
Nrf2 Plays a Key Role in Erythropoiesis during Aging


 , ,
, , 
 ,
,  ,
,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Design of the Study
2.2. Flow Cytometric Analysis of Erythropoiesis
2.3. Analysis of Red Cells and Sorted Erythroid Precursors
2.3.1. Molecular Analysis of Erythroid Precursors
2.3.2. Immunoblot Analysis of Red Cells and Erythroid Precursors
2.4. Preparation and Characterization of Astaxanthin-Loaded PLGA Nanoparticles
2.5. Mass Spectrometric Analysis of Plasma and Organ ATS-NP Distribution
2.6. Splenic Macrophage Analysis and Erythrophagocytosis
2.7. Statistics
3. Results
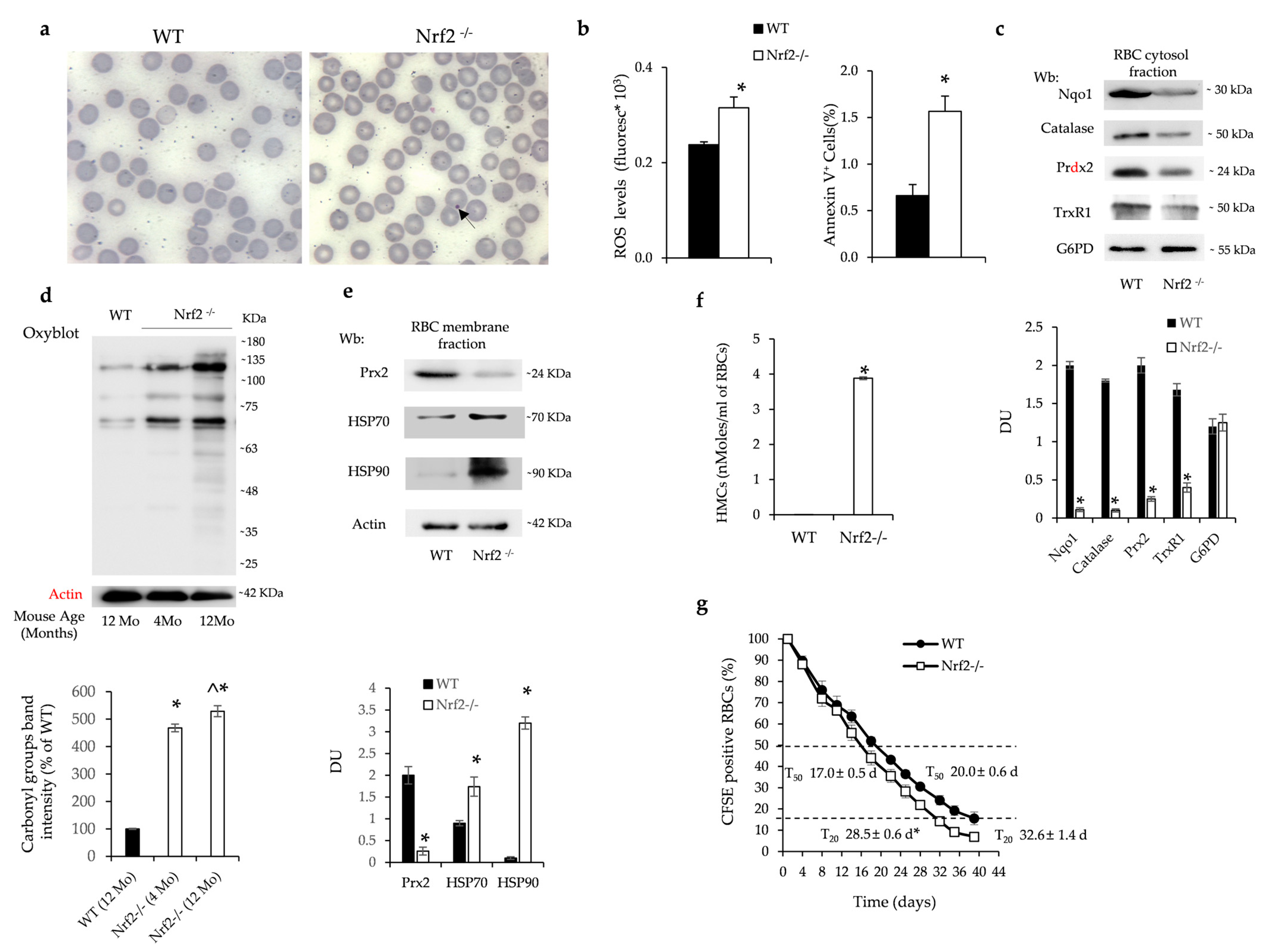
3.1. Nrf2−/− Mice Show Accelerated Red Cell Senescence, Downregulation of Nrf2/ARE Antioxidant Systems and Age-Dependent Hyporegenerative Anemia
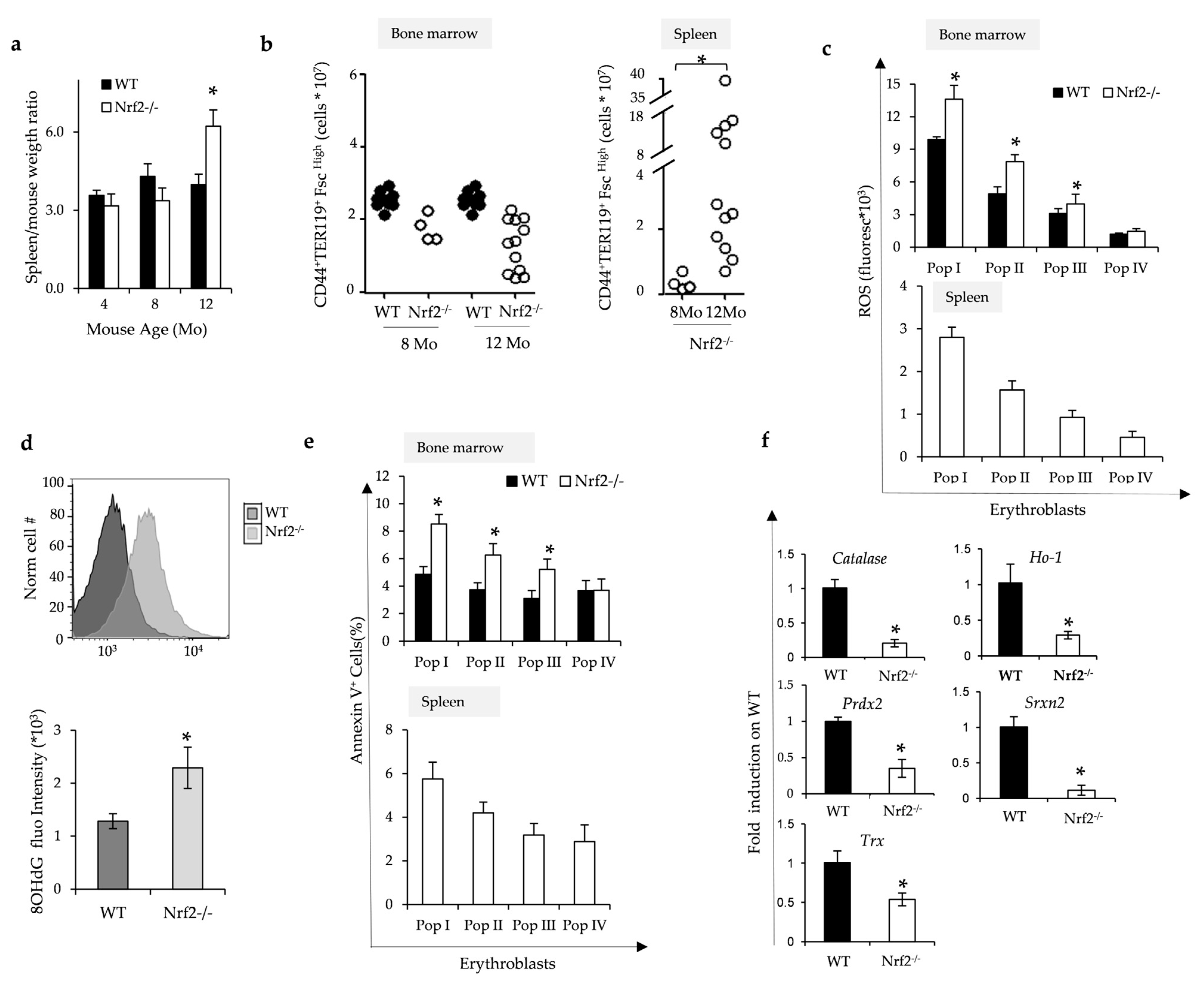
3.2. Nrf2−/− Mice Develop Age-Dependent Ineffective Erythropoiesis Associated with Oxidative Damage Due to Lack of Nrf2/ARE-Dependent Antioxidant Systems
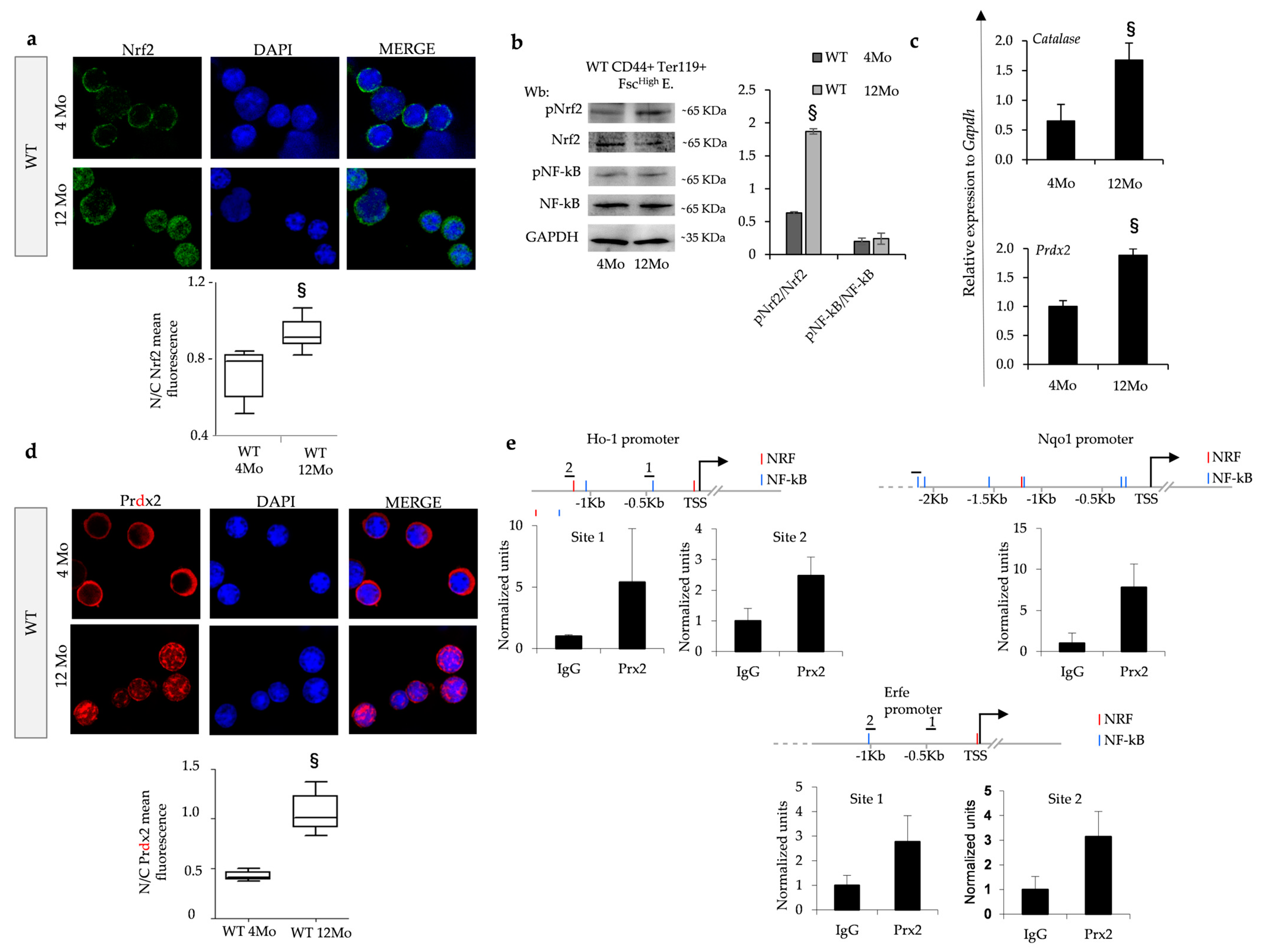
3.3. Nrf2 Is Activated during Erythropoiesis and Facilitated by Nuclear Translocation of Prdx2 during Aging
3.4. Nrf2−/− Mice Display Increased Sensitivity to Doxorubicin-Induced Stress Erythropoiesis
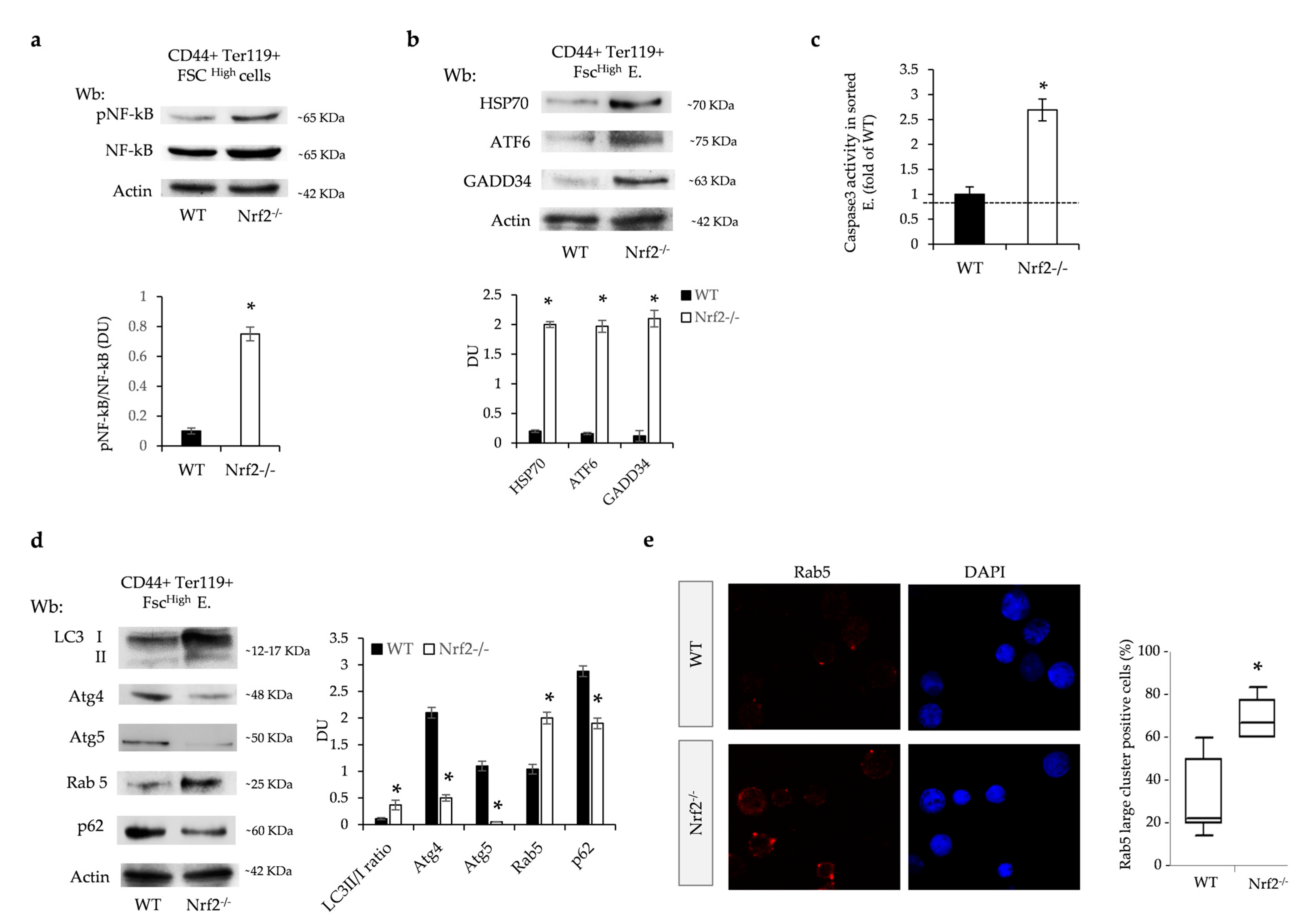
3.5. The Absence of Nrf2 Promotes Overactivation of UPR System and Impairment of Autophagy and Facilitates the Activation of Caspase-3 Pro-Apoptotic Pathway
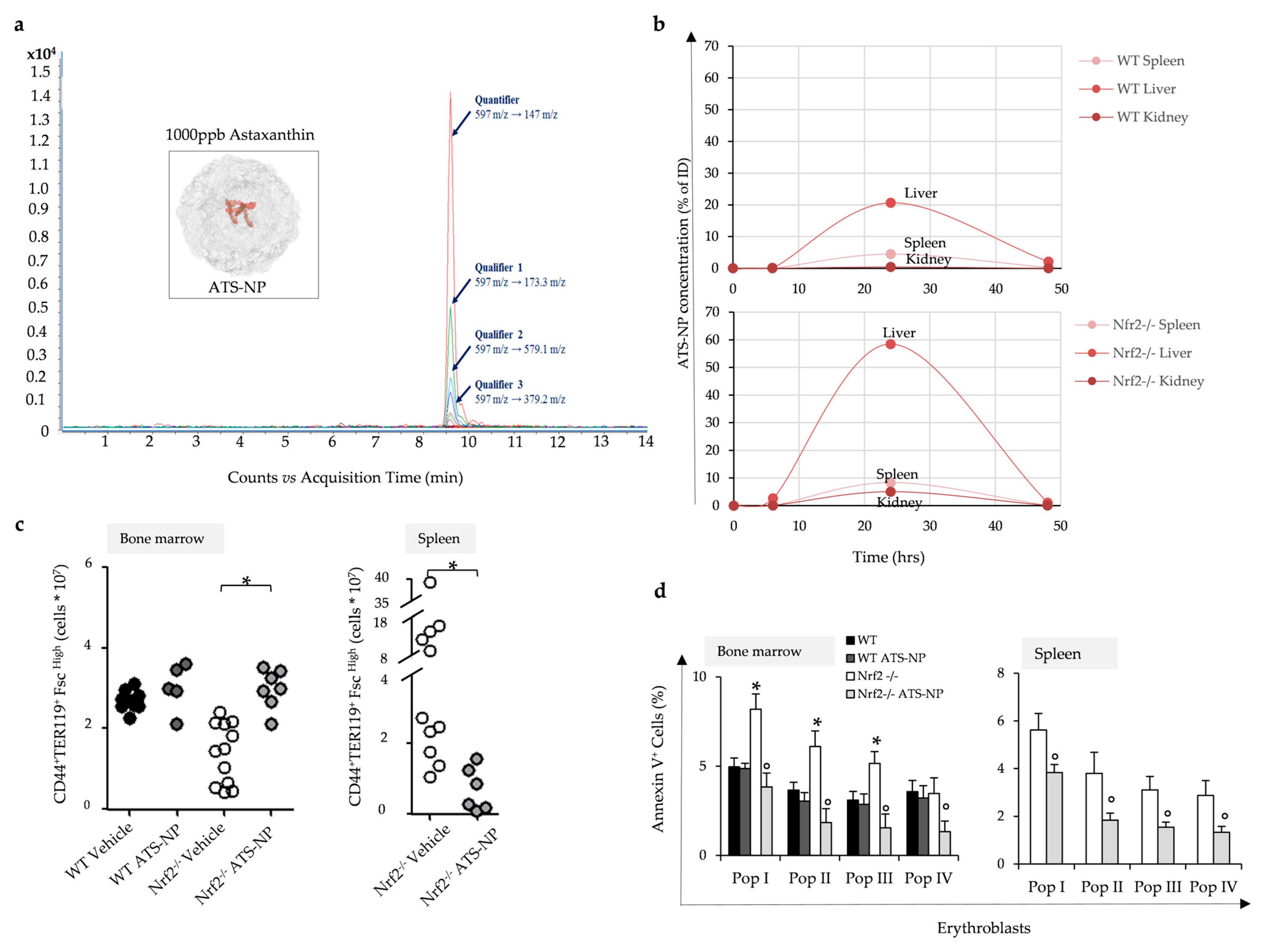
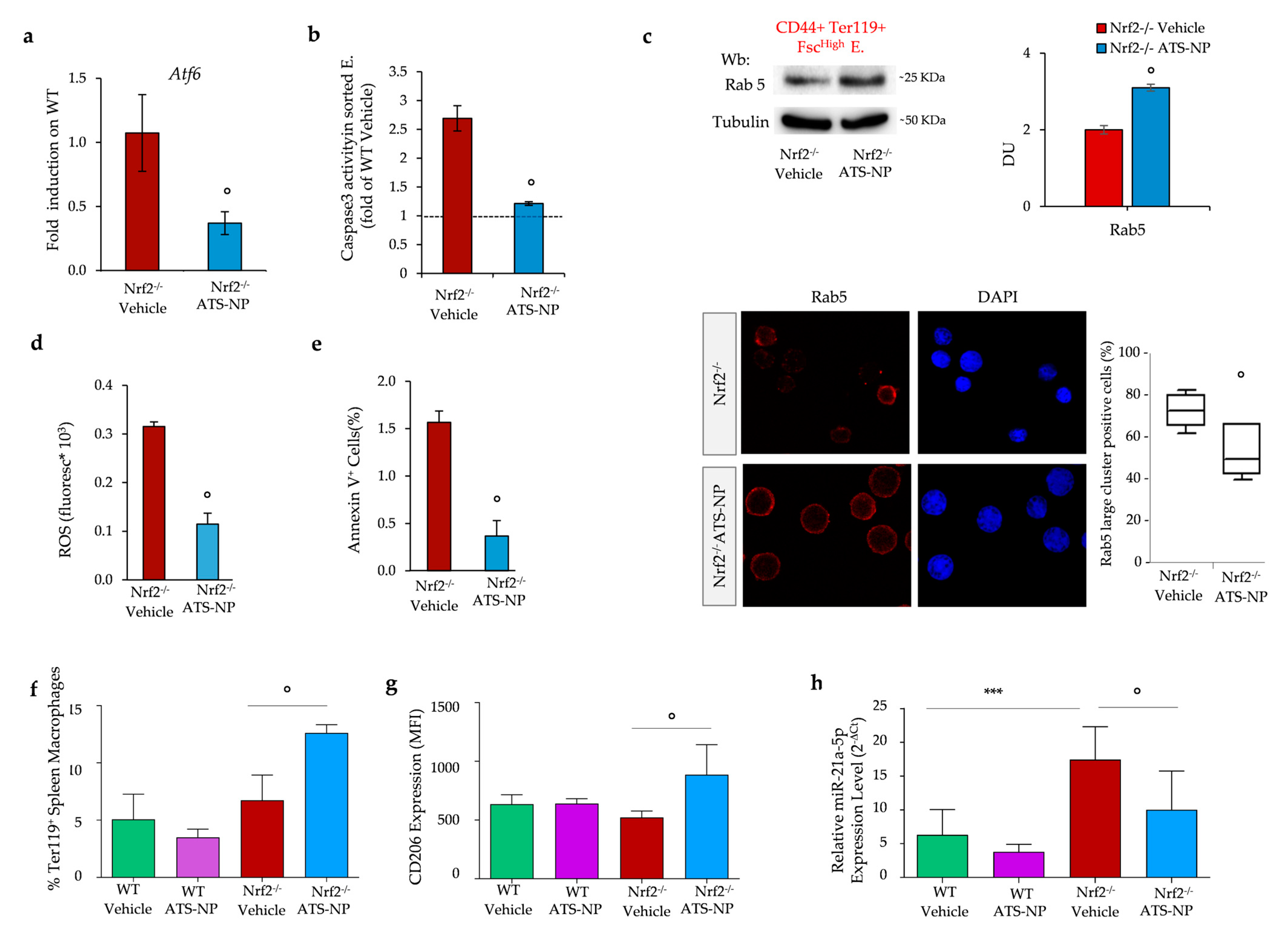
3.6. In Nrf2−/− Mice, Treatment with Astaxanthin PLGA Nanoparticles Enhances Erythropoiesis and Improves Anemia
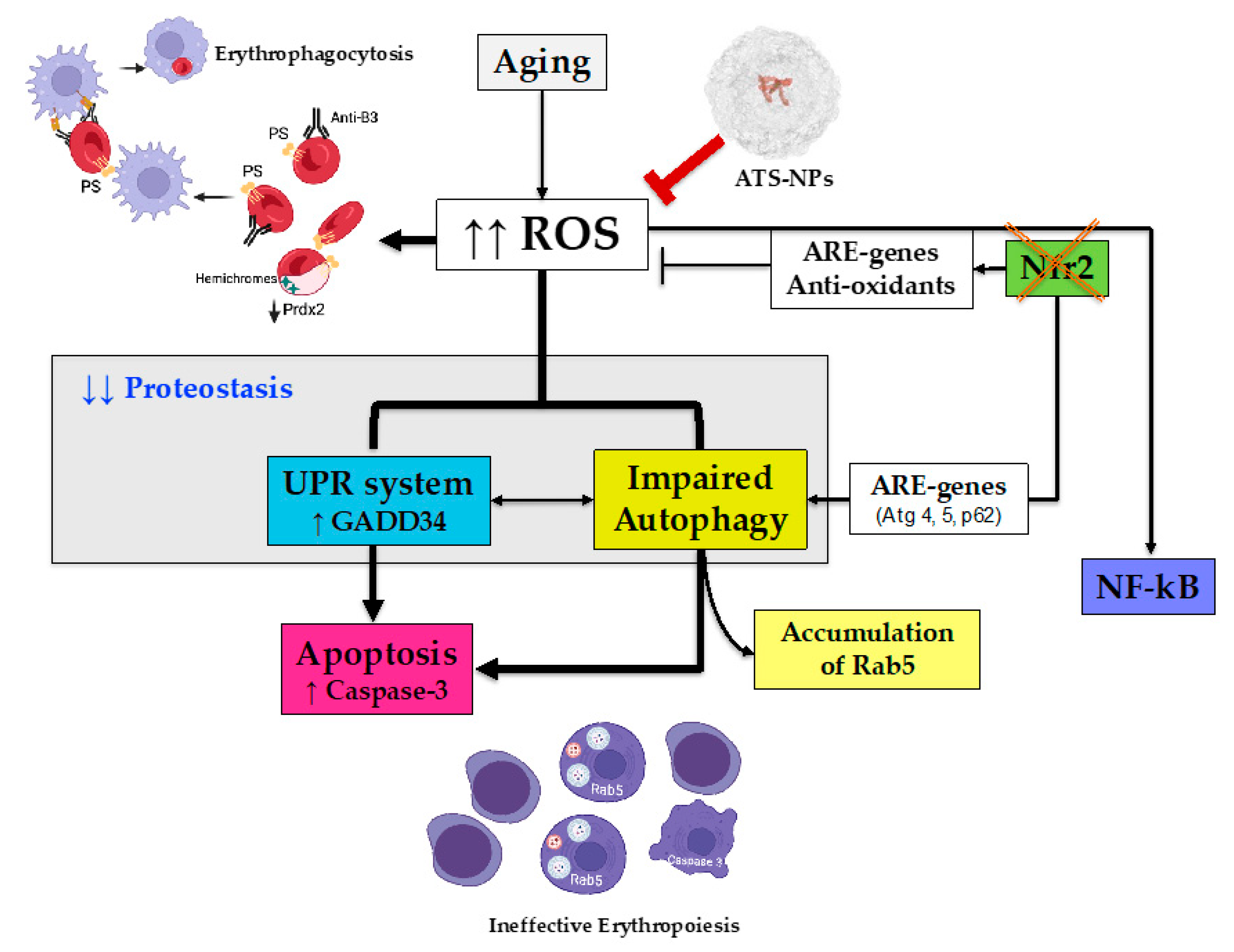
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Chan, K.; Kan, Y.W.; Johnson, J.A. Targeted disruption of Nrf2 causes regenerative immune-mediated hemolytic anemia. Proc. Natl. Acad. Sci. USA 2004, 101, 9751–9756. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Warabi, E.; Yanagawa, T. Novel roles of peroxiredoxins in inflammation, cancer and innate immunity. J. Clin. Biochem. Nutr. 2012, 50, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Leiser, S.F.; Miller, R.A. Nrf2 signaling, a mechanism for cellular stress resistance in long-lived mice. Mol. Cell Biol. 2010, 30, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Osburn, W.O.; Kensler, T.W. Nrf2 signaling: An adaptive response pathway for protection against environmental