
Obesity Is Associated with Oxidative Stress Markers and Antioxidant Enzyme Activity in Mexican Children

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Anthropometric Measurements
2.3. Blood Sample and Biochemical Measurements
2.4. Measurement of Oxidative Stress Markers
2.4.1. Malondialdehyde (MDA)
2.4.2. Protein Carbonylation
2.5. Measurement of Antioxidant Enzyme Activity
2.5.1. Determination of Total Protein Content
2.5.2. Superoxide Dismutase-Specific (SOD) Enzyme Activity
2.5.3. Catalase-Specific (CAT) Enzyme Activity
2.5.4. Glutathione Peroxidase-Specific (GPx) Enzyme Activity
2.6. Data Analysis
3. Results
3.1. General Characteristics of the Study Sample
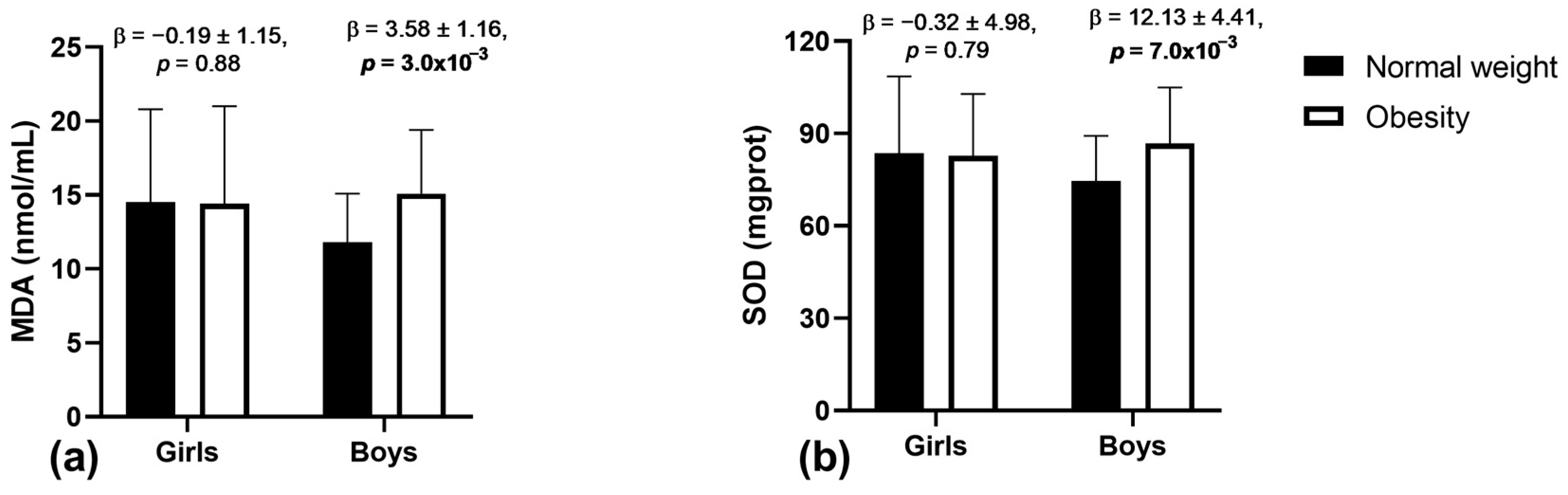
3.2. Associations of Obesity with Oxidative Stress Markers and Antioxidant Enzyme Activity
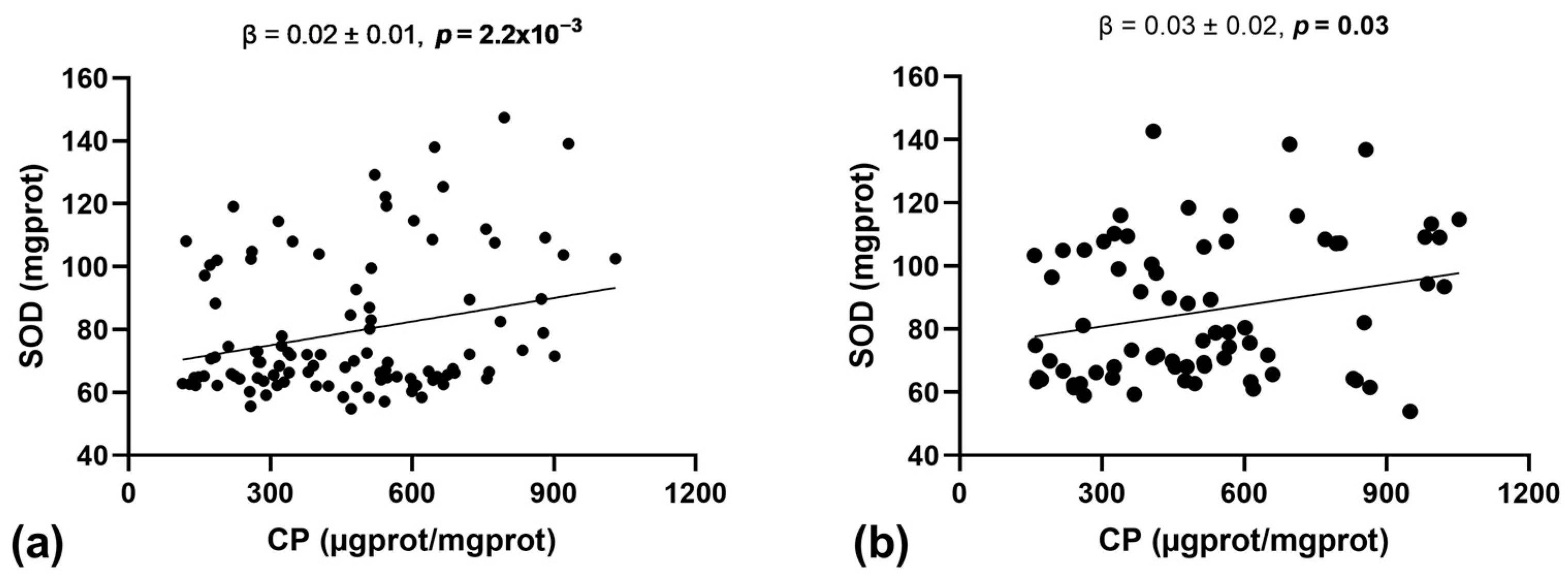
3.3. Association between Oxidative Stress Markers and Antioxidant Enzyme Activity
3.4. Associations between Cardiometabolic Traits, Oxidative Stress Markers, and Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Trait | Shapiro–Wilk Test (p-Value) | |
|---|---|---|
| Untransformed | Transformed | |
| Carbonylated proteins (µgprot/mgprot) | 7.4 × 10−6 | 0.550 |
| Lipoperoxidation (nmol/L) | 2.3 × 10−15 | 0.991 |
| Superoxide dismutase (mgprot) | 3.2 × 10−10 | 0.583 |
| Catalase (mgprot) | 7.7 × 10−12 | 0.813 |
| Glutathione peroxidase (U/mL) | 7.3 × 10−14 | 0.276 |
References
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Reilly, J.J.; Kelly, J. Long-term impact of overweight and obesity in childhood and adolescence on morbidity and premature mortality in adulthood: Systematic review. Int. J. Obes. 2011, 35, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, K.R.; Redden, D.T.; Wang, C.; Westfall, A.O.; Allison, D.B. Years of life lost due to obesity. JAMA 2003, 289, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Shamah-Levy, T.; Gaona-Pineda, E.B.; Cuevas-Nasu, L.; Morales-Ruan, C.; Valenzuela-Bravo, D.G.; Humaran, I.M.; Ávila-Arcos, M.A. Prevalencias de sobrepeso y obesidad en población escolar y adolescente de México. Ensanut Continua 2020–2022. Salud Publica Mexico 2023, 65, S218–S224. [Google Scholar] [CrossRef] [PubMed]
