
Suitable Cottonseed Protein Concentrate Supplementation in Common Carp (Cyprinus carpio) Serves as an Effective Strategy for Fish Meal Sparing Based on Improvement in Intestinal Antioxidant Capacity, Barrier and Microbiota Composition
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Design and Production Process of Experimental Diets
2.2. Design and Management of the Feeding Trial
2.3. Sample Acquisition and Analysis Methods
2.3.1. Calculation of Growth and Feed Utilization
2.3.2. Detection of Serum Biochemical Indexes
2.3.3. Analysis of the Intestinal Activities of Digestion and Antioxidant Enzymes
2.3.4. Intestinal mRNA Expression Analysis
2.3.5. Histological Observation of the Intestine
2.3.6. Intestinal Microbiota Analysis Procedure
2.4. Design and Preparation of Experimental Diets
3. Results
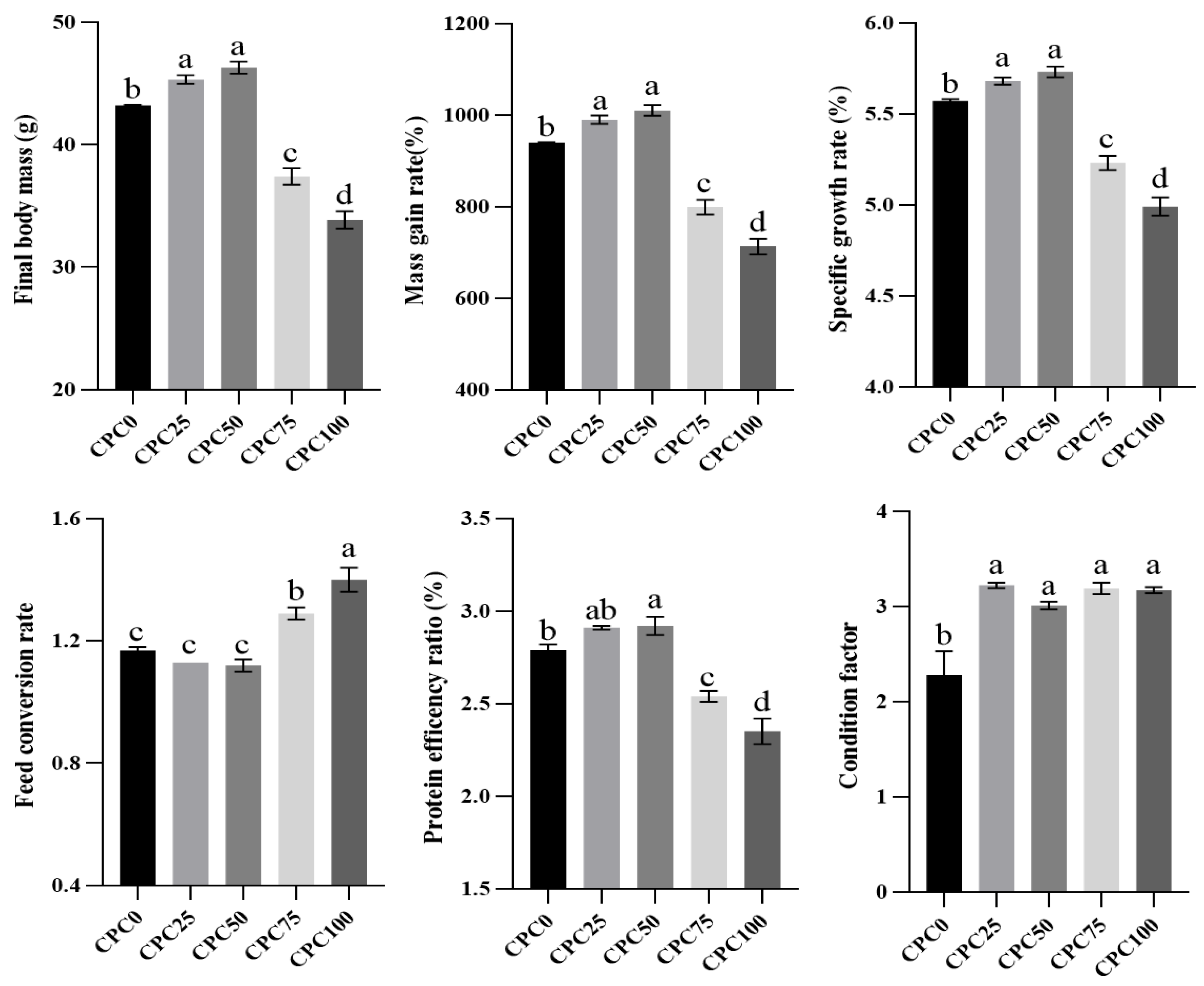
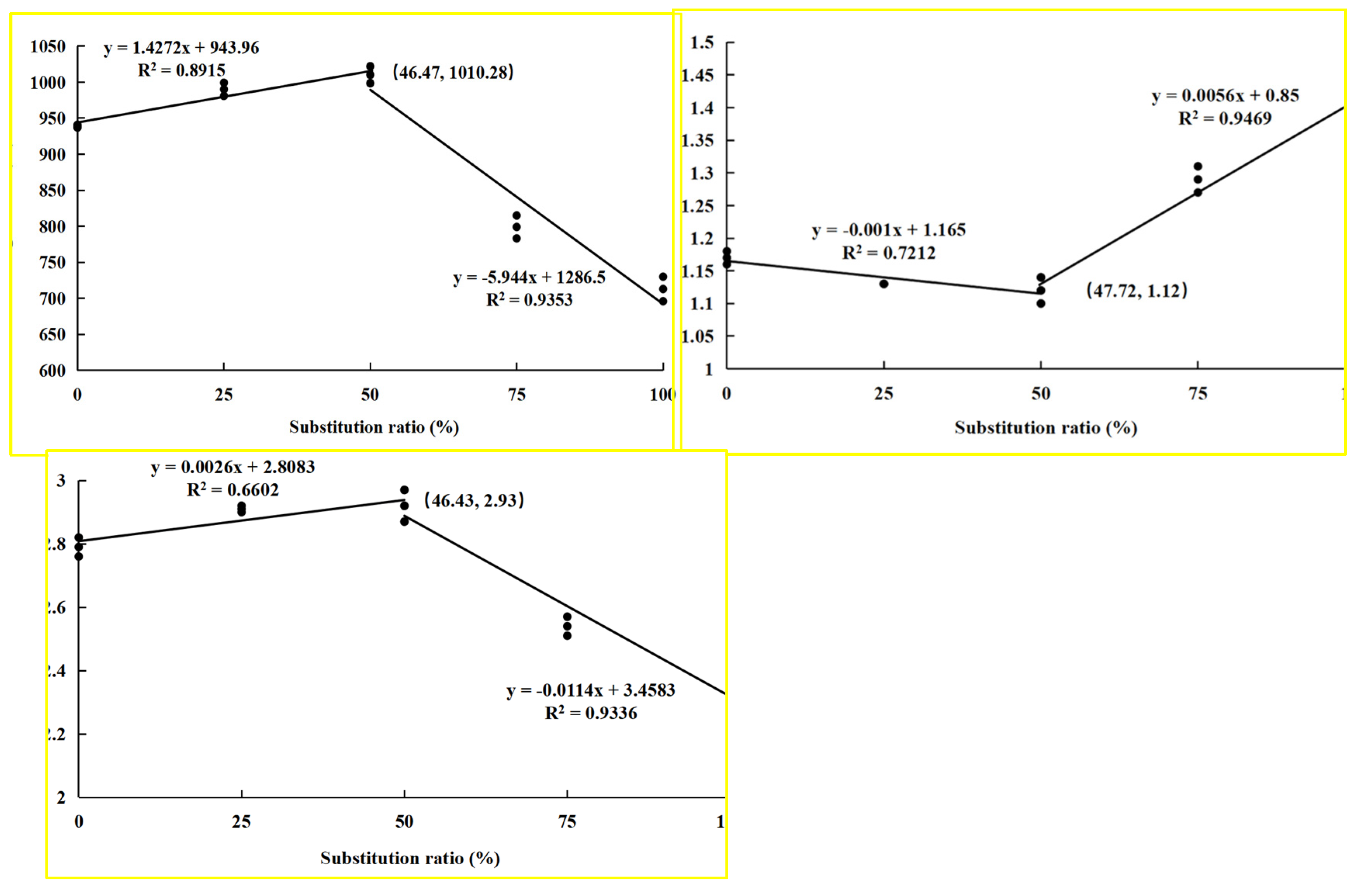
3.1. Parameters of Growth and Feed Utilization
3.2. Biochemical Components of the Serum
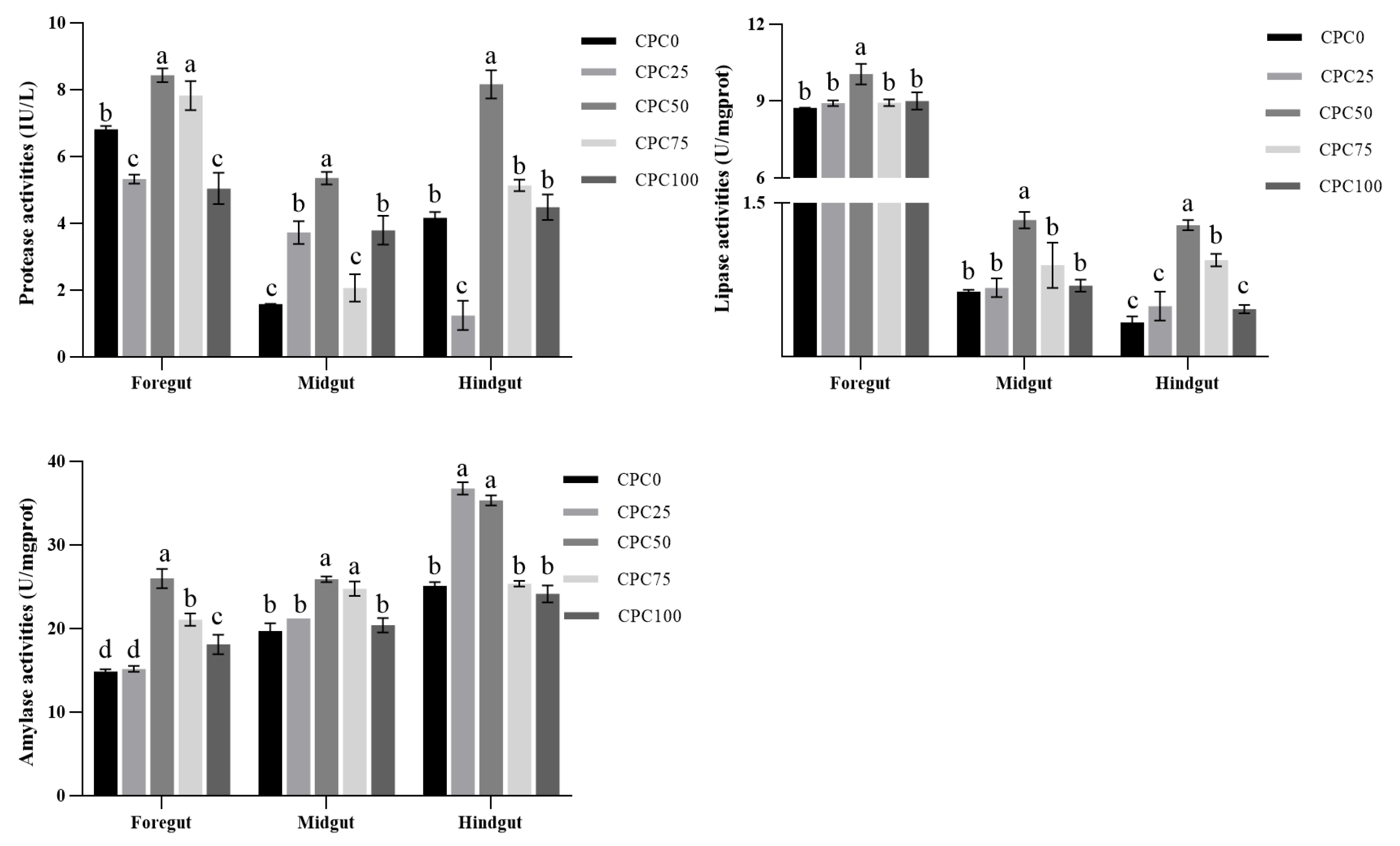
3.3. Intestinal Digestive Enzyme Activities
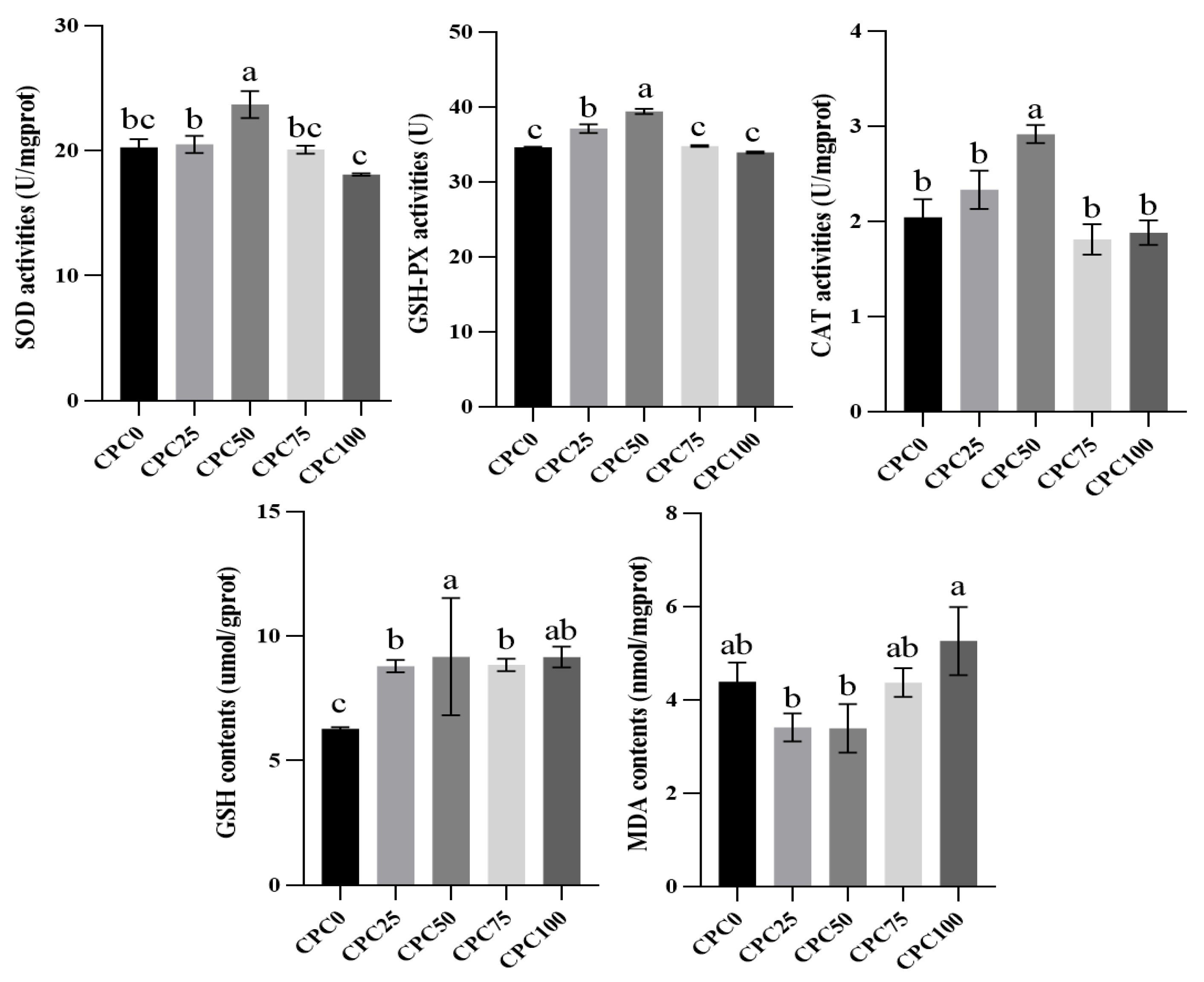
3.4. Intestinal Antioxidant Indexes Evaluation
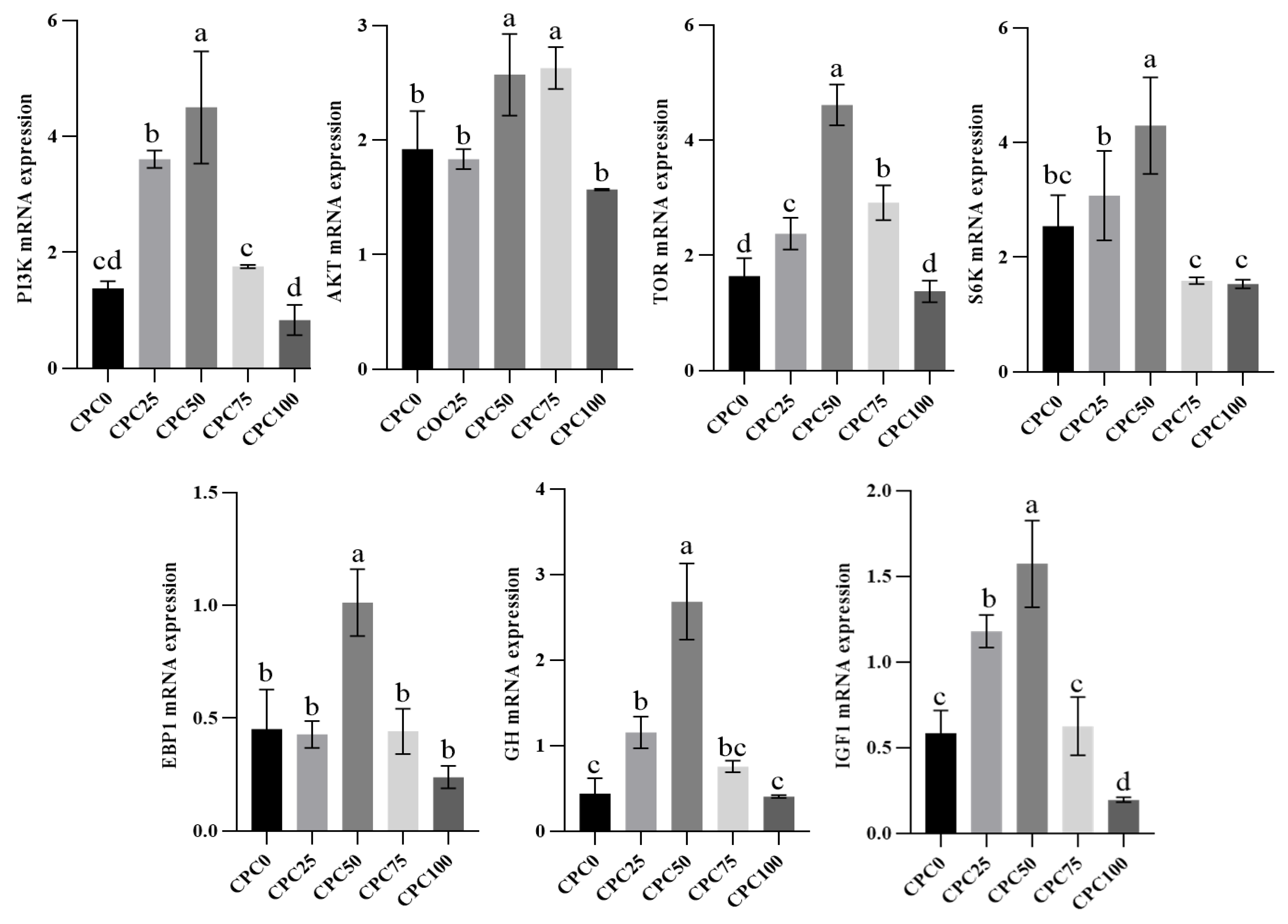
3.5. Intestinal Relative Expressions of Growth Axis and Protein Synthesis Genes
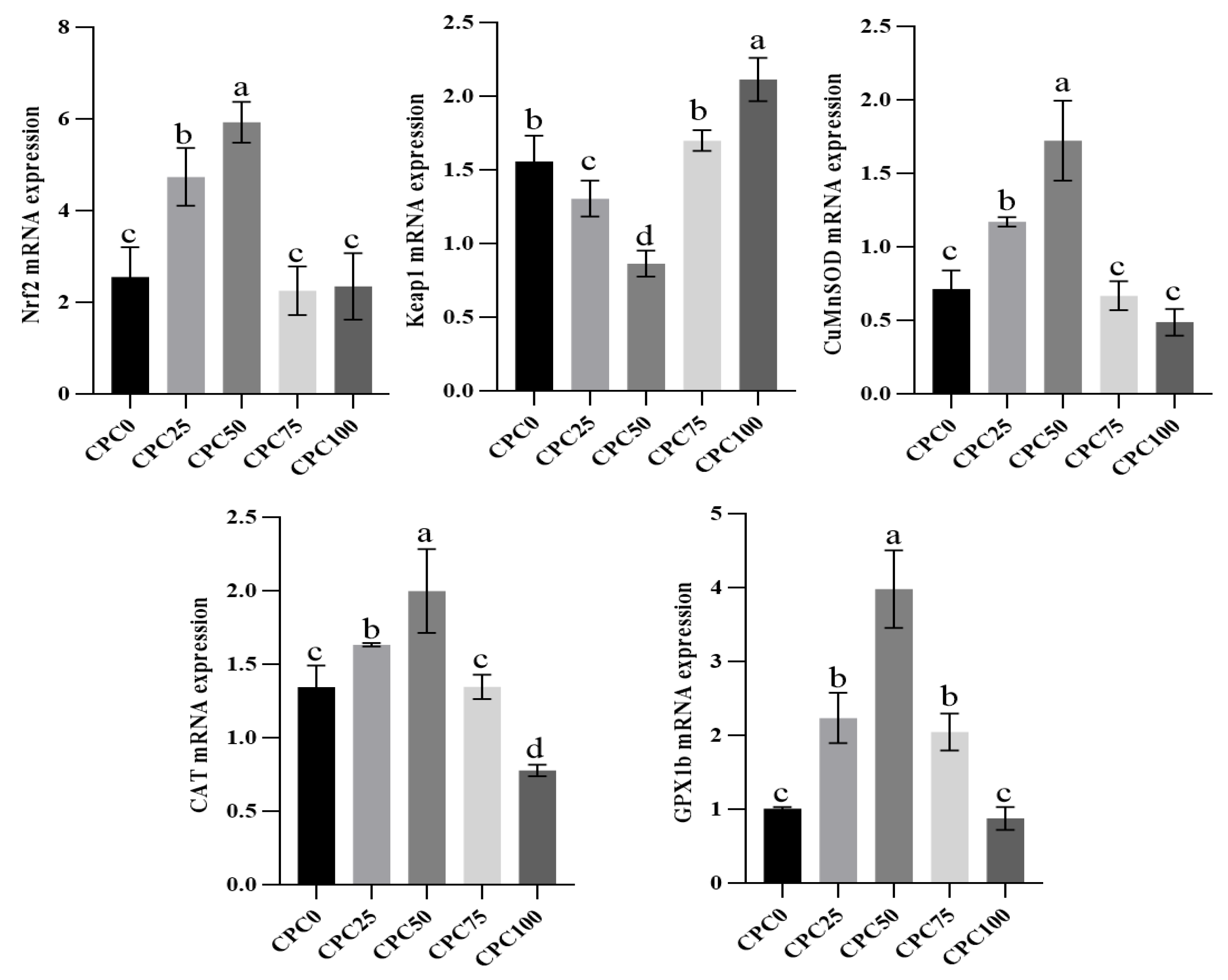
3.6. Intestinal Relative Expressions of Antioxidation Genes
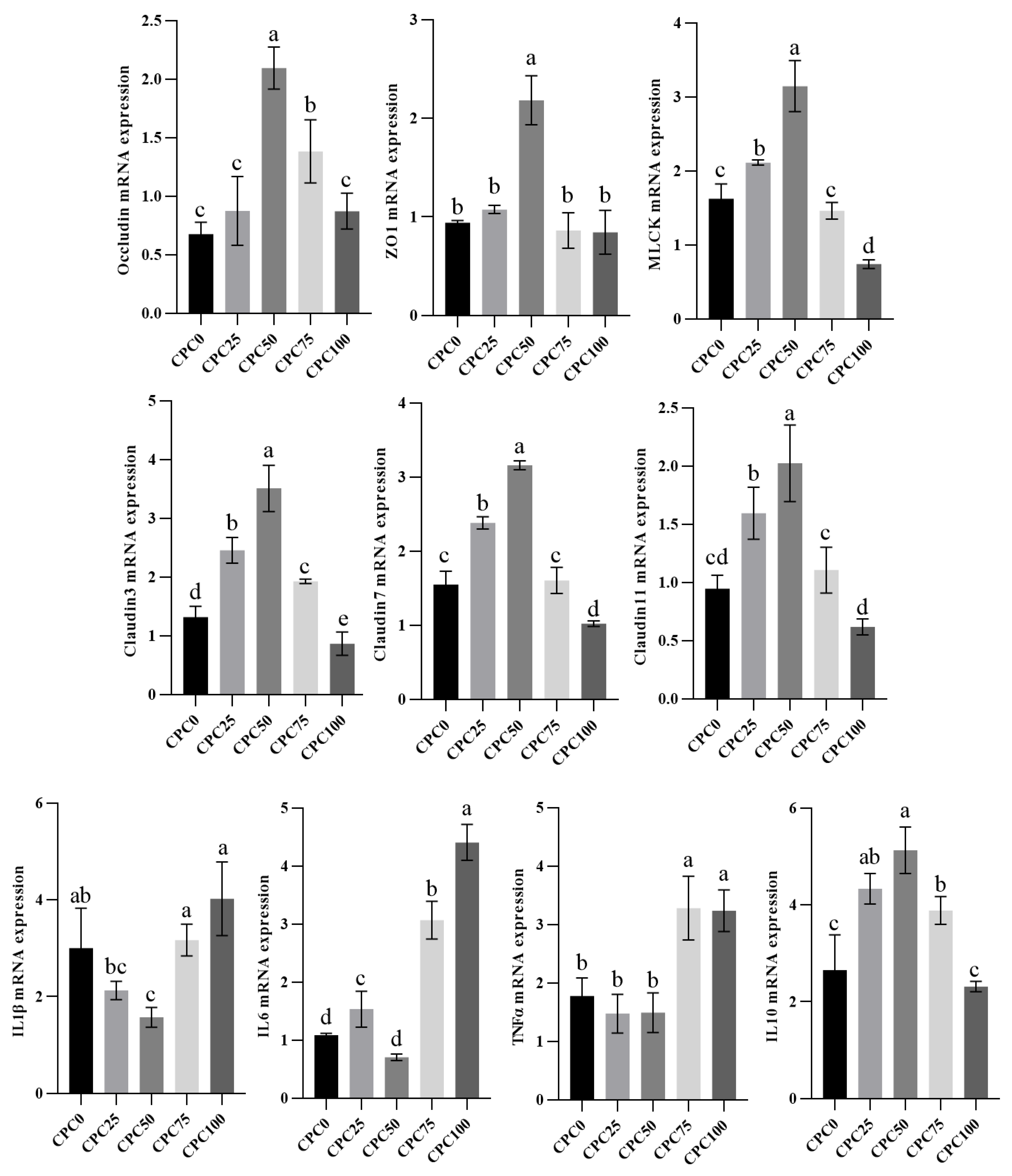
3.7. Intestinal Relative Expressions of Immune Barrier and Cytokines Genes
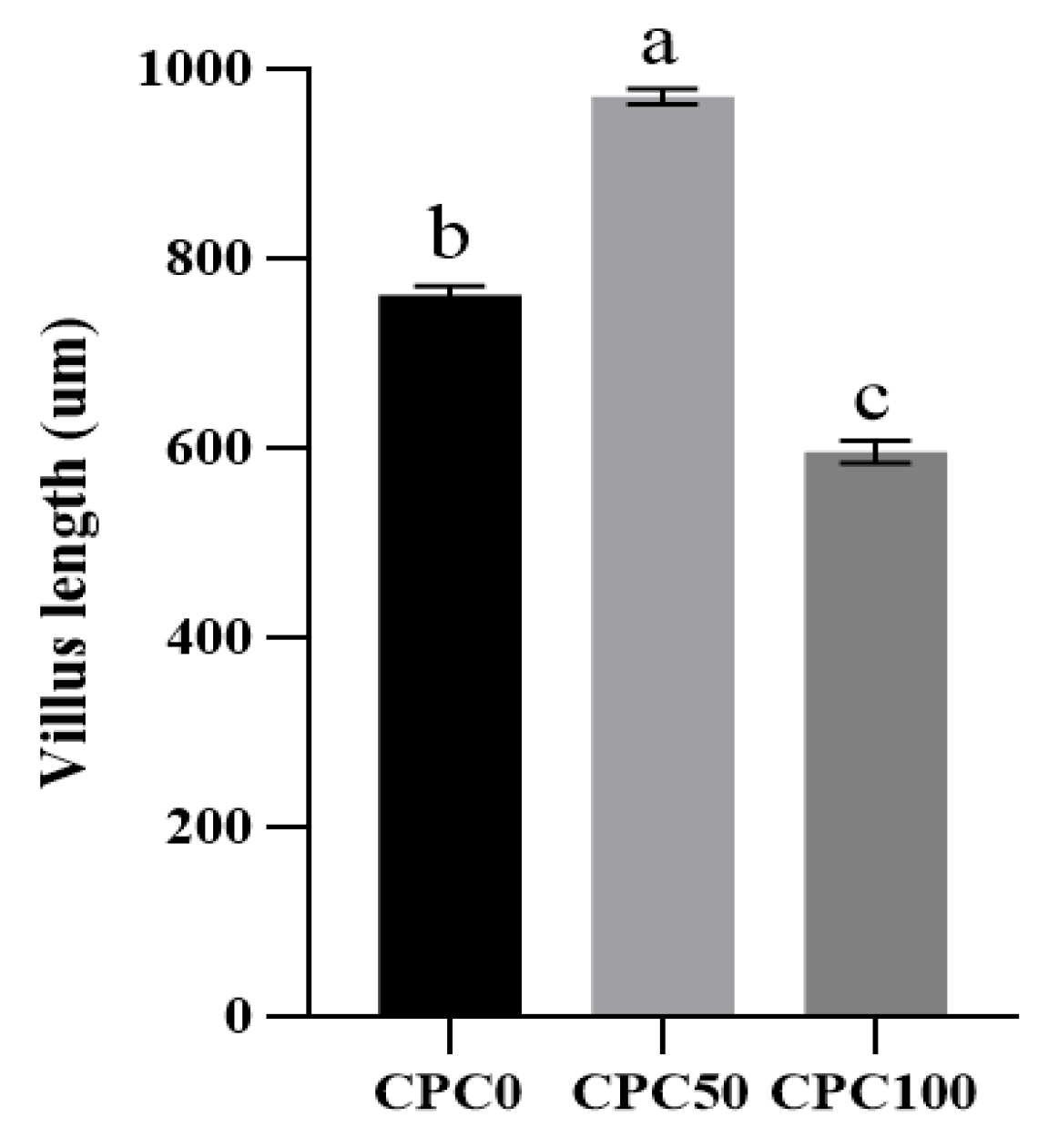
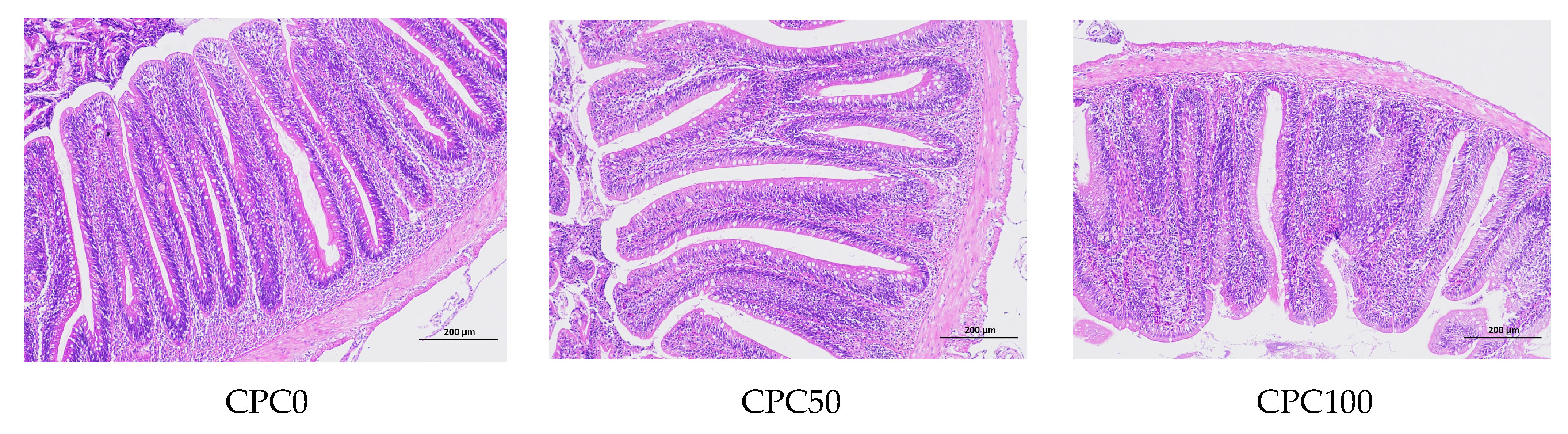
3.8. Histological Analysis of the Midguts
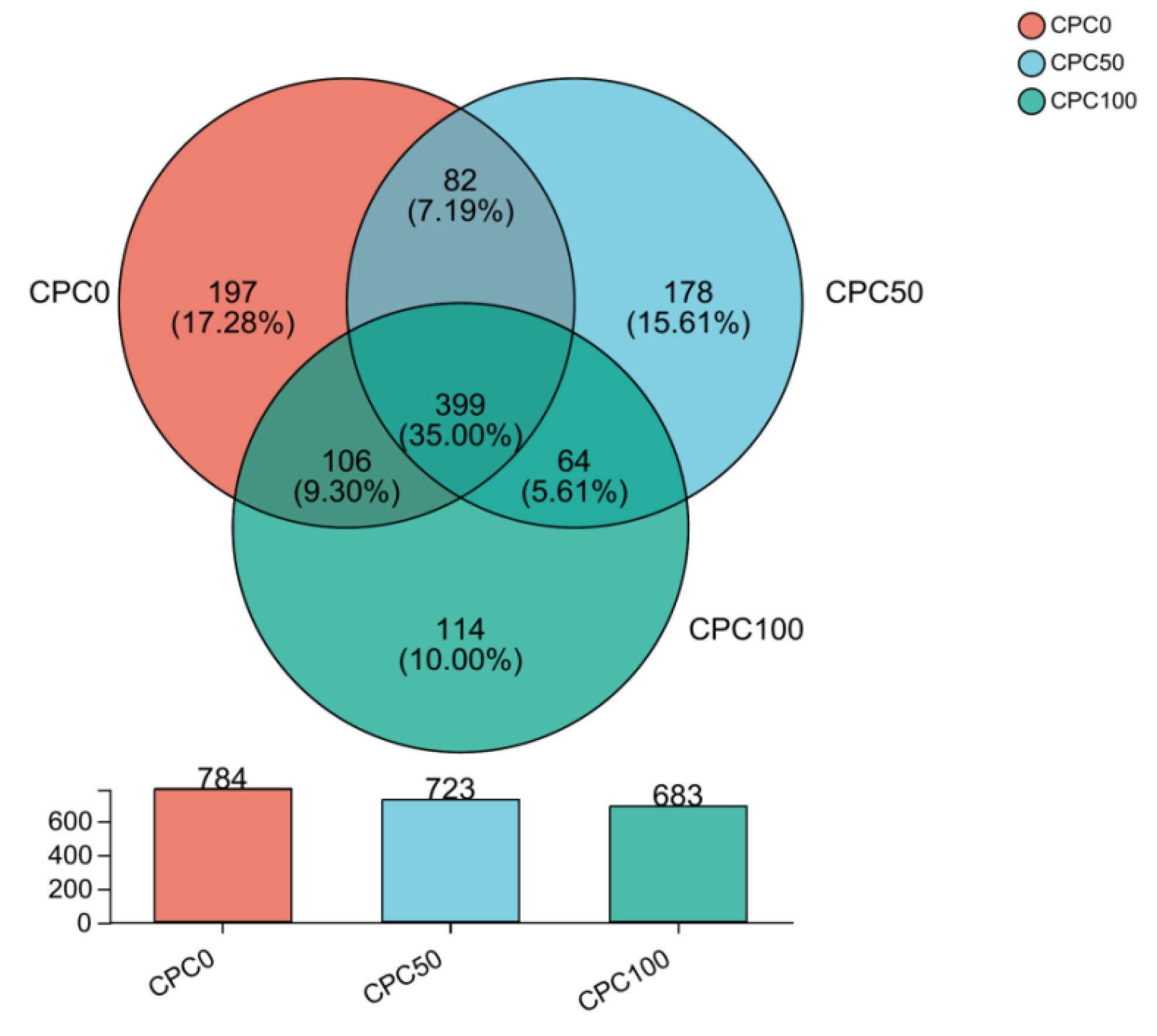
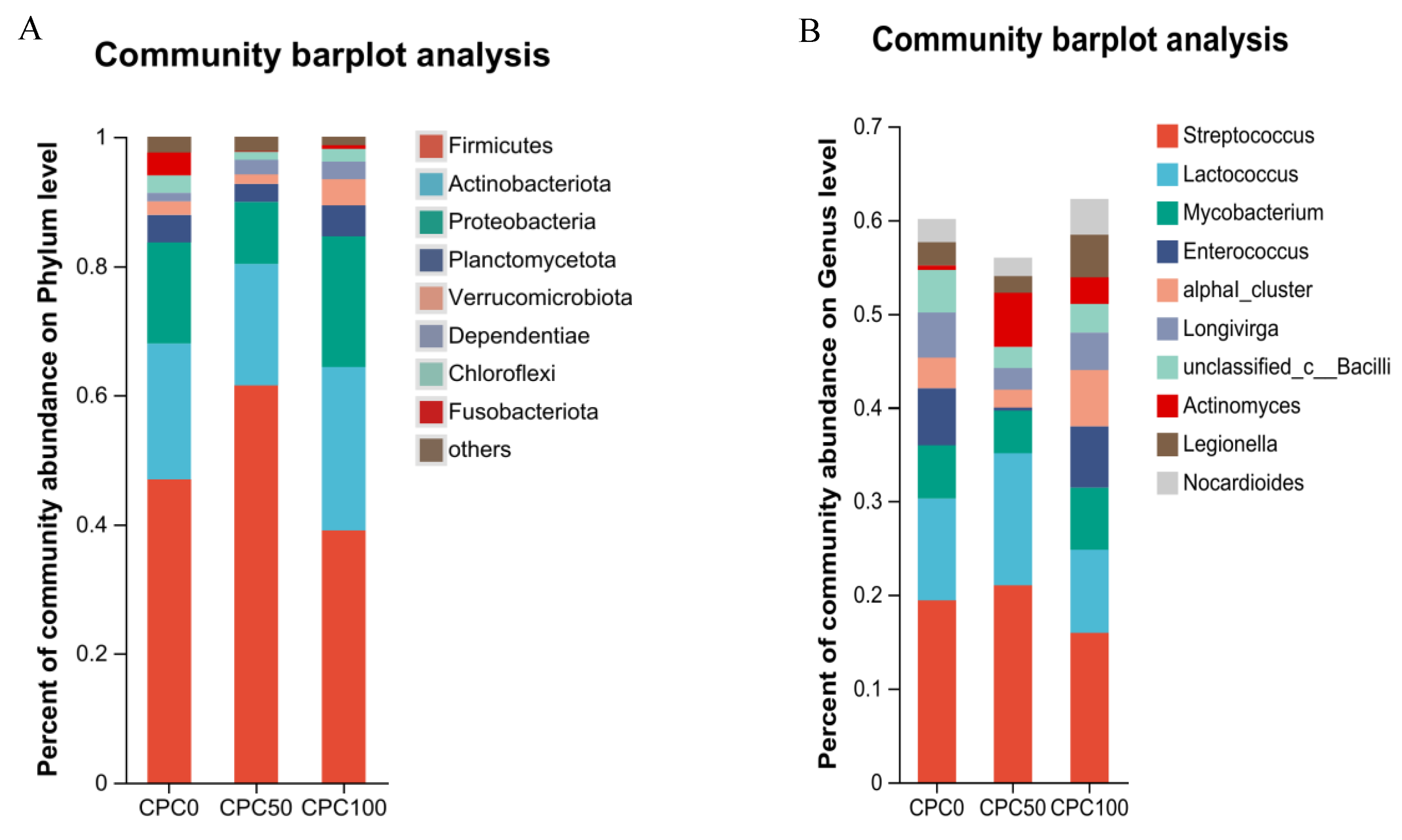
3.9. Intestinal Microbiota Analyses
4. Discussion
