
Tributyrin Supplementation Rescues Chronic–Binge Ethanol-Induced Oxidative Stress in the Gut–Lung Axis in Mice

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chronic–Binge Ethanol Feeding Model
2.2. Shotgun Metagenomics Sequencing, Bioinformatics, and Statistical Analysis
2.3. Small Intestinal Isolation of Intraepithelial Lymphocytes, Lamina Propria Lymphocytes, and Intestinal Epithelial Cells
2.4. Isolation of Lung Immune Cells
2.5. Flow Cytometry Analysis of IELs, LPLs, and Lung Immune Cells
2.6. Tissue RT-qPCR
2.7. Western Blotting: Intestinal Epithelial Cells
2.8. Plasma Endotoxin Quantification Using Limulus Amoebocyte Lysate (LAL) Assay
2.9. Cecal Secretory IgA (SIgA) Enzyme-Linked Immunoassay (ELISA)
2.10. Lipocalin 2 (LCN2) and Myeloperoxidase (MPO) ELISA in Lung Tissue
2.11. Plating of Peyer’s Patches on Enterococcosel Agar
2.12. Statistical Analysis
3. Results
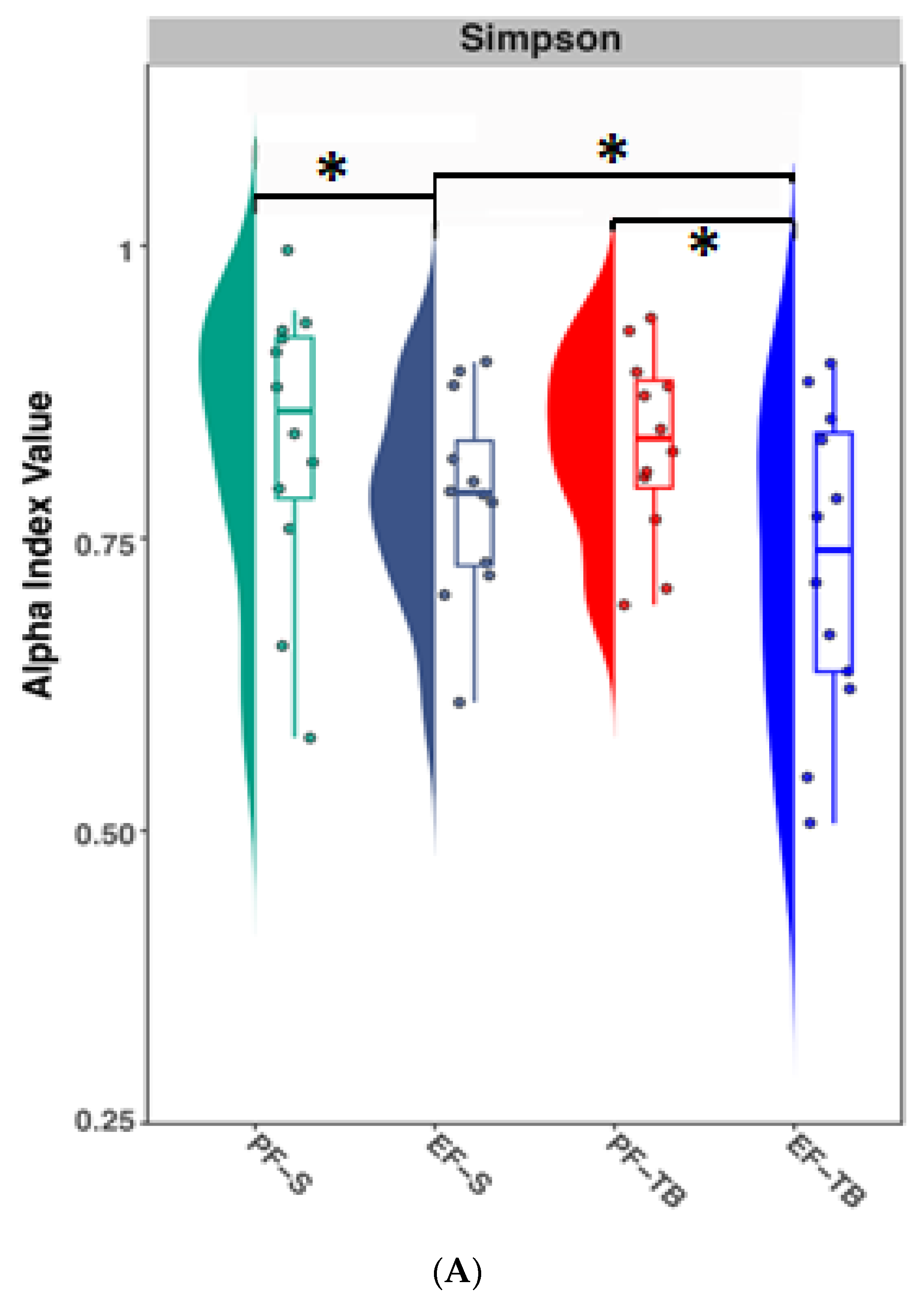
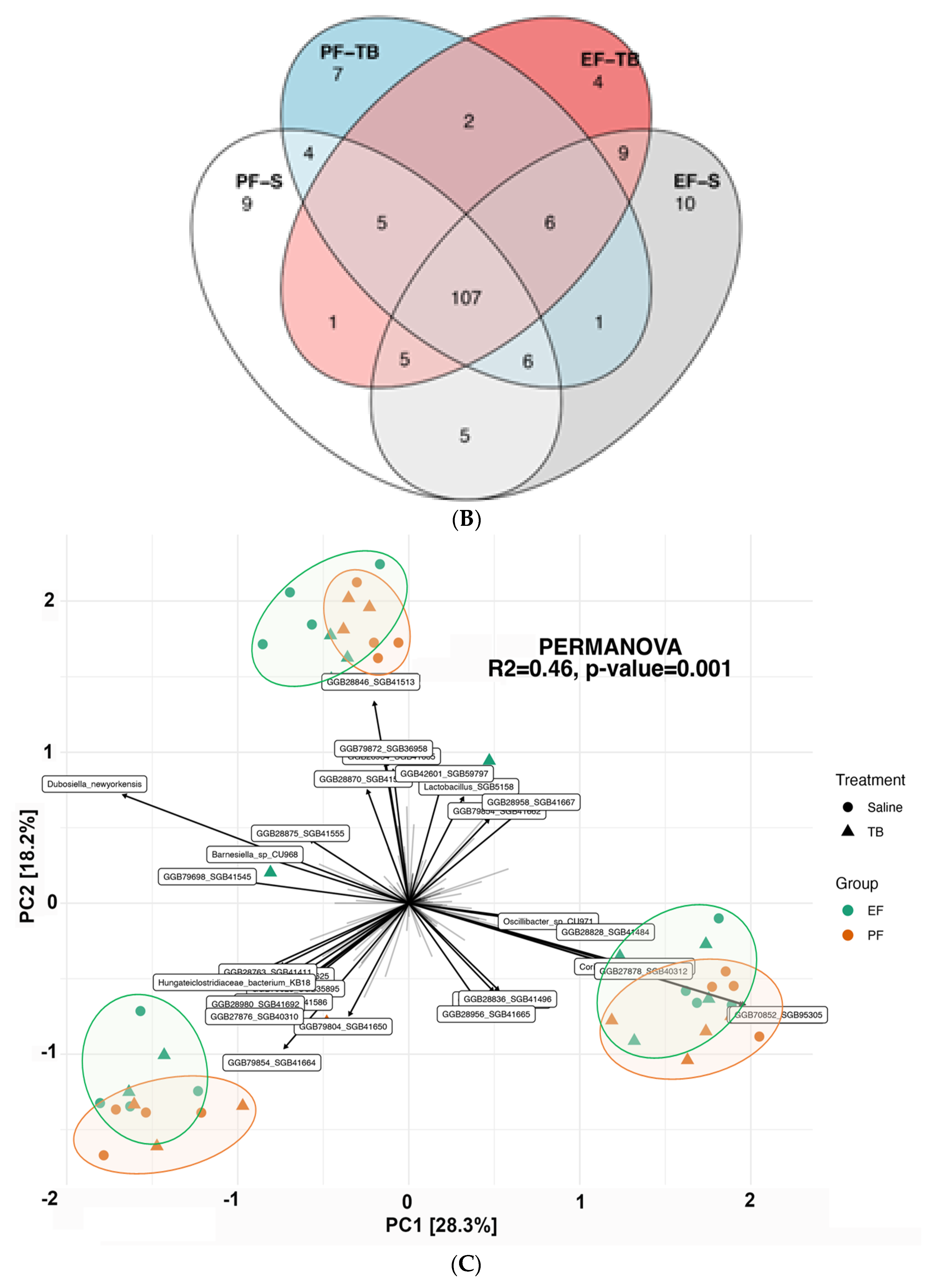
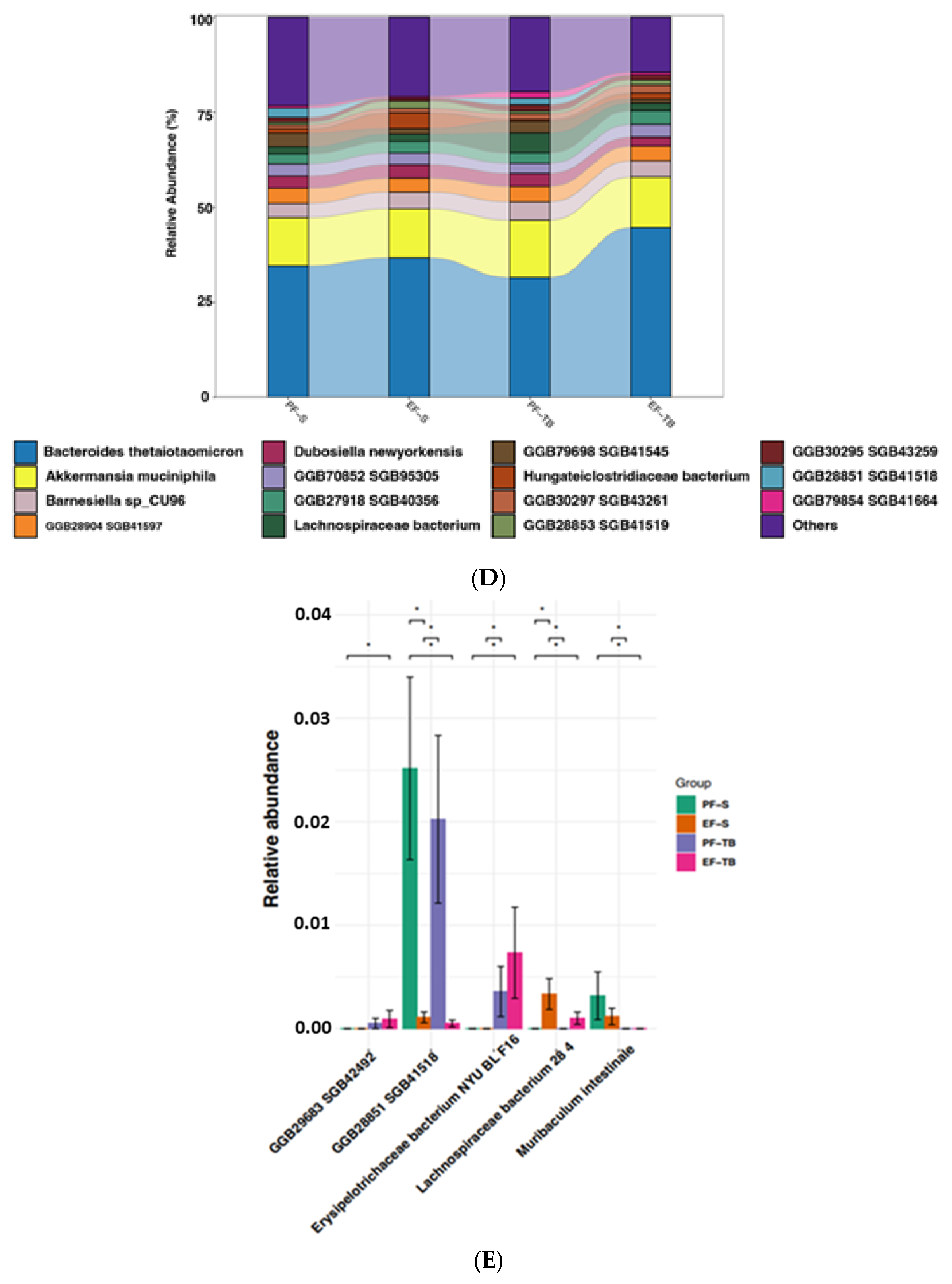
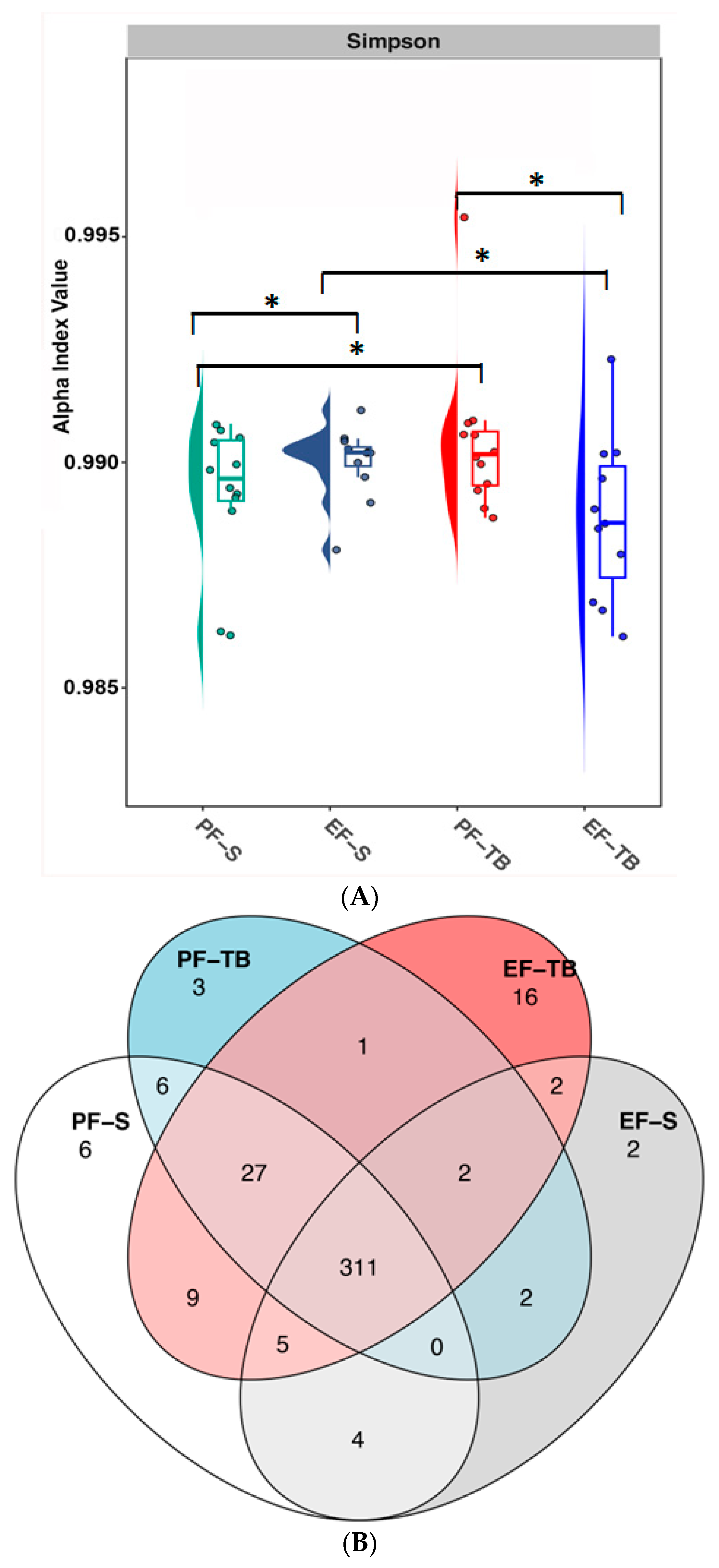
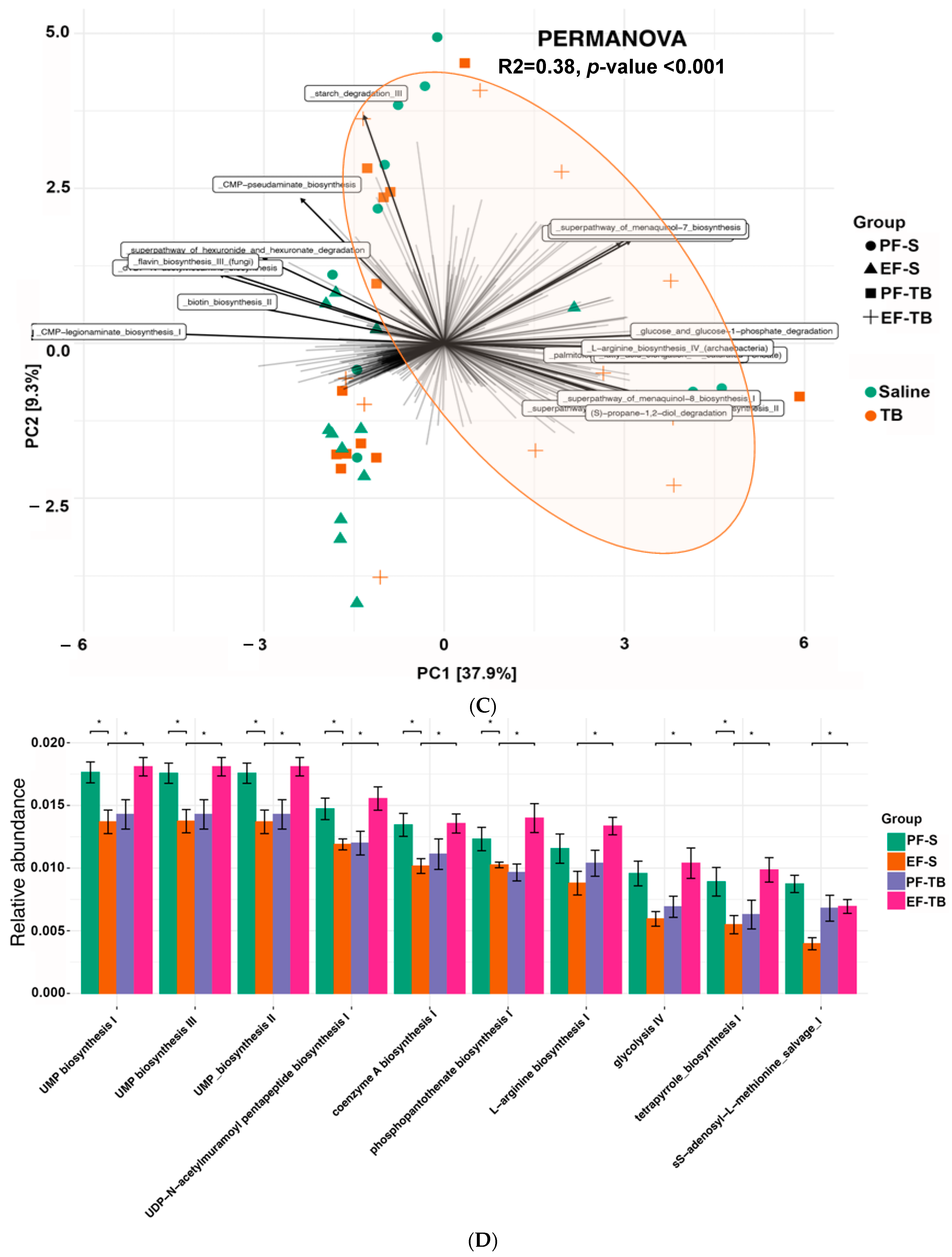
3.1. Ethanol Exposure and Tributyrin Supplementation Alter the Gut Microbiome and Its Functional Pathways
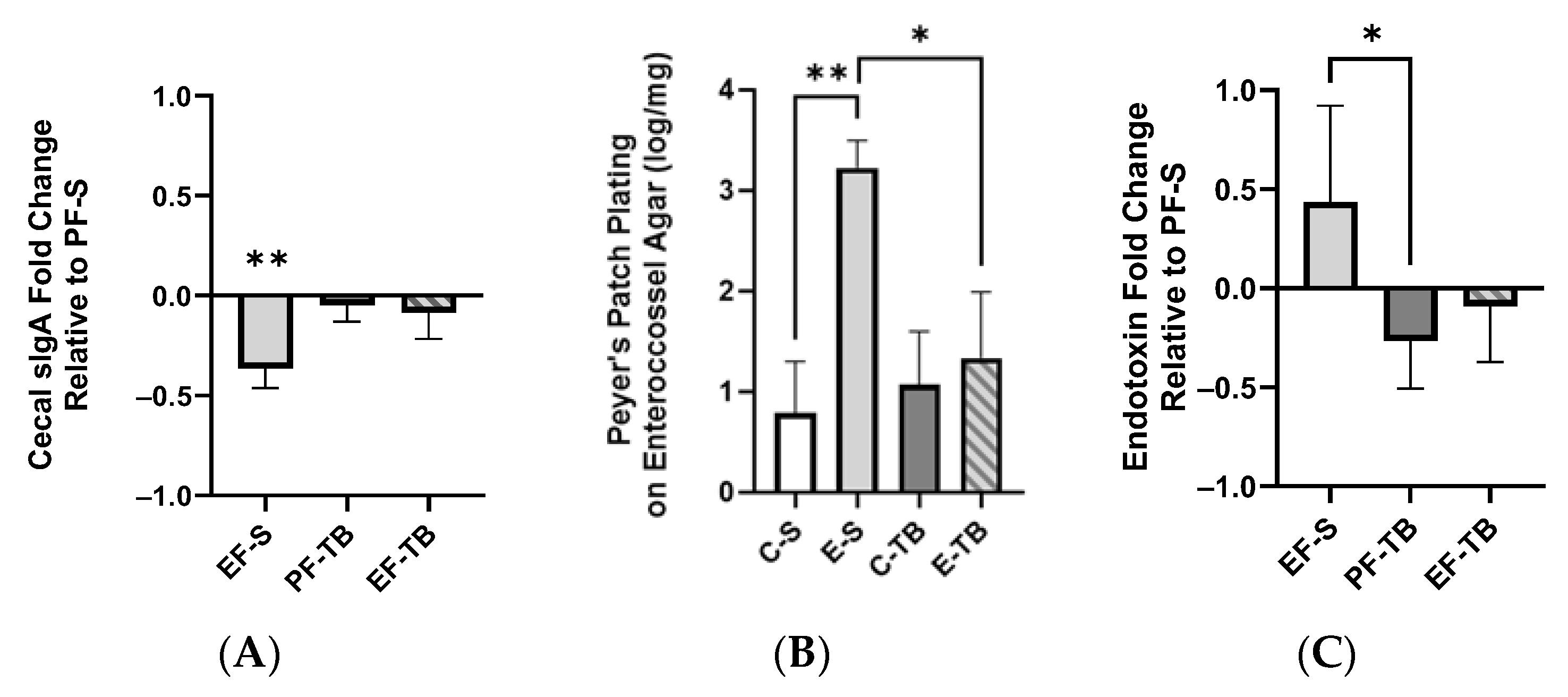
3.2. Ethanol Affects Intestinal Epithelial Immune Responses
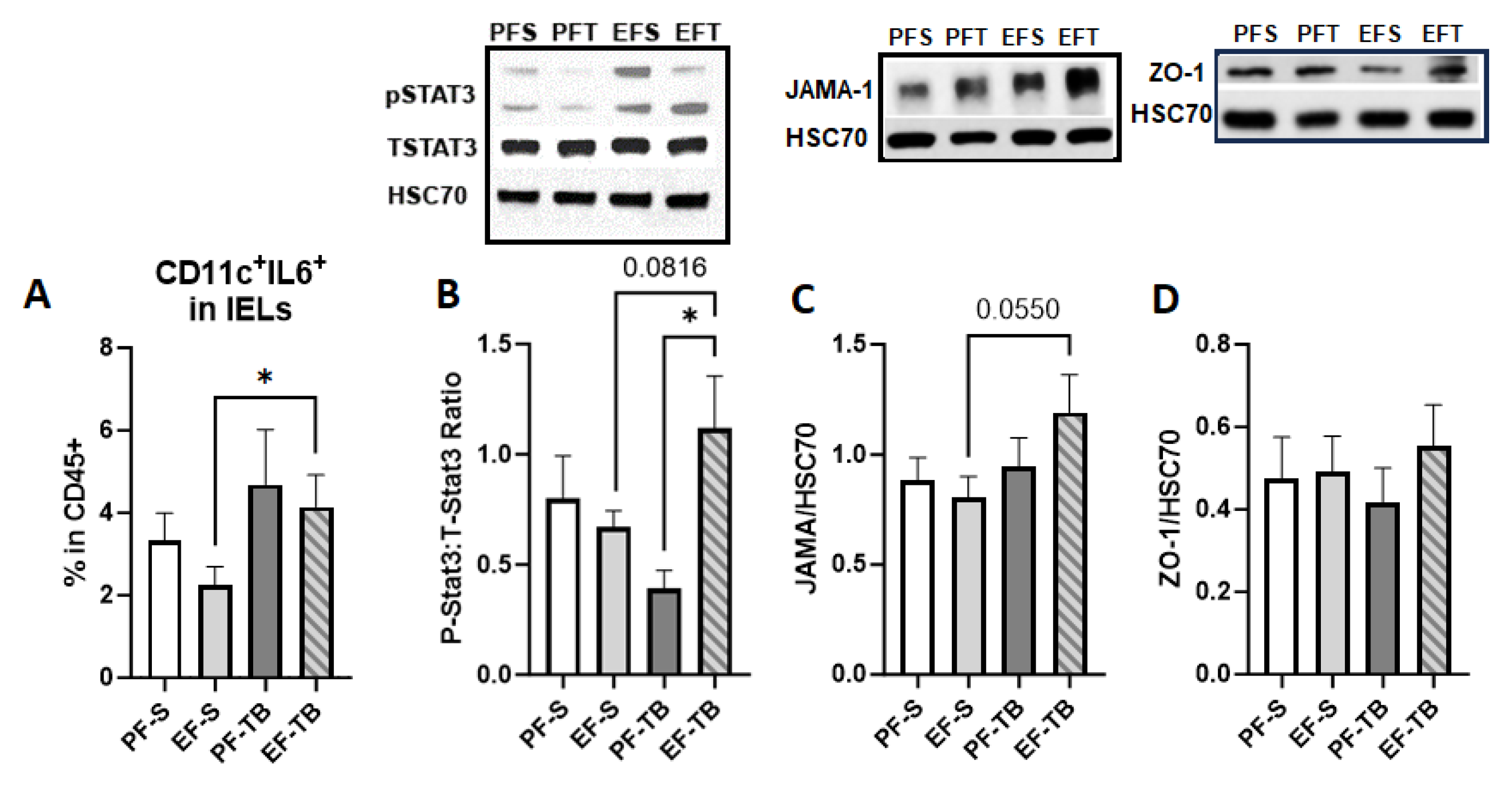
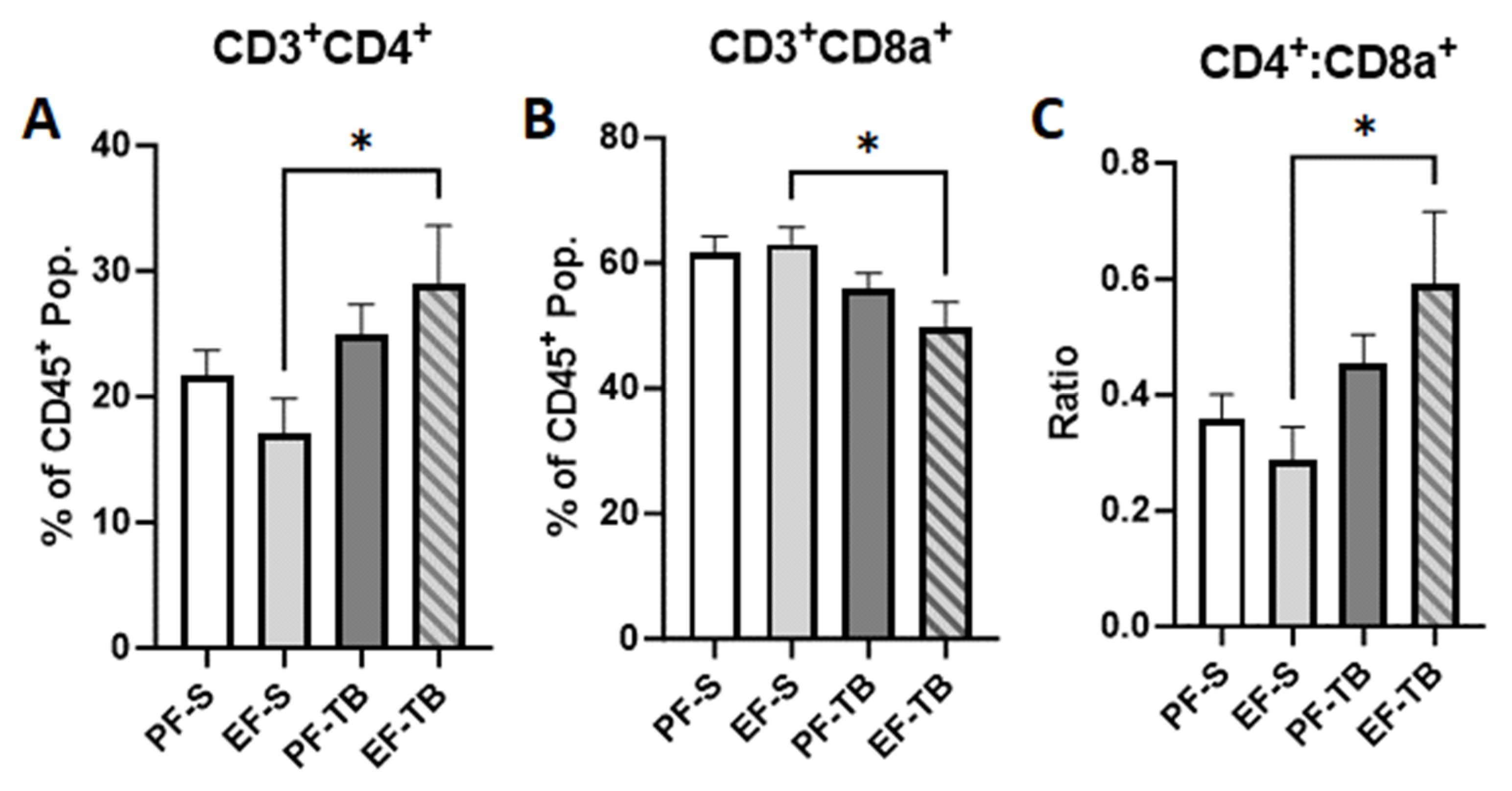
3.3. Tributyrin Supplementation Bolsters Immune and Barrier Functions Both at the Intestinal Epithelium and within the Lamina Propria
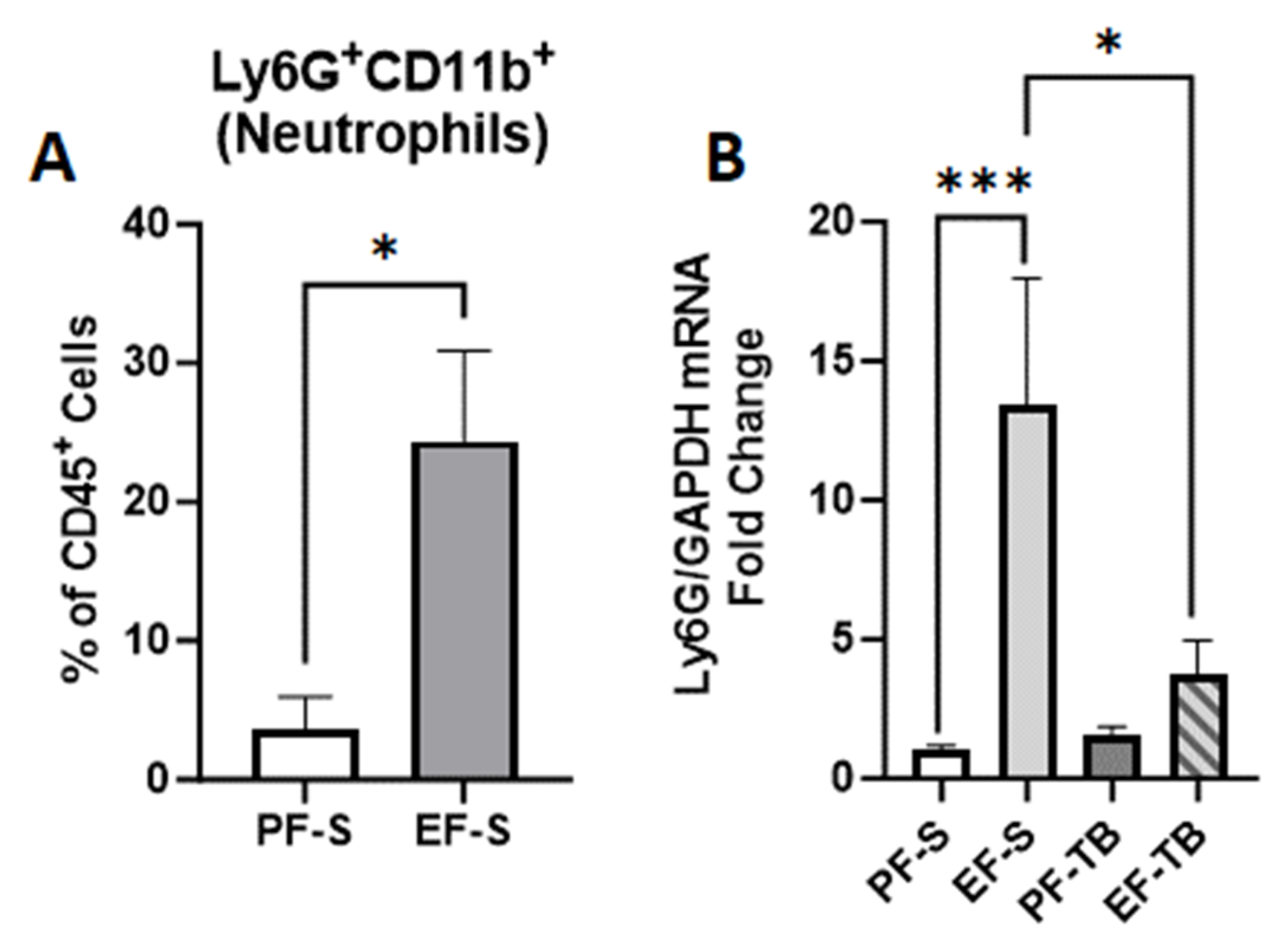
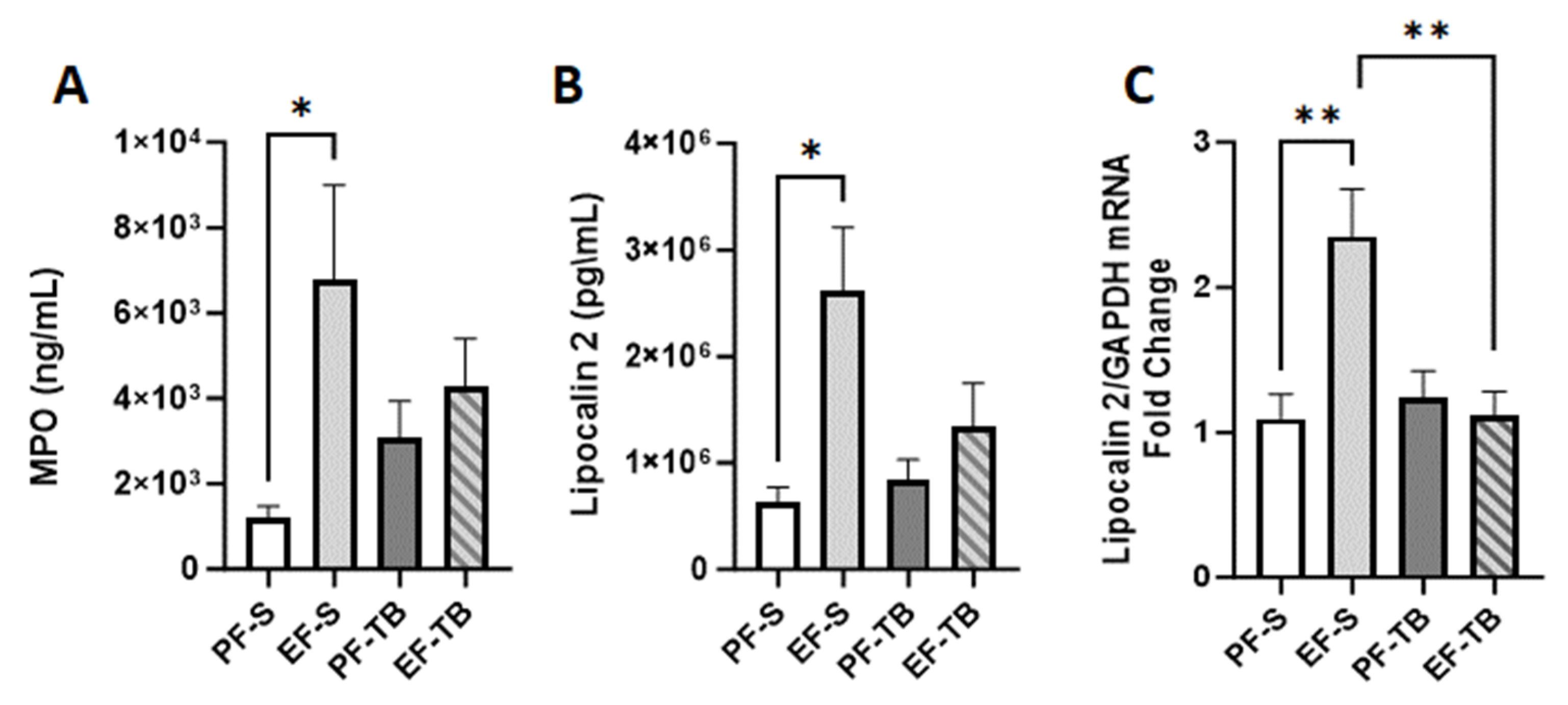