- Reddon, H.; Gueant, J.L.; Meyre, D. The importance of gene-environment interactions in human obesity. Clin. Sci. 2016, 130, 1571–1597. [Google Scholar] [CrossRef] [PubMed]
- Raut, S.K.; Khullar, M. Oxidative stress in metabolic diseases: Current scenario and therapeutic relevance. Mol. Cell Biochem. 2023, 478, 185–196. [Google Scholar] [CrossRef]
- Masenga, S.K.; Kabwe, L.S.; Chakulya, M.; Kirabo, A. Mechanisms of Oxidative Stress in Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 7898. [Google Scholar] [CrossRef] [PubMed]
- Balan, A.I.; Halatiu, V.B.; Scridon, A. Oxidative Stress, Inflammation, and Mitochondrial Dysfunction: A Link between Obesity and Atrial Fibrillation. Antioxidants 2024, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef]
- Frohnert, B.I.; Sinaiko, A.R.; Serrot, F.J.; Foncea, R.E.; Moran, A.; Ikramuddin, S.; Choudry, U.; Bernlohr, D.A. Increased adipose protein carbonylation in human obesity. Obesity 2011, 19, 1735–1741. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.C.; Arrais, R.F.; Almeida, M.G.; Souza, Z.M.; Pedrosa, L.F. Plasma lipid profile and lipid peroxidation in overweight or obese children and adolescents. J. Pediatr. 2004, 80, 23–28. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, Y.G.; Zhang, S.M.; Wang, J.Y.; Xiao, W.Q.; Wang, X.Y.; Zhou, J.F. Overweight and obesity-induced oxidative stress in children. Biomed. Environ. Sci. 2006, 19, 353–359. [Google Scholar] [PubMed]
- Faienza, M.F.; Francavilla, R.; Goffredo, R.; Ventura, A.; Marzano, F.; Panzarino, G.; Marinelli, G.; Cavallo, L.; Di Bitonto, G. Oxidative stress in obesity and metabolic syndrome in children and adolescents. Horm. Res. Paediatr. 2012, 78, 158–164. [Google Scholar] [CrossRef]
- Kilic, E.; Özer, Ö.F.; Erek, A.T.; Erman, H.; Torun, E.; Ayhan, S.K.; Caglar, H.G.; Selek, S.; Kocyigit, A. Oxidative Stress Status in Childhood Obesity: A Potential Risk Predictor. Med. Sci. Monit. 2016, 22, 3673–3679. [Google Scholar] [CrossRef] [PubMed]