toxic insults. Mutat. Res. 2008, 659, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1-Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Zhu, H.; Itoh, K.; Yamamoto, M.; Zweier, J.L.; Li, Y. Role of Nrf2 signaling in regulation of antioxidants and phase 2 enzymes in cardiac fibroblasts: Protection against reactive oxygen and nitrogen species-induced cell injury. FEBS Lett. 2005, 579, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.Y.; Jin, S.H.; Cho, I.J.; Ki, S.H. Nrf2-ARE pathway regulates induction of Sestrin-2 expression. Free Radic. Biol. Med. 2012, 53, 834–841. [Google Scholar] [CrossRef]
- Chan, K.; Lu, R.; Chang, J.C.; Kan, Y.W. NRF2, a member of the NFE2 family of transcription factors, is not essential for murine erythropoiesis, growth, and development. Proc. Natl. Acad. Sci. USA 1996, 93, 13943–13948. [Google Scholar] [CrossRef]
- Matte, A.; De Falco, L.; Federti, E.; Cozzi, A.; Iolascon, A.; Levi, S.; Mohandas, N.; Zamo, A.; Bruno, M.; Lebouef, C.; et al. Peroxiredoxin-2: A Novel Regulator of Iron Homeostasis in Ineffective Erythropoiesis. Antioxid. Redox Signal. 2018, 28, 1–14. [Google Scholar] [CrossRef]
- Matte, A.; De Falco, L.; Iolascon, A.; Mohandas, N.; An, X.; Siciliano, A.; Leboeuf, C.; Janin, A.; Bruno, M.; Choi, S.Y.; et al. The Interplay Between Peroxiredoxin-2 and Nuclear Factor-Erythroid 2 Is Important in Limiting Oxidative Mediated Dysfunction in beta-Thalassemic Erythropoiesis. Antioxid. Redox Signal. 2015, 23, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of Aging: An Autophagic Perspective. Front. Endocrinol. 2018, 9, 790. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Minakshi, R.; Rahman, S.; Jan, A.T.; Archana, A.; Kim, J. Implications of aging and the endoplasmic reticulum unfolded protein response on the molecular modality of breast cancer. Exp. Mol. Med. 2017, 49, e389. [Google Scholar] [CrossRef] [PubMed]
- Cominacini, L.; Mozzini, C.; Garbin, U.; Pasini, A.; Stranieri, C.; Solani, E.; Vallerio, P.; Tinelli, I.A.; Fratta Pasini, A. Endoplasmic reticulum stress and Nrf2 signaling in cardiovascular diseases. Free Radic. Biol. Med. 2015, 88, 233–242. [Google Scholar] [CrossRef]
- Lupo, F.; Tibaldi, E.; Matte, A.; Sharma, A.K.; Brunati, A.M.; Alper, S.L.; Zancanaro, C.; Benati, D.; Siciliano, A.; Bertoldi, M.; et al. A new molecular link between defective autophagy and erythroid abnormalities in chorea-acanthocytosis. Blood 2016, 128, 2976–2987. [Google Scholar] [CrossRef] [PubMed]
