
Effects of Dietary Inosine 5′-Monophosphate Supplementation on the Growth Performance and Salinity and Oxidative Stress Resistance of Gibel Carp (Carassius auratus gibelio)

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Experimental Diets
2.3. Fish and Rearing Conditions
2.4. Sampling
2.5. Salinity Stress Trial
2.6. Chemical and Physiological Analysis
2.7. Real-Time PCR Analysis
2.8. Statistical Analysis
3. Results
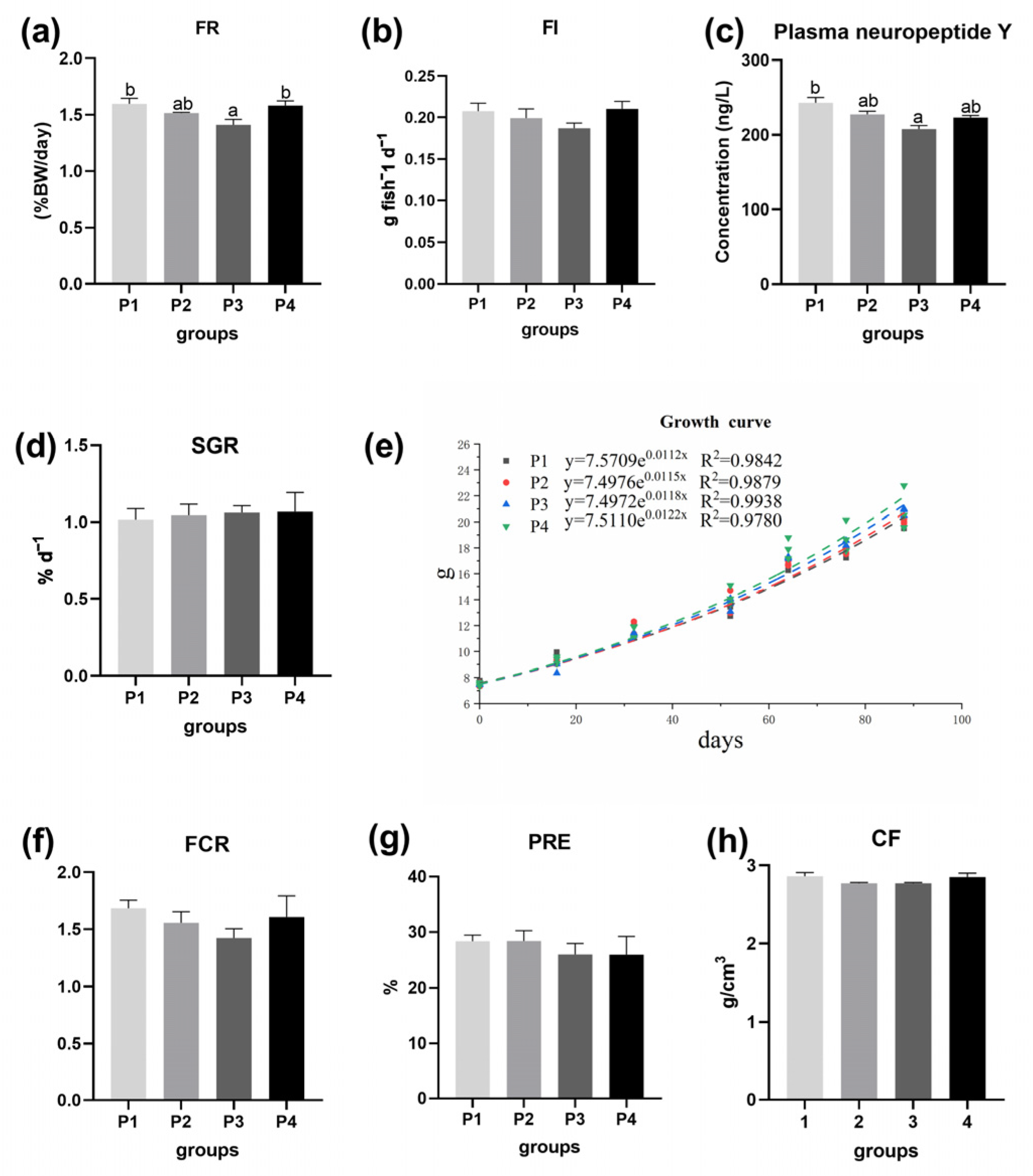
3.1. Growth Performance, Morphology and Body Chemical Composition
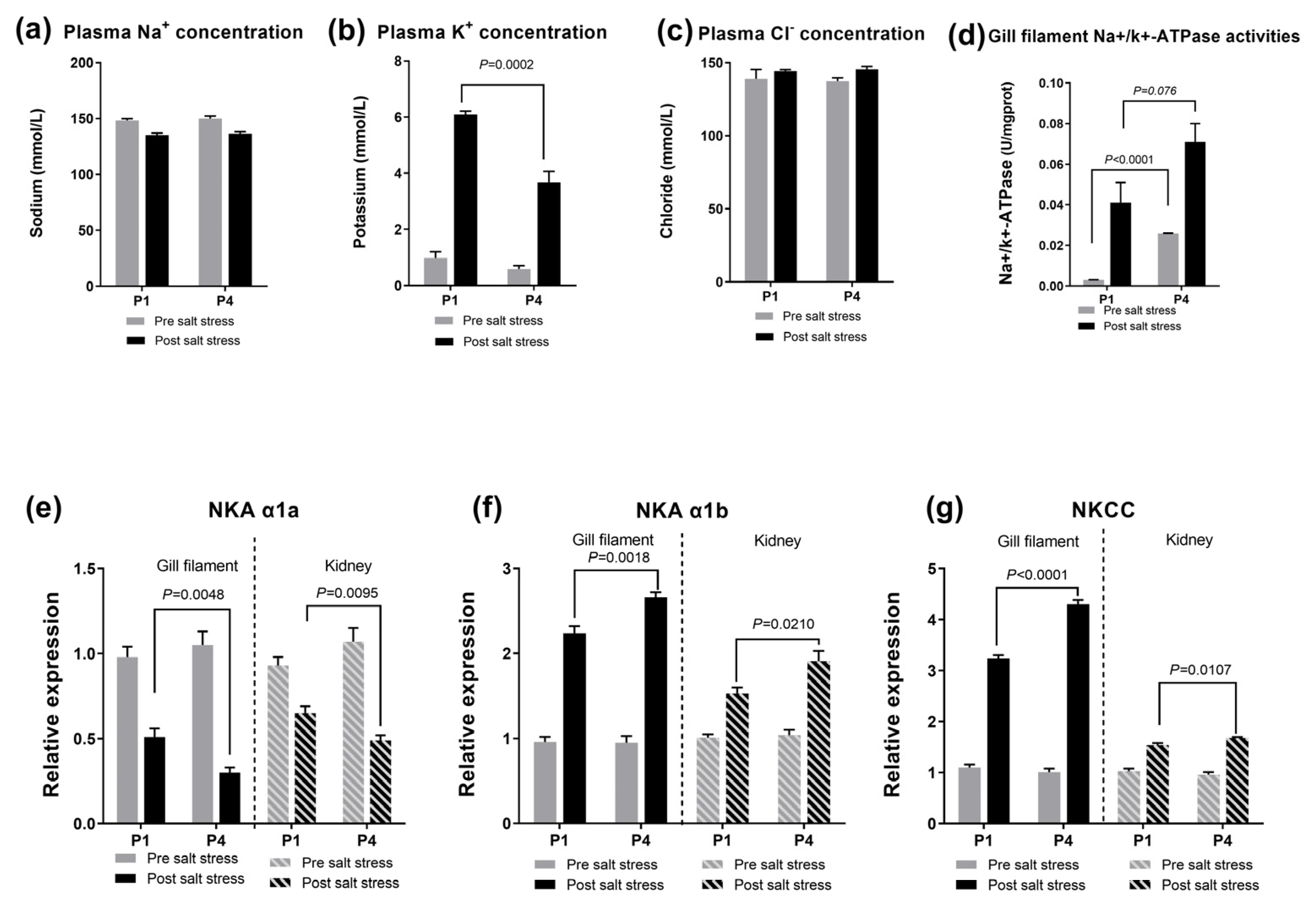
3.2. The Reaction to Salinity Stress and Related Gene Expression in the Gill Filament and in the Kidney
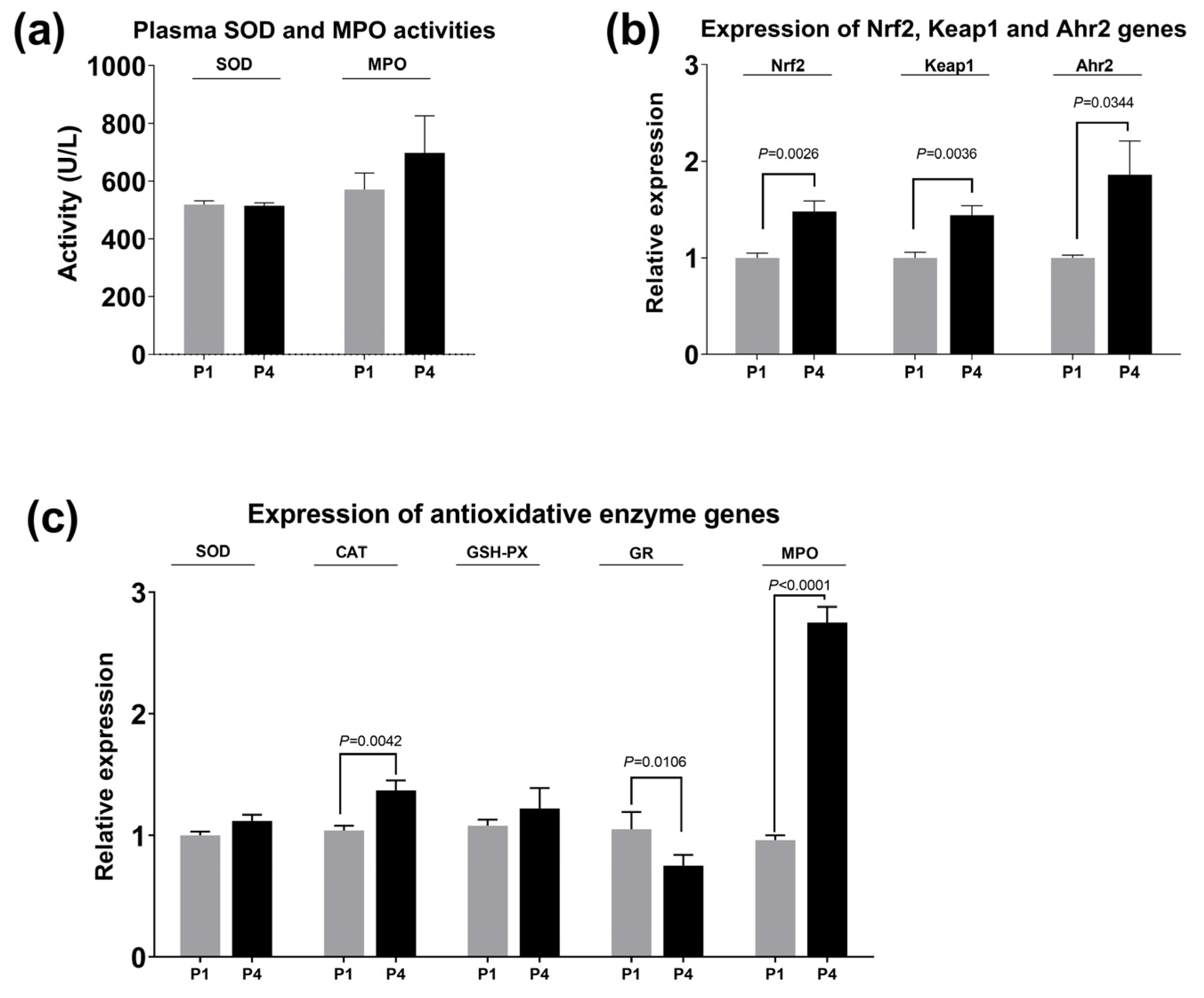
3.3. Activity of Plasma Antioxidative Enzymes and Hepatic-Related Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO Statistics Division. 2018 FAO Yearbook. Fishery and Aquaculture Statistics; FAO: Rome, Italy, 2018. [Google Scholar]
- Fisheries Administration, Chinese Ministry of Agriculture. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2017; pp. 32–35.