4.1. Suitable Substitution of Fishmeal with CPC Contributes to the Growth-Promoting Effect
4.2. Suitable Substitution of Fishmeal with CPC Improves Serum Biochemical Composition
4.3. Suitable Substitution of Fishmeal with CPC Elevates Intestinal Digestive and Absorptive Ability
4.4. Suitable Substitution of Fishmeal with CPC Remits the Weakening Effect of Reducing Fiashmeal on the Intestinal Antioxidant Capacity and Immune Function
4.5. Suitable Substitution of Fishmeal with CPC Enhances the Intestinal Mechanical Barrier and Biological Barrier
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Horvath, L.; Hegyi, A.; Lefler, K.-K.; Csorbai, B.; Kovacs, E.; Szabo, T.; Muller, T.; Urbanyi, B. Review of Central-Eastern European Propagation and Larvae Nursing Method for Common Carp (Cyprinus carpio L.). Life 2023, 13, 2334. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.; Gilligan, D. Optimising an integrated pest-management strategy for a spatially structured population of common carp (Cyprinus carpio) using meta-population modelling. Mar. Freshw. Res. 2014, 65, 538–550. [Google Scholar] [CrossRef]
- Weber, M.-J.; Hennen, M.-J.; Brown, M.-L. Simulated population responses of common carp to commercial exploitation. N. Am. J. Fish. Manag. 2011, 31, 269–279. [Google Scholar] [CrossRef]
- Food and Agriculture Organization, (FAO). The State of World Fisheries and Aquaculture: Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Shi, D.-H.; Ye, Y.-T. Statistical analysis of nutritional quality of common carp compound feed. Feed Res. 2019, 42, 73–77. [Google Scholar]
- Chiu, A.; Li, L.-P.; Guo, S.-J.; Bai, J.-F.; Fedor, C.; Naylor, R.-L. Feed and fishmeal use in the production of carp and tilapia in China. Aquaculture 2013, 414, 127–134. [Google Scholar] [CrossRef]