- Khungwanmaythawee, K.; Sornjai, W.; Paemanee, A.; Jaratsittisin, J.; Fucharoen, S.; Svasti, S.; Lithanatudom, P.; Roytrakul, S.; Smith, D.R. Mitochondrial changes in β0-thalassemia/Hb E disease. PLoS ONE 2016, 11, e0153831. [Google Scholar] [CrossRef]
- Zhang, S.; Macias-Garcia, A.; Velazquez, J.; Paltrinieri, E.; Kaufman, R.J.; Chen, J.J. HRI coordinates translation by eIF2alphaP and mTORC1 to mitigate ineffective erythropoiesis in mice during iron deficiency. Blood 2018, 131, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Kamath, B.S.; Srikanta, B.M.; Dharmesh, S.M.; Sarada, R.; Ravishankar, G.A. Ulcer preventive and antioxidative properties of astaxanthin from Haematococcus pluvialis. Eur. J. Pharmacol. 2008, 590, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rogers, H.M.; Zhang, X.; Noguchi, C.T. Sex difference in mouse metabolic response to erythropoietin. FASEB J. 2017, 31, 2661–2673. [Google Scholar] [CrossRef] [PubMed]
- Casarrubea, D.; Viatte, L.; Hallas, T.; Vasanthakumar, A.; Eisenstein, R.S.; Schumann, K.; Hentze, M.W.; Galy, B. Abnormal body iron distribution and erythropoiesis in a novel mouse model with inducible gain of iron regulatory protein (IRP)-1 function. J. Mol. Med. 2013, 91, 871–881. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Beneduce, E.; Matte, A.; De Falco, L.; Mbiandjeu, S.; Chiabrando, D.; Tolosano, E.; Federti, E.; Petrillo, S.; Mohandas, N.; Siciliano, A.; et al. Fyn kinase is a novel modulator of erythropoietin signaling and stress erythropoiesis. Am. J. Hematol. 2019, 94, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, M.C.; Kobari, L.; Lapillonne, H.; Chalmers, D.; Kiger, L.; Cynober, T.; Marden, M.C.; Wajcman, H.; Douay, L. Ex vivo generation of fully mature human red blood cells from hematopoietic stem cells. Nat. Biotechnol. 2005, 23, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Kundrat, L.; Pishesha, N.; Bilate, A.; Theile, C.; Maruyama, T.; Dougan, S.K.; Ploegh, H.L.; Lodish, H.F. Engineered red blood cells as carriers for systemic delivery of a wide array of functional probes. Proc. Natl. Acad. Sci. USA 2014, 111, 10131–10136. [Google Scholar] [CrossRef] [PubMed]