- Citarasu, T.; Babu, M.M.; Sekar, R.R.J.; Petermarian, M. Developing Artemia enriched herbal diet for producing quality larvae in Penaeus monodon, Fabricius. Asian Fish. Sci. 2002, 15, 21–32. [Google Scholar] [CrossRef]
- Li, Q.; Xi, M.; Wang, Q.; Kong, F.; Li, Y. Characterization of soil salinization in typical estuarine area of the Jiaozhou Bay, China. Phys. Chem. Earth Parts A/B/C 2018, 103, 51–61. [Google Scholar] [CrossRef]
- Islam, M.; Yasmin, R. Impact of Aquaculture and Contemporary environmental issues in Bangladesh. Int. J. Fish. Aquat. Stud. 2017, 5, 100–107. [Google Scholar]
- Iglesias, M.C.A. A review of recent advances and future challenges in freshwater salinization. Limnetica 2020, 39, 185–211. [Google Scholar]
- Hosios, A.M.; Vander Heiden, M.G. The redox requirements of proliferating mammalian cells. J. Biol. Chem. 2018, 293, 7490–7498. [Google Scholar] [CrossRef] [PubMed]
- Grimble, G.K.; Westwood, O.M. Nucleotides. In Nutrition and Immunology; Humana Press: Totowa, NJ, USA, 2000; pp. 135–144. [Google Scholar]
- Gil, A. Modulation of the immune response mediated by dietary nucleotides. Eur. J. Clin. Nutr. 2002, 56, S1. [Google Scholar] [CrossRef] [PubMed]
- Mackie, A.M. The chemical basis of food detection in the lobster Homarus gammarus. Mar. Biol. 1973, 21, 103–108. [Google Scholar] [CrossRef]
- Mackie, A.M.; Adron, J.W. Identification of inosine and inosine 5′-monophosphate as the gustatory feeding stimulants for the turbot, Scophthalmus maximus. Comp. Biochem. Physiol. Part A Physiol. 1978, 60, 79–83. [Google Scholar] [CrossRef]
- Kubitza, F.; Lovshin, L.L.; Lovell, R.T. Identification of feed enhancers for juvenile largemouth bass Micropterus salmoides. Aquaculture 1997, 148, 191–200. [Google Scholar] [CrossRef]
- Li, P.; Gatlin, D.M., III. Nucleotide nutrition in fish: Current knowledge and future applications. Aquaculture 2006, 251, 141–152. [Google Scholar] [CrossRef]
- Burrells, C.; Williams, P.D.; Southgate, P.J.; Wadsworth, S.L. Dietary nucleotides: A novel supplement in fish feeds: 2. Effects on vaccination, salt water transfer, growth rates and physiology of Atlantic salmon (Salmo salar L.). Aquaculture 2001, 199, 171–184. [Google Scholar] [CrossRef]
- Sakai, M.; Taniguchi, K.; Mamoto, K.; Ogawa, H.; Tabata, M. Immunostimulant effects of nucleotide isolated from yeast RNA on carp, Cyprinus carpio L. J. Fish Dis. 2001, 24, 433–438. [Google Scholar] [CrossRef]
- Li, P.; Lewis, D.H.; Gatlin, D.M., III. Dietary oligonucleotides from yeast RNA influence immune responses and resistance of hybrid striped bass (Morone chrysops × Morone saxatilis) to Streptococcus iniae infection. Fish Shellfish. Immunol. 2004, 16, 561–569. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wang, H.; Shiau, S.Y. Dietary nucleotide supplementation enhances growth and immune responses of grouper, Epinephelus malabaricus. Aquac. Nutr. 2009, 15, 117–122. [Google Scholar] [CrossRef]
- Cheng, Z.; Buentello, A.; Gatlin, D.M., III. Dietary nucleotides influence immune responses and intestinal morphology of red drum Sciaenops ocellatus. Fish Shellfish. Immunol. 2011, 30, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi-Kohyani, A.; Keyvanshokooh, S.; Nematollahi, A.; Mahmoudi, N.; Pasha-Zanoosi, H. Effects of dietary nucleotides supplementation on rainbow trout (Oncorhynchus mykiss) performance and acute stress response. Fish Physiol. Biochem. 2012, 38, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Song, J.W.; Lim, S.J.; Lee, K.J. Effects of dietary supplementation of inosine monophosphate on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus). Fish Shellfish. Immunol. 2012, 33, 1050–1054. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Ono, S.; Fujieda, T. Comparison of the effects of inosine and inosine monophosphate on growth, immune response, stress resistance and gut morphology of juvenile red sea bream, Pagrus major. Aquaculture 2016, 458, 64–74. [Google Scholar] [CrossRef]
