarrell, A.R.; Townsend, M.J.; Gaudier, G.I.; Hiserodt, J.M.; Stanford, F.C. Socioeconomics of Obesity. Curr. Obes. Rep. 2020, 9, 272–279. [Google Scholar] [CrossRef]
- Kim, M.K.; Lee, S.M.; Bae, S.-H.; Kim, H.J.; Lim, N.G.; Yoon, S.-J.; Lee, J.Y.; Jo, M.-W. Socioeconomic status can affect pregnancy outcomes and complications, even with a universal healthcare system. Int. J. Equity Health 2018, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Sow, M.; Raynault, M.-F.; De Spiegelaere, M. Associations between socioeconomic status and pregnancy outcomes: A greater magnitude of inequalities in perinatal health in Montreal than in Brussels. BMC Public Health 2022, 22, 829. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.; Moffat, M.; Arisa, O.; Jesurasa, A.; Richmond, C.; Odeniyi, A.; Bambra, C.; Rankin, J.; Brown, H.; Bishop, J.; et al. Socioeconomic inequalities and adverse pregnancy outcomes in the UK and Republic of Ireland: A systematic review and meta-analysis. BMJ Open 2021, 11, e042753. [Google Scholar] [CrossRef] [PubMed]
- Knickmeyer, R.C.; Xia, K.; Lu, Z.; Ahn, M.; Jha, S.C.; Zou, F.; Zhu, H.; Styner, M.; Gilmore, J.H. Impact of Demographic and Obstetric Factors on Infant Brain Volumes: A Population Neuroscience Study. Cereb. Cortex 2016, 27, 5616–5625. [Google Scholar] [CrossRef]
- Smith, T.A.; Kievit, R.A.; Astle, D.E. Maternal mental health mediates links between socioeconomic status and child development. Curr. Psychol. 2022, 42, 21967–21978. [Google Scholar] [CrossRef] [PubMed]
- Miko, E.; Csaszar, A.; Bodis, J.; Kovacs, K. The Maternal–Fetal Gut Microbiota Axis: Physiological Changes, Dietary Influence, and Modulation Possibilities. Life 2022, 12, 424. [Google Scholar] [CrossRef]
- Sajdel-Sulkowska, E.M. The Impact of Maternal Gut Microbiota during Pregnancy on Fetal Gut–Brain Axis Development and Life-Long Health Outcomes. Microorganisms 2023, 11, 2199. [Google Scholar] [CrossRef]
- Zavatta, A.; Parisi, F.; Mandò, C.; Scaccabarozzi, C.; Savasi, V.M.; Cetin, I. Role of Inflammaging on the Reproductive Function and Pregnancy. Clin. Rev. Allergy Immunol. 2022, 64, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Dawson, S.L.; O’Hely, M.; Jacka, F.N.; Ponsonby, A.-L.; Symeonides, C.; Loughman, A.; Collier, F.; Moreno-Betancur, M.; Sly, P.; Burgner, D.; et al. Maternal prenatal gut microbiota composition predicts child behaviour. EBioMedicine 2021, 68, 103400. [Google Scholar] [CrossRef] [PubMed]
- Cetin, I.; Alvino, G. Intrauterine Growth Restriction: Implications for Placental Metabolism and Transport. A Review. Placenta 2009, 30, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lubrano, C.; Taricco, E.; Coco, C.; Di Domenico, F.; Mandò, C.; Cetin, I. Perinatal and Neonatal Outcomes in Fetal Growth Restriction and Small for Gestational Age. J. Clin. Med. 2022, 11, 2729. [Google Scholar] [CrossRef] [PubMed]
- Figueras, F.; Cruz-Martinez, R.; Sanz-Cortes, M.; Arranz, A.; Illa, M.; Botet, F.; Costas-Moragas, C.; Gratacos, E. Neurobehavioral outcomes in preterm, growth-restricted infants with and without prenatal advanced signs of brain-sparing. Ultrasound Obstet. Gynecol. 2011, 38, 288–294. [Google Scholar] [CrossRef]
- Wan, L.; Luo, K.; Chen, P. Mechanisms Underlying Neurologic Injury in Intrauterine Growth Restriction. J. Child Neurol. 2021, 36, 776–784. [Google Scholar] [CrossRef]
- Brembilla, G.; Righini, A.; Scelsa, B.; Lista, G.; Balestriero, M.; Cesari, E.; Castoldi, F.M.; Di Stasi, M.; Ciardi, C.; Ligato, E.; et al. Neuroimaging and neurodevelopmental outcome after early fetal growth restriction: NEUROPROJECT—FGR. Pediatr. Res. 2021, 90, 869–875. [Google Scholar] [CrossRef]
- Welling, M.S.; Husen, S.C.; Go, A.T.; Groenenberg, I.A.; Willemsen, S.P.; Bijma, H.H.; Steegers-Theunissen, R.P. Growth trajectories of the human fetal brain in healthy and complicated pregnancies and associations with neurodevelopmental outcome in the early life course. Early Hum. Dev. 2020, 151, 105224. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J.D.; Peterson, L.S.; Nesamoney, S.N.; Chang, A.L.; Pasca, A.M.; Marić, I.; Shaw, G.M.; Gaudilliere, B.; Wong, R.J.; Sylvester, K.G.; et al. Perinatal infection, inflammation, preterm birth, and brain injury: A review with proposals for future investigations. Exp. Neurol. 2022, 351, 113988. [Google Scholar] [CrossRef] [PubMed]