- Matte, A.; Low, P.S.; Turrini, F.; Bertoldi, M.; Campanella, M.E.; Spano, D.; Pantaleo, A.; Siciliano, A.; De Franceschi, L. Peroxiredoxin-2 expression is increased in β-thalassemic mouse red cells but is displaced from the membrane as a marker of oxidative stress. Free Radic. Biol. Med. 2010, 49, 457–466. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.; Olivieri, O.; Miraglia del Giudice, E.; Perrotta, S.; Sabato, V.; Corrocher, R.; Iolascon, A. Membrane cation and anion transport activities in erythrocytes of hereditary spherocytosis: Effects of different membrane protein defects. Am. J. Hematol. 1997, 55, 121–128. [Google Scholar] [CrossRef]
- Xu, L.; Gao, Z.; Yang, Z.; Qu, M.; Li, H.; Chen, L.; Lv, Y.; Fan, Z.; Yue, W.; Li, C.; et al. Evaluation of Reliable Reference Genes for In Vitro Erythrocyte Generation from Cord Blood CD34+ Cells. DNA Cell Biol. 2021, 40, 1200–1210. [Google Scholar] [CrossRef]
- Makadia, H.K.; Siegel, S.J. Poly Lactic-co-Glycolic Acid (PLGA) as Biodegradable Controlled Drug Delivery Carrier. Polymers 2011, 3, 1377–1397. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Luo, W.; Chen, I.; Chen, Y.; Alkam, D.; Wang, Y.; Semenza, G.L. PRDX2 and PRDX4 are negative regulators of hypoxia-inducible factors under conditions of prolonged hypoxia. Oncotarget 2016, 7, 6379–6397. [Google Scholar] [CrossRef] [PubMed]
- Leak, R.K.; Zhang, L.; Luo, Y.; Li, P.; Zhao, H.; Liu, X.; Ling, F.; Jia, J.; Chen, J.; Ji, X. Peroxiredoxin 2 battles poly(ADP-ribose) polymerase 1- and p53-dependent prodeath pathways after ischemic injury. Stroke 2013, 44, 1124–1134. [Google Scholar] [CrossRef]
- Gan, Y.; Ji, X.; Hu, X.; Luo, Y.; Zhang, L.; Li, P.; Liu, X.; Yan, F.; Vosler, P.; Gao, Y.; et al. Transgenic overexpression of peroxiredoxin-2 attenuates ischemic neuronal injury via suppression of a redox-sensitive pro-death signaling pathway. Antioxid. Redox Signal. 2012, 17, 719–732. [Google Scholar] [CrossRef]
- Rodier, F.; Munoz, D.P.; Teachenor, R.; Chu, V.; Le, O.; Bhaumik, D.; Coppe, J.P.; Campeau, E.; Beausejour, C.M.; Kim, S.H.; et al. DNA-SCARS: Distinct nuclear structures that sustain damage-induced senescence growth arrest and inflammatory cytokine secretion. J. Cell Sci. 2011, 124, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.; Ordonez, R.; Reiter, R.J.; Gonzalez-Gallego, J.; Mauriz, J.L. Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 2015, 59, 292–307. [Google Scholar] [CrossRef]