- Hossain, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.; Usami, M.; Ono, S.; Fujieda, T. Inosine supplementation effectively provokes the growth, immune response, oxidative stress resistance and intestinal morphology of juvenile red sea bream, Pagrus major. Aquac. Nutr. 2017, 23, 952–963. [Google Scholar] [CrossRef]
- Mateo, C.D.; Peters, D.N.; Dave, R.I.; Rosa, A.; Stein, H.H. Effects of dietary nucleotides on intestinal morphology and microbial activity in newly weaned pigs. J. Anim. Sci. 2005, 83, 69. [Google Scholar]
- Stein, H.H.; Kil, D.Y. Reduced use of antibiotic growth promoters in diets fed to weanling pigs: Dietary tools, part 2. Anim. Biotechnol. 2006, 17, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Asaduzzaman, M.; Ikeda, D.; Abol-Munafi, A.B.; Bulbul, M.; Ali, E.; Kinoshita, S.; Watabe, S.; Kader, A. Dietary supplementation of inosine monophosphate promotes cellular growth of muscle and upregulates growth-related gene expression in Nile tilapia Oreochromis niloticus. Aquaculture 2017, 468, 297–306. [Google Scholar] [CrossRef]
- Zhang, P.; Fu, L.; Liu, H.; Huda, N.U.; Zhu, X.; Han, D.; Jin, J.; Yang, Y.; Kim, Y.; Xie, S. Effects of inosine 5′-monophosphate supplementation in high fishmeal and high soybean diets on growth, immune-related gene expression in gibel carp (Carassius auratus gibelio var. CAS III), and its challenge against Aeromonas hydrophila infection. Fish Shellfish. Immunol. 2019, 86, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S. Increased salinization decreases safe drinking water. Environ. Sci. Technol. 2016, 50, 2765–2766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Z.; Dong, S. Advance in research on the effects of pH, salinity and alkalinity on freshwater cultured species. J. Fish. Sci. China 1999, 6, 95–98. [Google Scholar]
- Huang, K.; Wang, W.; Lu, J.; Dai, X.W.; Zhou, J.N. Salinity effects on growth and biochemical composition of Penaeus vannamei. Mar. Sci. Qingdao-Chin. Ed. 2004, 28, 20–25. [Google Scholar]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2003. [Google Scholar]
- Speckner, W.; Schindler, J.F.; Albers, C. Age-dependent changes in volume and haemoglobin content of erythrocytes in the carp (Cyprinus carpio L.). J. Exp. Biol. 1989, 141, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jin, J.; Zhu, X.; Han, D.; Yang, Y.; Xie, S. Effect of substitution of dietary fish meal by soybean meal on different sizes of gibel carp (Carassius auratus gibelio): Digestive enzyme gene expressions and activities, and intestinal and hepatic histology. Aquac. Nutr. 2017, 23, 129–147. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef]
- Hoskins, L.J.; Volkoff, H. Daily patterns of mRNA expression of two core circadian regulatory proteins, Clock2 and Per1, and two appetite-regulating peptides, OX and NPY, in goldfish (Carassius auratus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163, 127–136. [Google Scholar] [CrossRef]
- Volkoff, H.; Wyatt, J.L. Apelin in goldfish (Carassius auratus): Cloning, distribution and role in appetite regulation. Peptides 2009, 30, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerda-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Chang, Q. Identification of feeding stimulants for shrimp (Penaeus chinensis). Mar. Fish. Res. 2001, 22, 71–74. [Google Scholar]
- Liang, M.Q.; Hong, Y.U.; Chang, Q.; Chen, C.; Sun, S.G. Feeding attraction activities of food attractants for 3 species of fishes. J. Fish. China 2000, 7, 60–63. [Google Scholar]
- Ishida, Y.; Hidaka, I. Gustatory response profiles for amino acids, glycinebetaine, and nucleotides in several marine teleosts. Nippon. Suisan Gakkaishi 1987, 53, 1391–1398. [Google Scholar] [CrossRef]