- Fan, Z.; Cheng, M.; Wang, L.-S.; Li, C.-H.; Wu, D.; Li, J.-N.; Cheng, Z.-Y.; Zhang, H.-T.; Li, W.-Q. Feasibility evaluation of fermented peanut meal to replace soybean meal in the diet of common carp (Cyprinus carpio): Growth performance, serum biochemistry, intestinal health and microflora composition. Aquac. Rep. 2023, 31, 101675. [Google Scholar] [CrossRef]
- Hussain, S.-M.; Bano, A.-A.; Ali, S.; Rizwan, M.; Adrees, M.; Zahoor, A.-F.; Sarker, P.-K.; Hussain, M.; Arsalan, M.-Z.; Yong, J.-W.-H.; et al. Substitution of fishmeal: Highlights of potential plant protein sources for aquaculture sustainability. Heliyon 2024, 10, e26573. [Google Scholar] [CrossRef] [PubMed]
- van Riel, A.-J.; Nederlof, M.-A.-J.; Chary, K.; Wiegertjes, G.-F.; de Boer, I.-J.-M. Feed-food competition in global aquaculture: Current trends and prospects. Rev. Aquac. 2023, 15, 1142–1158. [Google Scholar] [CrossRef]
- Aragão, C.; Gonçalves, A.T.; Costas, B.; Azeredo, R.; Xavier, M.J.; Engrola, S. Alternative proteins for fish diets: Implications beyond growth. Animals 2022, 12, 1211. [Google Scholar] [CrossRef]
- Fu, H.; Song, H.-M.; Mu, X.-D.; Tian, X.; Liu, C.; Liu, Y.; Yang, Y.-X. Effects of fish meal replacement with fermented Antarctic krill meal on growth performance, body color and serum biochemical indexes of koi carp (Cyprinus carpio L.). Prog. Fish. Sci. 2023, 44, 80–89. [Google Scholar]
- Wu, D.; Zhang, Y.-Y.; Li, J.-N.; Fan, Z.; Xu, Q.-Y.; Wang, L.-S. Assessment of chicken intestinal hydrolysates as a new protein source to replace fishmeal on the growth performance, antioxidant capacity and intestinal health of common carp (Cyprinus carpio). Fish Shellfish Immunol. 2022, 125, 161–170. [Google Scholar] [CrossRef]