3.4. Tributyrin Supplementation Mitigates Ethanol-Induced Neutrophil Presence and Markers of Oxidative Stress in the Lungs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jew, M.H.; Hsu, C.L. Alcohol, the Gut Microbiome, and Liver Disease. J. Gastroenterol. Hepatol. 2023, 38, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An Insight into Gut Microbiota and Its Functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate a double edged sword for health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, Y.; Yang, G.; Zhang, Q.; Meng, L.; Xin, Y.; Jiang, X. The Role of Short-Chain Fatty Acids in Intestinal Barrier Function, Inflammation, Oxidative Stress, and Colonic Carcinogenesis. Pharmacol. Res. 2021, 165, 105420. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Puri, P.; Muthiah, M.D.; Daitya, K.; Brown, R.; Chalasani, N.; Liangpunsakul, S.; Shah, V.H.; Gelow, K.; Siddiqui, M.S.; et al. Fecal Microbiome Distinguishes Alcohol Consumption from Alcoholic Hepatitis but Does Not Discriminate Disease Severity. Hepatology 2020, 72, 271–286. [Google Scholar] [CrossRef]
- Cresci, G.A.; Bush, K.; Nagy, L.E. Tributyrin Supplementation Protects Mice from Acute Ethanol-Induced Gut Injury. Alcohol. Clin. Exp. Res. 2014, 38, 1489–1501. [Google Scholar] [CrossRef]
- Cresci, G.A.; Glueck, B.; McMullen, M.R.; Xin, W.; Allende, D.; Nagy, L.E. Prophylactic Tributyrin Treatment Mitigates Chronic-Binge Ethanol-Induced Intestinal Barrier and Liver Injury. J. Gastroenterol. Hepatol. 2017, 32, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Roychowdhury, S.; Glueck, B.; Han, Y.; Mohammad, M.A.; Cresci, G.A.M. A Designer Synbiotic Attenuates Chronic-Binge Ethanol-Induced Gut-Liver Injury in Mice. Nutrients 2019, 11, 97. [Google Scholar] [CrossRef]