- Masic, U.; Yeomans, M.R. Umami flavor enhances appetite but also increases satiety. Am. J. Clin. Nutr. 2014, 100, 532–538. [Google Scholar] [CrossRef]
- Weaver, A.C.; Kim, S.W. Supplemental nucleotides high in inosine 5′-monophosphate to improve the growth and health of nursery pigs. J. Anim. Sci. 2014, 92, 645–651. [Google Scholar] [CrossRef]
- Tavares-Dias, M. Toxicity, physiological, histopathological, handling, growth and antiparasitic effects of the sodium chloride (salt) in the freshwater fish aquaculture. Aquac. Res. 2022, 53, 715–734. [Google Scholar] [CrossRef]
- Alderdice, D.F. 3 Osmotic and Ionic Regulation in Teleost Eggs and Larvae. In Fish Physiology; Academic Press: Cambridge, MA, USA, 1988; Volume 11, pp. 163–251. [Google Scholar]
- Hegab, S.A.; Hanke, W. Electrolyte changes and volume regulatory processes in the carp (Cyprinus carpio) during osmotic stress. Comp. Biochem. Physiol. Part A Physiol. 1982, 71, 157–164. [Google Scholar] [CrossRef]
- Gerber, L.; Madsen, S.S.; Jensen, F.B. Cortisol regulates nitric oxide synthase in freshwater and seawater acclimated rainbow trout, Oncorhynchus mykiss. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 204, 1–8. [Google Scholar] [CrossRef]
- Basu, N.; Kennedy, C.J.; Iwama, G.K. The effects of stress on the association between hsp70 and the glucocorticoid receptor in rainbow trout. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 134, 655–663. [Google Scholar] [CrossRef]
- Tipsmark, C.K.; Madsen, S.S.; Seidelin, M.; Christensen, A.S.; Cutler, C.P.; Cramb, G. Dynamics of Na+, K+, 2Cl− cotransporter and Na+, K+-ATPase expression in the branchial epithelium of brown trout (Salmo trutta) and atlantic salmon (Salmo salar). J. Exp. Zool. 2002, 293, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.G.; Semple, J.W.; Bystriansky, J.S.; Schulte, P.M. Na+/K+-ATPase α-isoform switching in gills of rainbow trout (Oncorhynchus mykiss) during salinity transfer. J. Exp. Biol. 2003, 206, 4475–4486. [Google Scholar] [CrossRef]
- Nilsen, T.O.; Ebbesson, L.O.; Madsen, S.S.; McCormick, S.D.; Andersson, E.; Björnsson, B.T.; Prunet, P.; Stefansson, S.O. Differential expression of gill Na+, K+-ATPaseα-and β-subunits, Na+, K+, 2Cl−-cotransporter and CFTR anion channel in juvenile anadromous and landlocked Atlantic salmon Salmo salar. J. Exp. Biol. 2007, 210, 2885–2896. [Google Scholar] [CrossRef] [PubMed]
- Ip, Y.K.; Wilson, J.M.; Loong, A.M.; Chen, X.L.; Wong, W.P.; Delgado, I.L.S.; Lam, S.H.; Chew, S.F. Cystic fibrosis transmembrane conductance regulator in the gills of the climbing perch, Anabas testudineus, is involved in both hypoosmotic regulation during seawater acclimation and active ammonia excretion during ammonia exposure. J. Comp. Physiol. B 2012, 182, 793–812. [Google Scholar] [CrossRef]
- Urbina, M.A.; Schulte, P.M.; Bystriansky, J.S.; Glover, C.N. Differential expression of Na+, K+-ATPase α-1 isoforms during seawater acclimation in the amphidromous galaxiid fish Galaxias maculatus. J. Comp. Physiol. B 2013, 183, 345–357. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D.; Regish, A.; O’Dea, M.F.; Shrimpton, J.M. Are we missing a mineralocorticoid in teleost fish? Effects of cortisol, deoxycorticosterone and aldosterone on osmoregulation, gill Na+, K+-ATPase activity and isoform mRNA levels in Atlantic salmon. Gen. Comp. Endocrinol. 2008, 157, 35–40. [Google Scholar] [CrossRef]
- Leray, C.; Colin, D.A.; Florentz, A. Time course of osmotic adaptation and gill energetics of rainbow trout (Salmo gairdneri R.) following abrupt changes in external salinity. J. Comp. Physiol. 1981, 144, 175–181. [Google Scholar] [CrossRef]
- Atkinson, D.E. Cellular Energy Metabolism and Its Regulation; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Caldwell, C.A.; Hinshaw, J.M. Nucleotides and the adenylate energy charge as indicators of stress in rainbow trout (Oncorhynchus mykiss) subjected to a range of dissolved oxygen concentrations. Comp. Biochem. Physiol. Part B Comp. Biochem. 1994, 109, 313–323. [Google Scholar] [CrossRef]