- Marcela, A.S.; Tarraf, H.S.; Nicoletti, C.F.; Percário, S.; Oliveira, B.A.; Oliveira, C.C.; Gregório, M.L.; Pinhanelli, V.C.; Dorotéia, R.S. Oxidative Stress and Biochemical Parameters in Childhood Obesity. Austin. J. Nutri. Food Sci. 2018, 6, 1097. [Google Scholar]
- Lechuga-Sancho, A.M.; Gallego-Andujar, D.; Ruiz-Ocaña, P.; Visiedo, F.M.; Saez-Benito, A.; Schwarz, M.; Segundo, C.; Mateos, R.M. Obesity induced alterations in redox homeostasis and oxidative stress are present from an early age. PLoS ONE 2018, 13, e0191547. [Google Scholar] [CrossRef]
- Ustundag, B.; Gungor, S.; Aygun, A.D.; Turgut, M.; Yilmaz, E. Oxidative status and serum leptin levels in obese prepubertal children. Cell Biochem. Funct. 2007, 25, 479–483. [Google Scholar] [CrossRef]
- Sfar, S.; Boussoffara, R.; Sfar, M.T.; Kerkeni, A. Antioxidant enzymes activities in obese Tunisian children. Nutr. J. 2013, 12, 18. [Google Scholar] [CrossRef]
- Habib, S.A.; Saad, E.A.; Elsharkawy, A.A.; Attia, Z.R. Pro-inflammatory adipocytokines, oxidative stress, insulin, Zn and Cu: Interrelations with obesity in Egyptian non-diabetic obese children and adolescents. Adv. Med. Sci. 2015, 60, 179–185. [Google Scholar] [CrossRef]
- Fuentes-Venado, C.E.; Terán-Pérez, G.; Espinosa-Hernández, V.M.; Martínez-Herrera, E.; Segura-Uribe, J.J.; Mercadillo, R.E.; Pinto-Almazán, R.; Guerra-Araiza, C. Nutritional Status Influences Oxidative Stress and Insulin Resistance in Preschool Children. Metab. Syndr. Relat. Disord. 2021, 19, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Erdeve, O.; Siklar, Z.; Kocaturk, P.A.; Dallar, Y.; Kavas, G.O. Antioxidant superoxide dismutase activity in obese children. Biol. Trace Elem. Res. 2004, 98, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Flegal, K.M.; Cole, T.J. Construction of LMS Parameters for the Centers for Disease Control and Prevention 2000 Growth Charts; National Center for Health Statistics: Hyattsville, MD, USA, 2013; pp. 1–3.
- Kuczmarski, R.J.; Ogden, C.L.; Guo, S.S.; Grummer-Strawn, L.M.; Flegal, K.M.; Mei, Z.; Wei, R.; Curtin, L.; Roche, A.; Johnson, C.; et al. 2000 CDC Growth Charts for the United States: Methods and Development; National Center for Health Statistics: Hyattsville, MD, USA, 2002; pp. 1–190.