- Cheng, X.-F.; Wu, H.; Gao, J.-W.; Tian, X.; Xiang, J.; He, Z.-G.; Zou, L.; Song, R.; Li, C.-W. Influence of dietary replacement of fish meal with crayfish shell meal on the skin coloration, muscle texture, and amino acid composition of Oujiang color common carp Cyprinus carpio var. color. N. Am. J. Aquac. 2021, 83, 402–408. [Google Scholar] [CrossRef]
- Xue, M. Processing of cottonseed protein concentrated and its nutrient values in aquatic feed. Feed Ind. 2021, 42, 1–5. [Google Scholar]
- Shen, J.-F.; Liu, H.-Y.; Tan, B.-P.; Dong, X.-H.; Yang, Q.-H.; Chi, S.-Y.; Zhang, S. Effects of replacement of fishmeal with cottonseed protein concentrate on the growth, intestinal microflora, haematological and antioxidant indices of juvenile golden pompano (Trachinotus ovatus). Aquac. Nutr. 2020, 26, 1119–1130. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Li, X.-Q.; Xu, Z.; Yang, H.; Lin, X.; Leng, X.-J. Replacing fishmeal with cottonseed protein concentrate in practical diet of largemouth bass (Micropterus salmoides): Growth, flesh quality and metabolomics. Aquaculture 2024, 579, 740164. [Google Scholar] [CrossRef]
- Wang, J.; Clark, G.; Ju, M.; Castillo, S.; Gatlin, D.-M. Effects of replacing menhaden fishmeal with cottonseed flour on growth performance, feed utilization and body composition of juvenile red drum Sciaenops ocellatus. Aquaculture 2020, 523, 735217. [Google Scholar] [CrossRef]
- Li, X.-Q.; Li, H.-D.; Ji, H.; Sun, J.; Hu, Z.-C.; Liu, S. Substitution of soybean meal with cottonseed protein concentrate on growth performance, health status and flesh quality of grass carp (Ctenopharyngodon idella). Acta Hydrobiol. Sin. 2023, 47, 235–248. [Google Scholar]