- Han, Y.; Glueck, B.; Shapiro, D.; Miller, A.; Roychowdhury, S.; Cresci, G.A.M. Dietary Synbiotic Supplementation Protects Barrier Integrity of Hepatocytes and Liver Sinusoidal Endothelium in a Mouse Model of Chronic-Binge Ethanol Exposure. Nutrients 2020, 12, 373. [Google Scholar] [CrossRef]
- Sarkar, D.; Katherine Jung, M.; Joe Wang, H.; Sarkar, D.K. Alcohol and the Immune System. Alcohol Res. 2015, 37, 153–155. [Google Scholar]
- Simet, S.M.; Sisson, J.H. Alcohol’s Effects on Lung Health Immunity. Alcohol Res. 2015, 37, 199–208. [Google Scholar] [PubMed]
- Ma, Y.; Yang, X.; Chatterjee, V.; Wu, M.H.; Yuan, S.Y. The Gut–Lung Axis in Systemic Inflammation Role of Mesenteric Lymph as a Conduit. Am. J. Respir. Cell Mol. Biol. 2021, 64, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.B.; Farhadi, A.; Jakate, S.M.; Tang, Y.; Shaikh, M.; Keshavarzian, A. Lactobacillus GG Treatment Ameliorates Alcohol-Induced Intestinal Oxidative Stress, Gut Leakiness, and Liver Injury in a Rat Model of Alcoholic Steatohepatitis. Alcohol 2009, 43, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, N.; Zarraonaindia, I.; Hampton-Marcell, J.T.; Ssegane, H.; Eshoo, T.W.; Rijal, G.; Negri, M.C.; Gilbert, J.A. Differential Functional Constraints Cause Strain-Level Endemism in Polynucleobacter Populations. mSystems 2016, 1, 00003-16. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Waldron, L.; Ballarini, A.; Narasimhan, V.; Jousson, O.; Huttenhower, C. Metagenomic Microbial Community Profiling Using Unique Clade-Specific Marker Genes. Nat. Methods 2012, 9, 811–814. [Google Scholar] [CrossRef]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-Level Functional Profiling of Metagenomes and Metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y. Discovering the False Discovery Rate. J. R. Statist. Soc. B 2010, 72, 405–416. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 3 November 2023).