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-dinitrophenylhydrazine spectrophotometric assay for quantification of carbonyls in oxidized proteins. Anal. Biochem. 2014, 458, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Medina, J.P.; Zenteno-Savin, T.; Elsner, R. Antioxidant enzymes in ringed seal tissues: Potential protection against dive-associated ischemia/reperfusion. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Feise, R.J. Do multiple outcome measures require p-value adjustment? BMC Med. Res. Methodol. 2002, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2019, 54, 287–293. [Google Scholar] [CrossRef]
- Alkazemi, D.; Rahman, A.; Habra, B. Alterations in glutathione redox homeostasis among adolescents with obesity and anemia. Sci. Rep. 2021, 11, 3034. [Google Scholar] [CrossRef]
- Codoner-Franch, P.; Boix-Garcia, L.; Simo-Jorda, R.; Del Castillo-Villaescusa, C.; Maset-Maldonado, J.; Valls-Belles, V. Is obesity associated with oxidative stress in children? Int. J. Pediatr. Obes. 2010, 5, 56–63. [Google Scholar] [CrossRef]
- Mohseni, R.; Arab Sadeghabadi, Z.; Goodarzi, M.T.; Teimouri, M.; Nourbakhsh, M.; Razzaghy Azar, M. Evaluation of Mn-superoxide dismutase and catalase gene expression in childhood obesity: Its association with insulin resistance. J. Pediatr. Endocrinol. Metab. 2018, 31, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Amos, D.L.; Robinson, T.; Massie, M.B.; Cook, C.; Hoffsted, A.; Crain, C.; Santanam, N. Catalase overexpression modulates metabolic parameters in a new ‘stress-less’ leptin-deficient mouse model. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2293–2306. [Google Scholar] [CrossRef] [PubMed]
- Najafi, A.; Pourfarzam, M.; Zadhoush, F. Oxidant/antioxidant status in Type-2 diabetes mellitus patients with metabolic syndrome. J. Res. Med. Sci. 2021, 26, 6. [Google Scholar] [PubMed]
- Sobieska, M.; Gajewska, E.; Kalmus, G.; Samborski, W. Obesity, physical fitness, and inflammatory markers in Polish children. Med. Sci. Monit. 2013, 19, 493–500. [Google Scholar] [PubMed]
- Gómez-Pérez, Y.; Amengual-Cladera, E.; Català-Niell, A.; Thomàs-Moyà, E.; Gianotti, M.; Proenza, A.M.; Llado, I. Gender dimorphism in high-fat-diet-induced insulin resistance in skeletal muscle of aged rats. Cell Physiol. Biochem. 2008, 22, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Thomas-Moya, E.; Gomez-Perez, Y.; Fiol, M.; Gianotti, M.; Llado, I.; Proenza, A.M. Gender related differences in paraoxonase 1 response to high-fat diet-induced oxidative stress. Obesity 2008, 16, 2232–2238. [Google Scholar] [CrossRef]
- Vasconcelos, R.P.; Peixoto, M.S.; de Oliveira, K.A.; Ferreira, A.C.; Coelho-de-Souza, A.N.; Carvalho, D.P.; de Oliveira, A.C.; Fortunato, R.S. Sex differences in subcutaneous adipose tissue redox homeostasis and inflammation markers in control and high-fat diet fed rats. Appl. Physiol. Nutr. Metab. 2019, 44, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.A.; Kerr, C.J.; Whiting, P.; Finer, N.; McEneny, J.; Ashton, T. Oxidant stress in healthy normal-weight, overweight, and obese individuals. Obesity 2009, 17, 460–466. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Gómez-Hermosillo, L.; Casillas-Moreno, J.; Pacheco-Moisés, F.; Campos-Bayardo, T.I.; Román-Rojas, D.; Miranda-Díaz, A.G. Prevalence of Hypertension and Obesity: Profile of Mitochondrial Function and Markers of Inflammation and Oxidative Stress. Antioxidants 2023, 12, 165. [Google Scholar] [CrossRef]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of Oxidative Stress in Biological Samples Using the Thiobarbituric Acid Reactive Substances Assay. J. Vis. Exp. 2020, 159, e6112. [Google Scholar] [CrossRef]