- Fan, Z.; Li, C.-H.; Wu, D.; Li, J.-N.; Wang, L.-S.; Cao, D.-C.; Miao, L.-H.; Xie, S.-Q. Evaluation of four novel protein sources as alternatives to soybean meal for two specifications of cage-farmed grass carp (Ctenopharyngodon idellus) deeds: Effect on growth performance, flesh quality, and expressions of muscle-related genes. Front. Mar. Sci. 2022, 9, 935651. [Google Scholar] [CrossRef]
- Fan, Z.; Li, C.-H.; Wu, D.; Li, J.-N.; Liu, Q.-W.; Wang, L.; Wang, L.-S. Application effect of two new plant protein substitutes for soybean meal in large-size common carp feed. Chin. J. Fish. 2023, 36, 20–28+37. [Google Scholar]
- Fan, Z.; Wu, D.; Li, J.-N.; Li, C.-H.; Zheng, X.-H.; Wang, L.S. Phosphorus nutrition in Songpu mirror carp (Cyprinus carpio Songpu) during chronic carbonate alkalinity stress: Effects on growth, intestinal immunity, physical barrier function, and intestinal microflora. Front. Immunol. 2022, 13, 900793. [Google Scholar] [CrossRef]
- Hao, Q.-R.; Wang, P.; Qin, D.-L.; Chen, Z.-X.; Li, C.-H.; Huang, L.; Wu, S.; Yang, J.-X. Exposure to benzo(a)pyrene interfered with cell composition and immune ability of common carp (Cyprinus carpio) intestine through inducing cell heterogeneous responses. Aquaculture 2024, 579, 740229. [Google Scholar] [CrossRef]
- Dawood, M.-A.-O. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, Z.; Zhu, W.-J.; Chen, S.-Y.; Xiao, S.-Y.; Liu, J.; Wang, H.-Q.; Lan, S.-L. Effects of fermented soybean meal instead of fish meal on growth, serum biochemical\indexes and intestinal structure of Carassius auratus. Feed Ind. 2021, 42, 31–37. [Google Scholar]
- Lu, R.-H.; Chen, Y.-N.; Yu, W.-P.; Lin, M.-J.; Yang, G.-K.; Qin, C.-B.; Meng, X.-L.; Zhang, Y.-M.; Ji, H.; Nie, G.-X. Defatted black soldier fly (Hermetia illucens) larvae meal can replace soybean meal in juvenile grass carp (Ctenopharyngodon idellus) diets. Aquac. Rep. 2020, 18, 100520. [Google Scholar] [CrossRef]