- Inami, Y.; Waguri, S.; Sakamoto, A.; Kouno, T.; Nakada, K.; Hino, O.; Watanabe, S.; Ando, J.; Iwadate, M.; Yamamoto, M.; et al. Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J. Cell Biol. 2011, 193, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Hama, S.; Uenishi, S.; Yamada, A.; Ohgita, T.; Tsuchiya, H.; Yamashita, E.; Kogure, K. Scavenging of hydroxyl radicals in aqueous solution by astaxanthin encapsulated in liposomes. Biol. Pharm. Bull. 2012, 35, 2238–2242. [Google Scholar] [CrossRef]
- Donoso, A.; Gonzalez-Duran, J.; Munoz, A.A.; Gonzalez, P.A.; Agurto-Munoz, C. Therapeutic uses of natural astaxanthin: An evidence-based review focused on human clinical trials. Pharmacol. Res. 2021, 166, 105479. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef]
- Augusti, P.R.; Quatrin, A.; Somacal, S.; Conterato, G.M.; Sobieski, R.; Ruviaro, A.R.; Maurer, L.H.; Duarte, M.M.; Roehrs, M.; Emanuelli, T. Astaxanthin prevents changes in the activities of thioredoxin reductase and paraoxonase in hypercholesterolemic rabbits. J. Clin. Biochem. Nutr. 2012, 51, 42–49. [Google Scholar] [CrossRef]
- Genc, Y.; Bardakci, H.; Yucel, C.; Karatoprak, G.S.; Kupeli Akkol, E.; Hakan Barak, T.; Sobarzo-Sanchez, E. Oxidative Stress and Marine Carotenoids: Application by Using Nanoformulations. Mar. Drugs 2020, 18, 423. [Google Scholar] [CrossRef] [PubMed]
- Nabar, G.M.; Mahajan, K.D.; Calhoun, M.A.; Duong, A.D.; Souva, M.S.; Xu, J.; Czeisler, C.; Puduvalli, V.K.; Otero, J.J.; Wyslouzil, B.E.; et al. Micelle-templated, poly(lactic-co-glycolic acid) nanoparticles for hydrophobic drug delivery. Int. J. Nanomed. 2018, 13, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, R.; Madhyastha, H.; Nurrahmah, Q.I.; Purbasari, B.; Maruyama, M.; Nakajima, Y. MicroRNA 21 Elicits a Pro-inflammatory Response in Macrophages, with Exosomes Functioning as Delivery Vehicles. Inflammation 2021, 44, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Aumann, K.; Frey, A.V.; May, A.M.; Hauschke, D.; Kreutz, C.; Marx, J.P.; Timmer, J.; Werner, M.; Pahl, H.L. Subcellular mislocalization of the transcription factor NF-E2 in erythroid cells discriminates prefibrotic primary myelofibrosis from essential thrombocythemia. Blood 2013, 122, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, H.; Yan, W.; Xu, L.; Wang, X.; Zhao, X.; Yang, X.; Chen, G.; Ji, Y. Disruption of Nrf2 enhances upregulation of nuclear factor-kappaB activity, proinflammatory cytokines, and intercellular adhesion molecule-1 in the brain after traumatic brain injury. Mediat. Inflamm. 2008, 2008, 725174. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Guo, L.; Yang, Y.; Wang, Y.; Xia, S.; Gong, H.; Zhang, B.K.; Yan, M. Dissecting the Crosstalk Between Nrf2 and NF-kappaB Response Pathways in Drug-Induced Toxicity. Front. Cell Dev. Biol. 2021, 9, 809952. [Google Scholar] [CrossRef]