- Onyango, A.N. Cellular Stresses and Stress Responses in the Pathogenesis of Insulin Resistance. Oxid. Med. Cell Longev. 2018, 2018, 4321714. [Google Scholar] [CrossRef] [PubMed]
- Revin, V.V.; Gromova, N.V.; Revina, E.S.; Samonova, A.Y.; Tychkov, A.Y.; Bochkareva, S.S.; Moskovkin, A.A.; Kuzmenko, T.P. The Influence of Oxidative Stress and Natural Antioxidants on Morphometric Parameters of Red Blood Cells, the Hemoglobin Oxygen Binding Capacity, and the Activity of Antioxidant Enzymes. Biomed. Res. Int. 2019, 2019, 2109269. [Google Scholar] [CrossRef] [PubMed]
- Van Gaal, L.F.; Vertommen, J.; De Leeuw, I.H. The in vitro oxidizability of lipoprotein particles in obese and non-obese subjects. Atherosclerosis 1998, 137, S39–S44. [Google Scholar] [CrossRef] [PubMed]
- Berliner, J.A.; Territo, M.; Navab, M.; Andalibi, A.; Parhami, F.; Liao, F.; Kim, J.; Estworthy, S.; Lusis, A.J.; Fogelman, A.M. Minimally modified lipoproteins in diabetes. Diabetes 1992, 41 (Suppl. S2), 74–76. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Tang, J.; Chen, X. Sensitive determination of malondialdehyde in rat prostate by high performance liquid chromatography with fluorescence detection. Sci. Rep. 2020, 10, 3990. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, V.; Cena, H.; Rossi, V.; Santero, S.; Bianchi, A.; Zuccotti, G. Ultra-Processed Food, Reward System and Childhood Obesity. Children 2023, 10, 804. [Google Scholar] [CrossRef]
- Calcaterra, V.; Cena, H.; Magenes, V.C.; Vincenti, A.; Comola, G.; Beretta, A.; Di Napoli, I.; Zuccotti, G. Sugar-Sweetened Beverages and Metabolic Risk in Children and Adolescents with Obesity: A Narrative Review. Nutrients 2023, 15, 702. [Google Scholar] [CrossRef]


| Traits | Normal Weight n = 120 | Obese n = 81 | p-Value |
|---|---|---|---|
| Girls/Boys, n (%) | 68 (56.7)/52 (43.3) | 35 (43.2)/46 (56.8) | 0.06 |
| Age, years | 8.63 ± 1.58 | 8.77 ± 1.54 | 0.52 |
| Body mass index, kg/m2 | 16.86 ± 1.75 | 24.15 ± 2.49 | 1.48 × 10−47 |
| Systolic blood pressure, mmHg | 100.14 ± 9.01 | 109.53 ± 10.34 | 1.08 × 10−10 |
| Diastolic blood pressure, mmHg | 62.32 ± 7.37 | 69.19 ± 8.83 | 1.04 × 10−8 |
| Total cholesterol, mg/dL | 157.53 ± 28.58 | 164.25 ± 28.35 | 0.10 |
| High-density lipoprotein cholesterol, mg/dL | 52.54 ± 11.63 | 45.70 ± 10.33 | 3.0 × 10−5 |
| Low-density lipoprotein cholesterol, mg/dL | 89.95 ± 20.93 | 99.32 ± 19.71 | 0.02 |
| Triglycerides, mg/dL | 86.95 ± 41.92 | 130.72 ± 62.87 | 2.03 × 10−7 |
| Fasting plasma glucose, mg/dL | 77.67 ± 6.71 | 76.93 ± 6.43 | 0.444 |
| Fasting plasma insulin, μU/mL | 5.27 ± 3.32 | 10.27 ± 7.01 | 3.71 × 10−8 |
| HOMA-IR | 1.03 ± 0.69 | 1.95 ± 1.44 | 6.86 × 10−7 |
| Carbonylated proteins, µgprot/mgprot | 484.22 ± 246.05 | 514.69 ± 253.24 | 0.39 |
| malondialdehyde, nmol/L | 13.33 ± 5.36 | 14.83 ± 6.97 | 0.09 |