- Qiu, Z.; Sheridan, B.S. Isolating Lymphocytes from the Mouse Small Intestinal Immune System. J. Vis. Exp. 2018, 2018, e57281. [Google Scholar]
- Siddiqui, M.T.; Han, Y.; Shapiro, D.; West, G.; Fiocchi, C.; Cresci, G.A.M. The Postbiotic Butyrate Mitigates Gut Mucosal Disruption Caused by Acute Ethanol Exposure. Int. J. Mol. Sci. 2024, 25, 1665. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, D.; Kapourchali, F.R.; Santilli, A.; Han, Y.; Cresci, G.A.M. Targeting the Gut Microbiota and Host Immunity with a Bacilli-Species Probiotic during Antibiotic Exposure in Mice. Microorganisms 2022, 10, 1178. [Google Scholar] [CrossRef] [PubMed]
- Calleja-Conde, J.; Echeverry-Alzate, V.; Bühler, K.M.; Durán-González, P.; Morales-García, J.Á.; Segovia-Rodríguez, L.; Rodríguez de Fonseca, F.; Giné, E.; López-Moreno, J.A. The Immune System through the Lens of Alcohol Intake and Gut Microbiota. Int. J. Mol. Sci. 2021, 22, 7485. [Google Scholar] [CrossRef] [PubMed]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and Gut-Derived Inflammation. Alcohol Res. 2017, 38, 163–171. [Google Scholar]
- Gout, I. Coenzyme A: A Protective Thiol in Bacterial Antioxidant Defence. Biochem. Soc. Trans. 2019, 47, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ortiz, G.; Sidda, J.D.; Peate, J.; Ciccarelli, D.; Ding, Y.; Barry, S.M. Production of Copropophyrin III, Biliverdin and Bilirubin by the Rufomycin Producer, Streptomyces Atratus. Front. Microbiol. 2023, 14, 1092166. [Google Scholar] [CrossRef]
- Wilks, A.; Ikeda-Saito, M. Heme Utilization by Pathogenic Bacteria: Not All Pathways Lead to Biliverdin. Acc. Chem. Res. 2014, 47, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Layer, G.; Jahn, M.; Moser, J.; Jahn, D. Radical SAM Enzymes Involved in Tetrapyrrole Biosynthesis and Insertion. ACS Bio. Med. Chem. Au 2022, 2, 196–204. [Google Scholar] [CrossRef]