- Chawanpaiboon, S.; Vogel, J.P.; Moller, A.-B.; Lumbiganon, P.; Petzold, M.; Hogan, D.; Landoulsi, S.; Jampathong, N.; Kongwattanakul, K.; Laopaiboon, M.; et al. Global, regional, and national estimates of levels of preterm birth in 2014: A systematic review and modelling analysis. Lancet Glob. Health 2018, 7, e37–e46. [Google Scholar] [CrossRef] [PubMed]
- Ortinau, C.; Neil, J. The neuroanatomy of prematurity: Normal brain development and the impact of preterm birth. Clin. Anat. 2014, 28, 168–183. [Google Scholar] [CrossRef]
- Deger, J.; Goethe, E.A.; LoPresti, M.A.; Lam, S. Intraventricular Hemorrhage in Premature Infants: A Historical Review. World Neurosurg. 2021, 153, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Bokobza, C.; Van Steenwinckel, J.; Mani, S.; Mezger, V.; Fleiss, B.; Gressens, P. Neuroinflammation in preterm babies and autism spectrum disorders. Pediatr. Res. 2018, 85, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Rocha-Ferreira, E.; Fleiss, B.; Nijboer, C.H.; Gressens, P.; Mallard, C.; Hagberg, H. Neuroprotection offered by mesenchymal stem cells in perinatal brain injury: Role of mitochondria, inflammation, and reactive oxygen species. J. Neurochem. 2020, 158, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Cetin, I.; Giovannini, N.; Alvino, G.; Agostoni, C.; Riva, E.; Giovannini, M.; Pardi, G. Intrauterine Growth Restriction Is Associated with Changes in Polyunsaturated Fatty Acid Fetal-Maternal Relationships. Pediatr. Res. 2002, 52, 750–755. [Google Scholar] [CrossRef]
- Ramenghi, L.A.; Martinelli, A.; De Carli, A.; Brusati, V.; Mandia, L.; Fumagalli, M.; Triulzi, F.; Mosca, F.; Cetin, I. Cerebral Maturation in IUGR and Appropriate for Gestational Age Preterm Babies. Reprod. Sci. 2011, 18, 469–475. [Google Scholar] [CrossRef]
- Shah, T.J.; Leik, C.E.; Walsh, S.W. Neutrophil Infiltration and Systemic Vascular Inflammation in Obese Women. Reprod. Sci. 2010, 17, 116–124. [Google Scholar] [CrossRef]
- Mandò, C.; Calabrese, S.; Mazzocco, M.I.; Novielli, C.; Anelli, G.M.; Antonazzo, P.; Cetin, I. Sex specific adaptations in placental biometry of overweight and obese women. Placenta 2016, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Lahti-Pulkkinen, M.; Girchenko, P.; Robinson, R.; Lehto, S.M.; Toffol, E.; Heinonen, K.; Reynolds, R.M.; Kajantie, E.; Laivuori, H.; Villa, P.M.; et al. Maternal depression and inflammation during pregnancy. Psychol. Med. 2019, 50, 1839–1851. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, H.; Wang, C.; Broekman, B.F.P.; Chong, Y.-S.; Shek, L.P.; Gluckman, P.D.; Meaney, M.J.; Fortier, M.V.; Qiu, A. Inflammatory modulation of the associations between prenatal maternal depression and neonatal brain. Neuropsychopharmacology 2020, 46, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Zundel, C.G.; Ryan, P.; Brokamp, C.; Heeter, A.; Huang, Y.; Strawn, J.R.; Marusak, H.A. Air pollution, depressive and anxiety disorders, and brain effects: A systematic review. NeuroToxicology 2022, 93, 272–300. [Google Scholar] [CrossRef]
- Zawadzka, A.; Cieślik, M.; Adamczyk, A. The Role of Maternal Immune Activation in the Pathogenesis of Autism: A Review of the Evidence, Proposed Mechanisms and Implications for Treatment. Int. J. Mol. Sci. 2021, 22, 11516. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.S. Interventions to prevent periodontal disease in tobacco-, alcohol-, and drug-dependent individuals. Periodontol. 2000 2020, 84, 84–101. [Google Scholar] [CrossRef]