| Superoxide dismutase, mgprot | 79.79 ± 21.94 | 85.05 ± 22.08 | 0.11 |
| Catalase, mgprot | 0.12 ± 0.10 | 0.18 ± 0.11 | 3.0 × 10−3 |
| Glutathione peroxidase, U/mL | 0.05 ± 0.06 | 0.19 ± 0.11 | 2.12 × 10−19 |
| Trait/Sample | Main Effect (a) | Interaction (b) | ||
|---|---|---|---|---|
| Obesity | p-Value | Obesity × Sex | p-Value | |
| CPs (µgprot/mgprot) | 33.77 ± 36.47 | 0.35 | 6.26 ± 18.13 | 0.73 |
| MDA (nmol/L) | 1.71 ± 0.89 | 0.05 | 0.92 ± 0.43 | 0.03 |
| SOD (mgprot) | 5.79 ± 3.34 | 0.08 | 3.41 ± 1.64 | 0.04 |
| CAT (mgprot) | 0.05 ± 0.01 | 5.0 × 10−3 | 0.01 ± 0.01 | 0.95 |
| GPx (U/mL) | 0.13 ± 0.01 | 3.7 × 10−19 | 0.01 ± 0.01 | 0.51 |
| Trait | SOD (mgprot) | CAT (mgprot) | GPx (U/mL) |
|---|---|---|---|
| CPs (µgprot/mgprot) | 0.23 (1.7 × 10−3) | 0.07 (0.35) | 0.04 (0.58) |
| MDA (nmol/L) | 0.07 (0.36) | 0.04 (0.56) | −0.02 (0.78) |
| Trait | CPs | Lip | SOD | CAT | GPx |
|---|---|---|---|---|---|
| Systolic blood pressure, mmHg | 0.06 (0.38) | 0.08 (0.32) | 0.09 (0.32) | −0.01 (0.81) | −0.07 (0.33) |
| Diastolic blood pressure, mmHg | 0.16 (0.04) | 0.07 (0.31) | 0.08 (0.31) | 0.05 (0.50) | −0.09 (0.25) |
| Total cholesterol, mg/dL | 0.12 (0.13) | −0.03 (0.67) | 0.11 (0.16) | 0.04 (0.55) | 0.01 (0.95) |
| High-density lipoprotein cholesterol, mg/dL | 0.10 (0.18) | −0.03 (0.64) | 0.14 (0.07) | 0.02 (0.77) | 0.12 (0.11) |
| Low-density lipoprotein cholesterol, mg/dL | 0.09 (0.25) | −0.01 (0.82) | 0.05 (0.46) | 0.04 (0.59) | −0.04 (0.62) |
| Triglycerides, mg/dL | 0.04 (0.57) | −0.06 (0.44) | 0.03 (0.66) | 0.01 (0.99) | −0.02 (0.76) |
| Fasting plasma glucose, mg/dL | 0.03 (0.65) | 0.01 (0.82) | 0.02 (0.76) | −0.17 (0.03) | −0.05 (0.51) |
| Fasting plasma insulin, μU/mL | 0.09 (0.24) | −0.10 (0.17) | 0.24 (2.0 × 10−3) | −0.12 (0.12) | −0.18 (0.02) |
| HOMA-IR | 008 (0.32) | −0.10 (0.18) | 0.23 (3.0 × 10−3) | −0.13 (0.08) | −0.17 (0.03) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cota-Magaña, A.I.; Vazquez-Moreno, M.; Rocha-Aguado, A.; Ángeles-Mejía, S.; Valladares-Salgado, A.; Díaz-Flores, M.; López-Díazguerrero, N.E.; Cruz, M. Obesity Is Associated with Oxidative Stress Markers and Antioxidant Enzyme Activity in Mexican Children. Antioxidants 2024, 13, 457. https://doi.org/10.3390/antiox13040457
Cota-Magaña AI, Vazquez-Moreno M, Rocha-Aguado A, Ángeles-Mejía S, Valladares-Salgado A, Díaz-Flores M, López-Díazguerrero NE, Cruz M. Obesity Is Associated with Oxidative Stress Markers and Antioxidant Enzyme Activity in Mexican Children. Antioxidants. 2024; 13(4):457. https://doi.org/10.3390/antiox13040457
Chicago/Turabian StyleCota-Magaña, Ana Isabel, Miguel Vazquez-Moreno, Andrés Rocha-Aguado, Selene Ángeles-Mejía, Adán Valladares-Salgado, Margarita Díaz-Flores, Norma Edith López-Díazguerrero, and Miguel Cruz. 2024. "Obesity Is Associated with Oxidative Stress Markers and Antioxidant Enzyme Activity in Mexican Children" Antioxidants 13, no. 4: 457. https://doi.org/10.3390/antiox13040457
APA StyleCota-Magaña, A. I., Vazquez-Moreno, M., Rocha-Aguado, A., Ángeles-Mejía, S., Valladares-Salgado, A., Díaz-Flores, M., López-Díazguerrero, N. E., & Cruz, M. (2024). Obesity Is Associated with Oxidative Stress Markers and Antioxidant Enzyme Activity in Mexican Children. Antioxidants, 13(4), 457. https://doi.org/10.3390/antiox13040457