- Bolati, D.; Shimizu, H.; Yisireyili, M.; Nishijima, F.; Niwa, T. Indoxyl sulfate, a uremic toxin, downregulates renal expression of Nrf2 through activation of NF-kappaB. BMC Nephrol. 2013, 14, 56. [Google Scholar] [CrossRef]
- Hamza, E.; Vallejo-Mudarra, M.; Ouled-Haddou, H.; Garcia-Caballero, C.; Guerrero-Hue, M.; Santier, L.; Rayego-Mateos, S.; Larabi, I.A.; Alvarez, J.C.; Garcon, L.; et al. Indoxyl sulfate impairs erythropoiesis at BFU-E stage in chronic kidney disease. Cell Signal 2023, 104, 110583. [Google Scholar] [CrossRef] [PubMed]
- Kautz, L.; Jung, G.; Valore, E.V.; Rivella, S.; Nemeth, E.; Ganz, T. Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat. Genet. 2014, 46, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Coffey, R.; Ganz, T. Erythroferrone: An Erythroid Regulator of Hepcidin and Iron Metabolism. Hemasphere 2018, 2, e35. [Google Scholar] [CrossRef]
- Matte, A.; Federti, E.; Winter, M.; Koerner, A.; Harmeier, A.; Mazer, N.; Tomka, T.; Di Paolo, M.L.; Defalco, L.; Andolfo, I.; et al. Bitopertin, a selective oral GLYT1 inhibitor, improves anemia in a mouse model of β-thalassemia. JCI Insight 2019, 4, e130111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kaufman, R.J. The unfolded protein response: A stress signaling pathway critical for health and disease. Neurology 2006, 66, S102–S109. [Google Scholar] [CrossRef]
- Rhee, S.G.; Bae, S.H. The antioxidant function of sestrins is mediated by promotion of autophagic degradation of Keap1 and Nrf2 activation and by inhibition of mTORC1. Free Radic. Biol. Med. 2015, 88, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Jimenez-Moreno, N.; Garcia-Yague, A.J.; Escoll, M.; de Ceballos, M.L.; Van Leuven, F.; Rabano, A.; Yamamoto, M.; Rojo, A.I.; Cuadrado, A. Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 2016, 12, 1902–1916. [Google Scholar] [CrossRef] [PubMed]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Lamark, T.; Sjottem, E.; Larsen, K.B.; Awuh, J.A.; Overvatn, A.; McMahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [PubMed]