- Sigholt, T.; Erikson, U.; Rustad, T.; Johansen, S.; Nordtvedt, T.S.; Seland, A. Handling stress and storage temperature affect meat quality of farmed-raised Atlantic salmon (Salmo salar). J. Food Sci. 1997, 62, 898–905. [Google Scholar] [CrossRef]
- Leonardi, M.; Sandino, A.M.; Klempau, A. Effect of a nucleotide-enriched diet on the immune system, plasma cortisol levels and resistance to infectious pancreatic necrosis (IPN) in juvenile rainbow trout (Oncorhynchus mykiss). Bull. Eur. Assoc. Fish Pathol. 2003, 23, 52–59. [Google Scholar]
- Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell Biol. 2003, 15, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, A.; Luchetti, E.; Marchetti, V.; Cardini, G.; Iorio, E.L. Analytical performances of d-ROMs test and BAP test in canine plasma. Definition of the normal range in healthy Labrador dogs. Vet. Res. Commun. 2008, 32, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ambrosone, C.B.; Ahn, J.; Singh, K.K.; Rezaishiraz, H.; Furberg, H.; Sweeney, C.; Coles, B.; Trovato, A. Polymorphisms in genes related to oxidative stress (MPO, MnSOD, CAT) and survival after treatment for breast cancer. Cancer Res. 2005, 65, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Dalton, T.P.; Puga, A.; Shertzer, H.G. Induction of cellular oxidative stress by aryl hydrocarbon receptor activation. Chem.-Biol. Interact. 2002, 141, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Indumathi, M.C.; Swetha, K.; Abhilasha, K.V.; Siddappa, S.; Kumar, S.M.; Prasad, G.K.; Chen, C.H.; Marathe, G.K. Selenium Ameliorates Acetaminophen-Induced Oxidative Stress via MAPK and Nrf2 Pathways in Mice. Biol. Trace Elem. Res. 2023, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Takeuchi, T.; Yokoyama, M.; Murata, Y. Effect of dietary supplementation with taurine, β-alanine and GABA on the growth of juvenile and fingerling Japanese flounder Paralichthys olivaceus. Fish. Sci. 2003, 69, 242–248. [Google Scholar] [CrossRef]
- Li, X.; Mu, W.; Wu, X.; Dong, Y.; Zhou, Z.; Wang, X.; Ma, L.; Ye, B.; Geng, L. The optimum methionine requirement in diets of juvenile hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂): Effects on survival, growth performance, gut micromorphology and immunity. Aquaculture 2020, 520, 735014. [Google Scholar] [CrossRef]
- Garg, C.K.; Sardar, P.; Sahu, N.P.; Maiti, M.K.; Shamna, N.; Varghese, T.; Deo, A.D.; Harikrishna, V. Dietary lysine requirement of genetically improved farmed tilapia (GIFT) juvenile reared in inland saline water of 10 ppt salinity. Aquaculture 2022, 555, 738223. [Google Scholar] [CrossRef]
- Liang, H.; Mokrani, A.; Ji, K.; Ge, X.; Ren, M.; Xie, J.; Liu, B.; Xi, B.; Zhou, Q. Dietary leucine modulates growth performance, Nrf2 antioxidant signaling pathway and immune response of juvenile blunt snout bream (Megalobrama amblycephala). Fish Shellfish. Immunol. 2018, 73, 57–65. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | P1 | P2 | P3 | P4 |
|---|---|---|---|---|
| Casein | 430.0 | 430.0 | 430.0 | 430.0 |
| Corn starch | 295.0 | 295.0 | 295.0 | 295.0 |
| Soy oil | 68.0 | 68.0 | 68.0 | 68.0 |
| Mineral premix | 50.0 | 50.0 | 50.0 | 50.0 |
| Vitamin premix | 3.9 | 3.9 | 3.9 | 3.9 |
| Choline chloride | 1.1 | 1.1 | 1.1 | 1.1 |
| Carboxymethyl cellulose | 30.0 | 30.0 | 30.0 | 30.0 |
| Monocalcium phosphate | 20.0 | 20.0 | 20.0 | 20.0 |
| 5′-IMP | 0.0 | 1.0 | 2.0 | 4.0 |
| Alanine | 4.1 | 3.1 | 2.1 | 0.0 |
| Cellulose | 97.9 | 97.9 | 97.9 | 98.0 |
| Approximate composition | ||||
| Crude protein | 385.8 | 396.0 | 389.3 | 393.6 |
| Crude lipid | 62.5 | 55.4 | 53.4 | 57.1 |
| Moisture | 116.6 | 116.6 | 124.4 | 114.2 |
| Crude ash | 66.5 | 65.7 | 66.1 | 66.9 |
| Gross energy (MJ/kg) | 203.1 | 202.1 | 203.6 | 203.2 |