- Miyatake, H. Carp. Yoshoku 1997, 34, 108–111. (In Japanese) [Google Scholar]
- Damgaard, M.-V.; Treebak, J.-T. Protocol for qPCR analysis that corrects for cDNA amplification efficiency. STAR Protoc. 2022, 3, 101515. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, L.-S.; Li, J.-N.; Wu, D.; Li, C.-H.; Zheng, X.-H.; Zhang, H.-T.; Miao, L.-H.; Ge, X.-P. Momordica charantia saponins administration in low-protein-high-carbohydrate diet improves growth, blood biochemical, intestinal health and microflora composition of juvenile common carp (Cyprinus carpio). Fish Shellfish Immunol. 2023, 140, 108980. [Google Scholar] [CrossRef]
- Tang, X.-Y.; Zhao, L.-C.; Zhang, X.-Y.; Liu, J.; Jia, Z.W.; Dou, Y.-L. Experimental study on dephenolization of edible cottonseed protein by Liquid-liquid-solid three-phase extraction. Agric. Sci. Technol. Equip. 2019, 3, 45–46. [Google Scholar]
- He, Y.-F.; Guo, X.-W.; Tan, B.-P.; Dong, X.-H.; Yang, Q.-H.; Liu, H.-Y.; Zhang, S.; Chi, S.Y. Replacing fishmeal with cottonseed protein concentrate in feed for pearl gentian groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂): Effects on growth and expressions of key genes involved in appetite and hepatic glucose and lipid metabolism. Aquac. Rep. 2021, 20, 100710. [Google Scholar] [CrossRef]
- Ye, G.-L.; Dong, X.-H.; Yang, Q.-H.; Chi, S.-Y.; Liu, H.-Y.; Zhang, H.-T.; Tan, B.-P.; Zhang, S. Low-gossypol cottonseed protein concentrate used as a replacement of fish meal for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Effects on growth performance, immune responses and intestinal microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, S.-W.; Lv, W.