- Zambon, M.; Mandò, C.; Lissoni, A.; Anelli, G.M.; Novielli, C.; Cardellicchio, M.; Leone, R.; Monari, M.N.; Massari, M.; Cetin, I.; et al. Inflammatory and Oxidative Responses in Pregnancies with Obesity and Periodontal Disease. Reprod. Sci. 2018, 25, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.; Amaral, D.; Van de Water, J. Maternal and fetal antibrain antibodies in development and disease. Dev. Neurobiol. 2012, 72, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Bock, J.; Wainstock, T.; Braun, K.; Segal, M. Stress in Utero: Prenatal Programming of Brain Plasticity and Cognition. Biol. Psychiatry 2015, 78, 315–326. [Google Scholar] [CrossRef]


{kind=link}
| Author, Year | Type of Study | Investigated Variable | Population | Main Findings |
|---|---|---|---|---|
| A. Nakaki [8], 2023 | Randomized clinical trial | Mediterranean Diet | 90 fetal MRI | TBV is positively associated with walnut intake and biomarkers of olive oil consumption. |
| Stress | 90 fetal MRI | The stress reduction group has larger volume of left anterior cingulate lobe. | ||
| E. Ogundipe [9], 2018 | Double-blind randomized placebo-controlled study | DHA | 84 neonatal MRI | Males born from supplemented mothers (600 mg of DHA) showed larger volume in total brain, cortex, whole gray matter and corpus callosum compared to controls. |
| A. Rifkin-Graboi [10], 2013 | Prospective observational study | Depression | 157 neonatal MRI | Changes in microstructure of the right amygdala. |
| C. Lebel [11], 2016 | Prospective observational study | Depression | 52 children MRI | altered gray matter structure in children. |
| Y. Wu [12], 2020 | Prospective cohort study | Anxiety | 193 fetal MRI | Reduced fetal hippocampal volume, particularly in the left hippocampus. Maternal anxiety and stress are associated with changes in fetal cortical gyrification index of the frontal and temporal lobes. |
| S. W. Jacobson [13], 2017 | Prospective observational study | Alcohol | 32 children MRI | Smaller CC in infants with FAS diagnoses. |
| A. Roos [14], 2021 | Prospective observational study | Alcohol | 83 children MRI | Alterations in white matter microstructural integrity in children with PAE. |
| C. Çetin [15], 2023 | Prospective observational study | Smoke | 250 fetal US | Significant reduction in second trimester measurement compared to non-exposed fetuses. |
| B. S. Peterson [16], 2015 | Cross-sectional study | PAH exposure | 40 children MRI | Reduction in the surface area of WM, predominantly confined to the left hemisphere of the brain. |
| R. L. Triplett [17], 2022 | Prospective longitudinal cohort study | Prenatal social disadvantage | 280 neonatal MRI | Reductions in WM, cortical gray matter, and subcortical gray matter volumes and cortical folding. |
| Y. Lu [18], 2021 | Prospective longitudinal cohort study | Socioeconomic status | 144 fetal MRI | Higher socioeconomic status is associated with:
|
| M. P. Herzberg [19], 2023 | Longitudinal observational study | Socioeconomic status | 241 neonatal MRI | Socioeconomic disadvantages are associated with higher cortisol level and smaller amygdala volumes. |
| Author, Year | Model | Investigated Variable | Main Findings |
|---|---|---|---|
| Belluscio L [20], 2014 | Mouse | Low protein diet | Delays in the surface righting reflex and negative geotaxis response. |
| Fragoso J [21], 2021 | Rat | Low protein diet | Reduction in the expression of BDNF in the hypothalamus and motor cortex. |
| Li C [22], 2017 | Baboon | Undernourished | Suppression of neurotrophic factors, dysregulated cell proliferation, and impaired glial maturation and neural synthesis. |
| Rajarethnem HT [23], 2017 | Rat | DHA supplementation | Significant increase in the number of hippocampal neural cells. |
| Fedorova I [24], 2009 | Rat | DHA | Low brain DHA is associated with a deficit in spatial reversal learning that could be related to changes in dopamine transmission in critical brain circuits. |
| Liu D [25], 2010 | Rat | Iodine | Hypothyroidism is associated with: -decrease in BDNF mRNA expression in the hippocampus -long-term memory deficits of pups. |
| Zhan Y [26], 2015 | Rat | Iodine | Hypothyroidism is associated with: -decreased activation of the CREB signaling pathway -impairments of cognitive function. |