| 5′-IMP | 0.1 | 0.6 | 1.3 | 2.2 |
| Gene | Acronym | Primer Sequence | Amplicon Size (bp) | Annealing Temp. (°C) | Accession No. |
|---|---|---|---|---|---|
| β-actin | β-actin | F: TTGAGCAGGAGATGGGAACCG | 115 | 60.0 | AB039726.2 |
| R: AGAGCCTCAGGGCAACGGAAA | |||||
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH | F: ATCCAACCAGATGGGAGAACG | 103 | 60.0 | MH820410 |
| R: ACCGTGTATGTGACCTGATGG | |||||
| Glucocorticoid receptor 1 | GR1 | F: AAGAAGAAACTGATGCGATTAC | 253 | 57.0 | HQ656017.1 |
| R: ACTTGACGGCAGAAACGAC | |||||
| Heat-shock protein 70 | Hsp70 | F: CTCAACAAGAGCATCAACCCAG | 155 | 60.0 | JN006055.1 |
| R: ATGACTCCACCAGCCGTTTC | |||||
| Sodium/potassium-transporting ATPase subunit alpha-1a | NKA-α1a | F: CTCTACCAACTGTGTTGAAGGTAC | 331 | 60.0 | MG759381 |
| R: GTTCTTCTTTGCCATTCGTTTGG | |||||
| Sodium/potassium-transporting ATPase subunit alpha-1b | NKA-α1b | F: TTCATCCACATCATCACCGG | 199 | 59.0 | MG759383 |
| R: GGCAGTTCTTCTTGGCCAT | |||||
| Solute carrier family 12 Member a1 | Slc12a1/NKCC | F: TGGTGGCTGTTTGATGACGGA | 257 | 57.0 | MG759385 |
| R: CGGAACGGCTCAATCATGTCCT | |||||
| Nuclear factor erythroid 2-related factor | Nrf2 | F: CCCTTCACCAAAGACAAGCA | 128 | 60.0 | MG759384 |
| R: TTGAAGTCATCCACAGGCAG | |||||
| Kelch-like-ECH-associated Protein 1 | Keap1 | F: CTCACCCCCAACTTCCTGCAG | 150 | 58.0 | MG759382 |
| R: GATGAGCTGCGGCACCTTGGG | |||||
| Aryl hydrocarbon receptor 2 | Ahr2 | F: TACAAACGGACAAGGATTC | 111 | 57.0 | FJ554572.1 |
| R: CTGTTGGTGGTCAGATGAG | |||||
| Superoxide dismutase | SOD | F: GTCCGCACTACAACCCTCAT | 134 | 59.0 | JQ776518.1 |
| R: GGTCACCATTTTATCCACAA | |||||
| Catalase | CAT | F: CTCCAACGGCAACTTCCCAT | 102 | 57.0 | JX477239.1 |
| R: CACACCTTAGTCAAATCAAA | |||||
| Glutathione peroxidase | GSH-Px | F: GCCCACCCTCTGTTTGTGTT | 244 | 58.0 | DQ983598.1 |
| R: AGGTTTATTTCGCCCTCTTC | |||||
| Glutathione reductase | GR | F: AGGAGGGGTACTGAGGAA | 140 | 59.0 | EF139092.1 |
| R: CCAGCGTAACAGAAAGGC | |||||
| Myeloperoxidase | MPO | F: AATGATGGAGGTCTTCTACG | 112 | 57.0 | KF417504.1 |
| R: TAAGGATTTTCTGACGTGTG |
| Moisture | Crude Protein | Crude Lipid | Ash | |
|---|---|---|---|---|
| P1 | 69.65 ± 0.14 | 17.28 ± 0.12 | 7.06 ± 0.13 | 4.86 ± 0.04 |
| P2 | 69.50 ± 0.47 | 17.29 ± 0.21 | 7.05 ± 0.13 | 4.96 ± 0.06 |
| P3 | 70.36 ± 0.52 | 17.14 ± 0.26 | 6.50 ± 0.21 | 4.79 ± 0.05 |
| P4 | 69.25 ± 0.16 | 17.73 ± 0.19 | 7.03 ± 0.18 | 4.94 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hua, L.; Zhang, P.; Liu, H.; Xin, M.; Zhang, Z.; Han, D.; Zhang, Z.; Zhu, X.; Jin, J.; Yang, Y.; et al. Effects of Dietary Inosine 5′-Monophosphate Supplementation on the Growth Performance and Salinity and Oxidative Stress Resistance of Gibel Carp (Carassius auratus gibelio). Antioxidants 2024, 13, 487. https://doi.org/10.3390/antiox13040487
Hua L, Zhang P, Liu H, Xin M, Zhang Z, Han D, Zhang Z, Zhu X, Jin J, Yang Y, et al. Effects of Dietary Inosine 5′-Monophosphate Supplementation on the Growth Performance and Salinity and Oxidative Stress Resistance of Gibel Carp (Carassius auratus gibelio). Antioxidants. 2024; 13(4):487. https://doi.org/10.3390/antiox13040487
Chicago/Turabian StyleHua, Luohai, Peiyu Zhang, Haokun Liu, Mingze Xin, Zhiwei Zhang, Dong Han, Zhimin Zhang, Xiaoming Zhu, Junyan Jin, Yunxia Yang, and et al. 2024. "Effects of Dietary Inosine 5′-Monophosphate Supplementation on the Growth Performance and Salinity and Oxidative Stress Resistance of Gibel Carp (Carassius auratus gibelio)" Antioxidants 13, no. 4: 487. https://doi.org/10.3390/antiox13040487
APA StyleHua, L., Zhang, P., Liu, H., Xin, M., Zhang, Z., Han, D., Zhang, Z., Zhu, X., Jin, J., Yang, Y., & Xie, S. (2024). Effects of Dietary Inosine 5′-Monophosphate Supplementation on the Growth Performance and Salinity and Oxidative Stress Resistance of Gibel Carp (Carassius auratus gibelio). Antioxidants, 13(4), 487. https://doi.org/10.3390/antiox13040487