- Sarad, K.; Stefanska, M.; Kraszewska, I.; Szade, K.; Sluimer, J.C.; Blyszczuk, P.; Dulak, J.; Jazwa-Kusior, A. Single-cell transcriptomics reveals subtype-specific molecular profiles in Nrf2-deficient macrophages from murine atherosclerotic aortas. Front. Immunol. 2023, 14, 1249379. [Google Scholar] [CrossRef]
- Ruotsalainen, A.K.; Inkala, M.; Partanen, M.E.; Lappalainen, J.P.; Kansanen, E.; Makinen, P.I.; Heinonen, S.E.; Laitinen, H.M.; Heikkila, J.; Vatanen, T.; et al. The absence of macrophage Nrf2 promotes early atherogenesis. Cardiovasc. Res. 2013, 98, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Santoro, R.; Zanotto, M.; Carbone, C.; Piro, G.; Tortora, G.; Melisi, D. MEKK3 Sustains EMT and Stemness in Pancreatic Cancer by Regulating YAP and TAZ Transcriptional Activity. Anticancer Res. 2018, 38, 1937–1946. [Google Scholar] [CrossRef]
- Colman-Martinez, M.; Martinez-Huelamo, M.; Miralles, E.; Estruch, R.; Lamuela-Raventos, R.M. A New Method to Simultaneously Quantify the Antioxidants: Carotenes, Xanthophylls, and Vitamin A in Human Plasma. Oxid. Med. Cell. Longev. 2015, 2015, 9268531. [Google Scholar] [CrossRef]
- Dattilo, M.; Fontanarosa, C.; Spinelli, M.; Bini, V.; Amoresano, A. Modulation of Human Hydrogen Sulfide Metabolism by Micronutrients, Preliminary Data. Nutr. Metab. Insights 2022, 15, 11786388211065372. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5’→ 3’) | Reverse Primer Sequence (5’→ 3’) |
|---|---|---|
| Gapdh | CCACATCGCTCAGACACCAT | AGTTAAAAGCAGCCCTGGTGAC |
| Erfe Atf6 P62 Txn Srxn2 Prdx2 Ho-1 | ATGGGGCTGGAGAACAGC GCCCGGTGAATGGAAAACTT AGCGGTTACTCACTCCATGGAAGA CATGGCCAACAAAATCATGCAGCT TTTGGCAGAGACAT CGCCTAGTCCAGGCCTTTCCAA GCACAGGGTGACAGAAGAG | TGGCATTGTCCAAGAAGACA GTCTCCTCAGCACAGCGATA CTTGGGGAGGTTTCGTCTCT CGATCTGTTCAATTTTCGTTGGG ACTTTCAGCGTGGCTGGG GATGGTGTCACTGCCGGG GTCAGCATCACCTGCAGCTC |
| Wild-Type Mice | |||
|---|---|---|---|
| 4-Month-Old Mice (n = 6) | 8-Month-Old Mice (n = 6) | 12-Month-Old Mice (n = 6) | |
| Hct (%) | 46.1 ± 1.4 | 45.9 ± 0.7 | 44.8 ± 0.2 |
| Hb (g/dL) | 14.8 ± 0.5 | 15.0 ± 0.1 | 14.3 ± 0.4 |
| MCV (fl) | 51.3 ± 0.2 | 51.0 ± 0.1 | 52.2 ± 0.3 |
| MCH (pg) | 15.9 ± 0.7 | 16.5 ± 0.3 | 15.6 ± 0.2 |
| RDW (%) | 12.4 ± 0.08 | 13.5 ± 0.1 | 12.7 ± 0.3 |
| Retics (103 cells/uL) | 450 ± 22 | 431 ± 51 | 248 ± 24 ° |
| MCVr (fl) | 54.9 ± 2 | 56.7 ± 3 | 59.9 ± 1.8 ° |
| Nrf2−/− Mice | |||
| 4-Month-Old Mice (n = 6) | 8-Month-Old Mice (n = 6) | 12-MonthsOld Mice (n = 6) | |