-H.; Shi, H.-H.; Qiu, G.-W.; Chang, H.-M.; Lu, S.-X.; Wang, D.; Wang, C.-A.; Han, S.-C.; et al. Effects of replacing fishmeal with cottonseed protein concentrate on growth performance, blood metabolites, and the intestinal health of juvenile rainbow trout (Oncorhynchus mykiss). Front. Immunol. 2022, 13, 1079677. [Google Scholar] [CrossRef]
- Li, W.-J.; Wu, H.-X.; Zhang, L.; Li, M.; Wang, T.; Shan, C.-J.; Qiao, F.; Chen, L.-Q.; Zhang, W.-B.; Du, Z.-Y.; et al. Effects of replacing soybean meal protein with cottonseed protein concentrate on the growth condition and intestinal health of Nile tilapia (Oreochromis niloticus). Aquac. Nutr. 2021, 27, 2436–2447. [Google Scholar] [CrossRef]
- Vélez, E.-J.; Lutfi, E.; Perelló, S.-A.-M.; Salmerón, C.; Riera-Codina, M.; Ibarz, A.; Fernández-Borràs, J.; Blasco, J.; Capilla, E.; Navarro, I.; et al. Understanding fish muscle growth regulation to optimize aquaculture production. Aquaculture 2017, 467, 28–40. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Yan, X.-H. Sensors for the mTORC1 pathway regulated by amino acids. J. Zhejiang Univ. Sci. B 2019, 20, 699–712. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, Q.; Guo, M.; Li, F.; Qin, M.; Xie, Y.; Xu, J.; Liu, Y.; Tong, T. The effects of dietary fermented soybean meal supplementation on the growth, antioxidation, immunity, and mTOR signaling pathway of juvenile coho salmon (Oncorhynchus kisutch). Fishes 2023, 8, 448. [Google Scholar] [CrossRef]
- Xie, S.-C.; Zhou, Q.-C.; Zhang, X.-S.; Zhu, T.-T.; Guo, C.; Yang, Z.; Luo, J.-X.; Yuan, Y.; Hu, X.-Y.; Jiao, L.-F.; et al. Effect of dietary replacement of fish meal with low-gossypol cottonseed protein concentrate on growth performance and expressions of genes related to protein metabolism for swimming crab (Portunus trituberculatus). Aquaculture 2022, 549, 737820. [Google Scholar] [CrossRef]