- Li, J.Y.; Guo, Y.C.; Zhou, H.F.; Yue, T.T.; Wang, F.X.; Sun, F.; Wang, W.Z. Arginine Metabolism Regulates the Pathogenesis of Inflammatory Bowel Disease. Nutr. Rev. 2023, 81, 578–586. [Google Scholar] [CrossRef]
- Nüse, B.; Holland, T.; Rauh, M.; Gerlach, R.G.; Mattner, J. L-Arginine Metabolism as Pivotal Interface of Mutual Host–Microbe Interactions in the Gut. Gut Microbes 2023, 15, 2222961. [Google Scholar] [CrossRef] [PubMed]
- López, M.C. Chronic Alcohol Consumption Regulates the Expression of Poly Immunoglobulin Receptor (PIgR) and Secretory IgA in the Gut. Toxicol. Appl. Pharmacol. 2017, 333, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s Complex Roles in Immunity and Mucosal Homeostasis in the Gut. Mucosal. Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory Iga in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int. J. Mol. Sci. 2020, 21, 9254. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, B.; Wang, T.; Gao, L.; Yang, Z.J.; Wang, F.F.; Shang, H.W.; Hua, R.; Xu, J.D. Biological Characteristics of Il-6 and Related Intestinal Diseases. Int. J. Biol. Sci. 2020, 17, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Chen, W.D.; Wang, Y.D. Gut Microbiota: An Integral Moderator in Health and Disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal 2020, 32, 957–981. [Google Scholar] [CrossRef]