| Hct (%) | 44.3 ± 0.8 | 41.8 ± 1.1 °* | 33.6 ± 3 °* |
| Hb (g/dL) | 13.2 ± 0.5 | 12.0 ± 0.2 °* | 11.0 ± 0.5 °* |
| MCV (fl) | 51.8 ± 1.5 | 50.0 ± 2.0 | 57.2 ± 1.3 °* |
| MCH (pg) | 16.7 ± 1.1 | 16.0 ± 0.3 | 16.1 ± 0.4 |
| RDW (%) | 13.9 ± 0.55 | 13.2 ± 0.4 | 14.1 ± 0.4 °* |
| Retics (103 cells/uL) | 380 ± 20 * | 190 ± 59 °* | 180 ± 12 °* |
| MCVr (fl) | 61.2 ± 1.3 * | 61.0 ± 1.4 * | 65.0 ± 0.2 °* |
| Vehicle Wild-Type Mice (n = 6) | ATS-NP Wild-Type Mice (n = 6) | Vehicle Nrf2−/− Mice (n = 6) | ATS-NP Nrf2−/− Mice (n = 5) | |
|---|---|---|---|---|
| Hct (%) | 44.8 ± 0.2 | 45.9 ± 0.7 | 33.6 ± 3 * | 43.5 ± 0.9 *^ |
| Hb (g/dL) | 14.3 ± 0.4 | 15.0 ± 0.1 | 11.0 ± 0.5 * | 14.0 ± 0.4 ^ |
| MCV (fl) | 52.2 ± 0.3 | 51.0 ± 0.1 | 57.2 ± 1.3 * | 53.5 ± 1.1 ^ |
| MCH (pg) | 15.6 ± 0.2 | 16.5 ± 0.3 | 16.1 ± 0.4 | 16.3 ± 0.2 |
| RDW (%) | 12.7 ± 0.3 | 13.5 ± 0.1 | 14.1 ± 0.4 * | 12.6 ± 0.4 ^ |
| Retics (103 cells/uL) | 248 ± 24 | 431 ± 51 ^ | 180 ± 12 * | 355 ± 65 ^ |
| MCVr (fl) | 59.9 ± 1.8 | 56.1 ± 2.0 | 65.0 ± 0.2 * | 62.0 ± 1.6 ^ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbiandjeu, S.C.T.; Siciliano, A.; Mattè, A.; Federti, E.; Perduca, M.; Melisi, D.; Andolfo, I.; Amoresano, A.; Iolascon, A.; Valenti, M.T.; et al. Nrf2 Plays a Key Role in Erythropoiesis during Aging. Antioxidants 2024, 13, 454. https://doi.org/10.3390/antiox13040454
Mbiandjeu SCT, Siciliano A, Mattè A, Federti E, Perduca M, Melisi D, Andolfo I, Amoresano A, Iolascon A, Valenti MT, et al. Nrf2 Plays a Key Role in Erythropoiesis during Aging. Antioxidants. 2024; 13(4):454. https://doi.org/10.3390/antiox13040454
Chicago/Turabian StyleMbiandjeu, Serge Cedrick Toya, Angela Siciliano, Alessandro Mattè, Enrica Federti, Massimiliano Perduca, Davide Melisi, Immacolata Andolfo, Angela Amoresano, Achille Iolascon, Maria Teresa Valenti, and et al. 2024. "Nrf2 Plays a Key Role in Erythropoiesis during Aging" Antioxidants 13, no. 4: 454. https://doi.org/10.3390/antiox13040454
APA StyleMbiandjeu, S. C. T., Siciliano, A., Mattè, A., Federti, E., Perduca, M., Melisi, D., Andolfo, I., Amoresano, A., Iolascon, A., Valenti, M. T., Turrini, F., Bovi, M., Pisani, A., Recchiuti, A., Mattoscio, D., Riccardi, V., Dalle Carbonare, L., Brugnara, C., Mohandas, N., & De Franceschi, L. (2024). Nrf2 Plays a Key Role in Erythropoiesis during Aging. Antioxidants, 13(4), 454. https://doi.org/10.3390/antiox13040454