| Tran P [27], 2015 | Rat | Iron Deficiency | Significant epigenetic modifications lead to long-term repression of BDNF. |
| Lien Y [28], 2019 | Rat | Iron Deficiency | Changes in DNA methylation in neural gene. |
| Barks A [29], 2022 | Rat | Iron Deficiency | TET/DNA hydroxymethylation system is disrupted in a brain region-specific manner. |
| Zhan X [30], 2012 | Rat | Folic Acid deficiency | Reduces neuronal cell proliferation by impairing mitosis and increases apoptosis. |
| Araki R [31], 2021 | Mouse | Low folate | Influences DNA and histone methylation, leading to the downregulation of neural gene expression and impaired fetal neurogenesis. |
| Gawlinska K [32], 2021 | Rat | High-fat diet | Changes in genes methylation, which are involved in synaptic function, chromatin remodeling and transcription regulation. |
| Gawlinska K [33], 2021 | Rat | High-fat diet | Alteration in mTOR and MAPK pathways are associated with autistic-like behavior. |
| Czarzasta K [34], 2019 | Rat | Depression | Altered levels of BDNF in the cerebellum and hippocampus have been associated with neurodevelopmental and behavioral delays in offspring. |
| Jones J [35], 2010 | Mouse | Prenatal stress | Combination of 5-HTT polymorphisms and prenatal stress increases the risk of ASD. |
| Matsui F [36], 2018 | Mouse | Prenatal stress | In mice with 5-HTT polymorphisms, dopamine levels increase significantly in the striatum. DHA supplementation reduces dopamine levels. |
| Chan Y [37], 2016 | Mouse | Smoke exposure | Smoke increases markers of hypoxia, oxidative stress and inflammation in neural cells, which may render dams and their offspring vulnerable to additional brain insults. |
| Jung S-Y [38], 2010 | Rat | Smoke exposure | Neuroligin-1 can modulate synaptic plasticity in the amygdala circuits of adult animals, likely by regulating the abundance of postsynaptic NMDA receptors. |
| Xiao M-F [39], 2009 | Mouse | Smoke exposure | NCAM is a modulator of the dopaminergic system, playing an important role in the etiology of psychiatric disorders. |
| Niedzwiedz-Massey V-M [40], 2021 | Mouse | Alcohol | Ethanol induces neuroinflammation, reduces the expression of molecules associated with mature oligodendrocytes, and leads to a decrease in genes expressed in oligodendrocyte progenitor cells. |
| Cantacorps L [41], 2019 | Mouse | Alcohol | Alcohol induces persistent epigenetic modifications, leading to long-term cognitive and behavioral impairments. |
| Milbocker K [42], 2022 | Rat | Alcohol | Alcohol leads to alterations in corpus callosum development and in myelination process. |
| Cui J [43], 2019 | Mouse | Air pollution | Activation of the dopamine pathway with the inhibition of glycine pathway can lead to locomotor hyperactivities. |
| Ehsanifar M [44], 2019 | Mouse | Air pollution | Alteration in the expression of pro-inflammatory cytokines and N-methyl-D-aspartate receptor subunits in the hippocampus, leads to increased anxiety and spatial memory dysfunction in offspring. |
| Nicolas Z M [45], 2017 | Mouse | Air pollution | Prenatal exposure to diesel exhaust particles leads to a significant upregulation of TLR4 expression and alterations in microglia activation, resulting in a higher vulnerability to behavioral deficits in male adult offspring. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubrano, C.; Parisi, F.; Cetin, I. Impact of Maternal Environment and Inflammation on Fetal Neurodevelopment. Antioxidants 2024, 13, 453. https://doi.org/10.3390/antiox13040453
Lubrano C, Parisi F, Cetin I. Impact of Maternal Environment and Inflammation on Fetal Neurodevelopment. Antioxidants. 2024; 13(4):453. https://doi.org/10.3390/antiox13040453
Chicago/Turabian StyleLubrano, Chiara, Francesca Parisi, and Irene Cetin. 2024. "Impact of Maternal Environment and Inflammation on Fetal Neurodevelopment" Antioxidants 13, no. 4: 453. https://doi.org/10.3390/antiox13040453
APA StyleLubrano, C., Parisi, F., & Cetin, I. (2024). Impact of Maternal Environment and Inflammation on Fetal Neurodevelopment. Antioxidants, 13(4), 453. https://doi.org/10.3390/antiox13040453