- An, H.S.; Yoo, J.W.; Jeong, J.H.; Heo, M.; Hwang, S.H.; Jang, H.M.; Jeong, E.A.; Lee, J.; Shin, H.J.; Kim, K.E.; et al. Lipocalin-2 Promotes Acute Lung Inflammation and Oxidative Stress by Enhancing Macrophage Iron Accumulation. Int. J. Biol. Sci. 2023, 19, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Jeong, E.A.; Lee, J.Y.; An, H.S.; Jang, H.M.; Ahn, Y.J.; Lee, J.; Kim, K.E.; Roh, G.S. Lipocalin-2 Deficiency Reduces Oxidative Stress and Neuroinflammation and Results in Attenuation of Kainic Acid-Induced Hippocampal Cell Death. Antioxidants 2021, 10, 100. [Google Scholar] [CrossRef]
- Xiao, X.; Yeoh, B.S.; Vijay-Kumar, M. Lipocalin 2: An Emerging Player in Iron Homeostasis and Inflammation. Annu. Rev. Nutr. 2017, 37, 103–130. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, E.; Liu, C.; Wicks, H.; Yildiz, S.; Razack, F.; Ying, Z.; Kooijman, S.; Koonen, D.P.Y.; Heijink, M.; et al. Dietary Butyrate Ameliorates Metabolic Health Associated with Selective Proliferation of Gut Lachnospiraceae Bacterium 28-4. JCI Insight 2023, 8, e166655. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Chiang, H.C.; Gordon, J.I. Mucosal Glycan Foraging Enhances Fitness and Transmission of a Saccharolytic Human Gut Bacterial Symbiont. Cell Host Microbe 2008, 4, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Blaser, M.J. Probiotic Compositions for Improving Metabolism and Immunity. US Patent 10,653,728 B2, 19 May 2020. [Google Scholar]
- Bang, S.; Shin, Y.H.; Ma, X.; Park, S.M.; Graham, D.B.; Xavier, R.J.; Clardy, J. A Cardiolipin from Muribaculum Intestinale Induces Antigen-Specific Cytokine Responses. J. Am. Chem. Soc. 2023, 145, 23422–23426. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, N.; Moser, J.; Jahn, D. Bacterial Heme Biosynthesis and Its Biotechnological Application. Appl. Microbiol. Biotechnol. 2003, 63, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Pascale, R.M.; Simile, M.M.; Calvisi, D.F.; Feo, C.F.; Feo, F. S-Adenosylmethionine: From the Discovery of Its Inhibition of Tumorigenesis to Its Use as a Therapeutic Agent. Cells 2022, 11, 409. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cui, J.; Fang, C.; Liu, M.; Min, G.; Li, L. S-Adenosylmethionine Attenuates Oxidative Stress and Neuroinflammation Induced by Amyloid-β Through Modulation of Glutathione Metabolism. J. Alzheimer’s Dis. 2017, 58, 549–558. [Google Scholar] [CrossRef]
- Shandilya, S.; Kumar, S.; Kumar Jha, N.; Kumar Kesari, K.; Ruokolainen, J. Interplay of Gut Microbiota and Oxidative Stress: Perspective on Neurodegeneration and Neuroprotection. J. Adv. Res. 2022, 38, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Kunst, C.; Schmid, S.; Michalski, M.; Tümen, D.; Buttenschön, J.; Müller, M.; Gülow, K. The Influence of Gut Microbiota on Oxidative Stress and the Immune System. Biomedicines 2023, 11, 1388. [Google Scholar] [CrossRef] [PubMed]
- Bastos, P.A.D.; Wheeler, R.; Boneca, I.G. Uptake, Recognition and Responses to Peptidoglycan in the Mammalian Host. FEMS Microbiol. Rev. 2021, 45, fuaa044. [Google Scholar] [CrossRef]
- Wolf, A.J.; Underhill, D.M. Peptidoglycan Recognition by the Innate Immune System. Nat. Rev. Immunol. 2018, 18, 243–254. [Google Scholar] [CrossRef]
- Tezuka, H.; Ohteki, T. Regulation of IgA Production by Intestinal Dendritic Cells and Related Cells. Front. Immunol. 2019, 10, 1891. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Yuan, X.; Cao, Q.; Chen, Y.; Peng, X.; Lin, J.; Li, Y.; Ma, W.; Gao, S.; Chen, D. Gut Microbiota Regulate Migration of Lymphocytes from Gut to Lung. Microb. Pathog. 2023, 183, 106311. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.G.; Beier, J.I.; Torres-Gonzales, E.; Schlueter, C.F.; Hudson, S.V.; Artis, A.; Warner, N.L.; Nguyen-Ho, C.T.; Dolin, C.E.; Ritzenthaler, J.D.; et al. Chronic + Binge Alcohol Exposure Promotes Inflammation and Alters Airway Mechanics in the Lung. Alcohol 2019, 80, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Role of Myeloperoxidase and Oxidant Formation in the Extracellular Environment in Inflammation-Induced Tissue Damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Sequence |
|---|---|---|
| Lymphocyte Antigen 6 Complex Locus G6D (Ly6G) NM_001310438 | Ly6G F | TGC GTT GCT CTG GAG ATA GA |
| Ly6G R | CAG AGT AGT GGG GCA GAT GG | |
| Lipocalin 2 NM-008491 | LCN2 F | TGG CCC TGA GTG TCA TGT G |
| LCN2 R | CTC TTG TAG CTC ATA GAT GGT GC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santilli, A.; Shapiro, D.; Han, Y.; Sangwan, N.; Cresci, G.A.M. Tributyrin Supplementation Rescues Chronic–Binge Ethanol-Induced Oxidative Stress in the Gut–Lung Axis in Mice. Antioxidants 2024, 13, 472. https://doi.org/10.3390/antiox13040472
Santilli A, Shapiro D, Han Y, Sangwan N, Cresci GAM. Tributyrin Supplementation Rescues Chronic–Binge Ethanol-Induced Oxidative Stress in the Gut–Lung Axis in Mice. Antioxidants. 2024; 13(4):472. https://doi.org/10.3390/antiox13040472
Chicago/Turabian StyleSantilli, Anthony, David Shapiro, Yingchun Han, Naseer Sangwan, and Gail A. M. Cresci. 2024. "Tributyrin Supplementation Rescues Chronic–Binge Ethanol-Induced Oxidative Stress in the Gut–Lung Axis in Mice" Antioxidants 13, no. 4: 472. https://doi.org/10.3390/antiox13040472
APA StyleSantilli, A., Shapiro, D., Han, Y., Sangwan, N., & Cresci, G. A. M. (2024). Tributyrin Supplementation Rescues Chronic–Binge Ethanol-Induced Oxidative Stress in the Gut–Lung Axis in Mice. Antioxidants, 13(4), 472. https://doi.org/10.3390/antiox13040472