- Wang, K.-Z.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhang, Y.-A.; Zhou, X.-Q.; Feng, L. Gossypol reduced the intestinal amino acid absorption capacity of young grass carp (Ctenopharyngodon idella). Aquaculture 2018, 492, 46–58. [Google Scholar] [CrossRef]
- Chantranupong, L.; Scaria, S.-M.; Saxton, R.-A.; Gygi, M.-P.; Shen, K.; Wyant, G.-A.; Sabatini, D.-M. The CASTOR proteins are arginine sensors for the mTORC1 pathway. Cell 2016, 165, 153–164. [Google Scholar] [CrossRef]
- Columbus, D.-A.; Fiorotto, M.-L.; Davis, T.-A. Leucine is a major regulator of muscle protein synthesis in neonates. Amino Acids 2015, 47, 259–270. [Google Scholar] [CrossRef]
- Esmaeili, N. Blood Performance: A New Formula for Fish Growth and Health. Biology 2021, 10, 1236. [Google Scholar] [CrossRef]
- Shen, J.-F.; Chen, M.-C.; Yang, Q.-H.; Liu, H.-Y.; Chi, S.-Y.; Tan, B.-P.; Wang, H.-L. Effects of fish meal replacement by concentrated cottonseed protein on growth performance, serum biochemical indices, liver antioxidant indices and gastrointestinal tract protease activities of juvenile Trachinotus ovatus. Chin. J. Anim. Nutr. 2019, 31, 746–756. [Google Scholar]
- Zheng, S.; Shi, Y.; Zhang, J.; Dai, J.; Hu, Y.; Zhong, L. Effects of Replacing Fish Meal with Stickwater Hydrolysate and Meal on the Growth, Serum Biochemical Indexes, and Muscle Quality of Yellow Catfish (Tachysurus fulvidraco). Fishes 2023, 8, 566. [Google Scholar] [CrossRef]
- Chen, H.-J.; Luom, D.-J. Application of haematology parameters for health management in fish farms. Rev. Aquac. 2023, 15, 704–737. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Yang, H.; Zhang, C.-Y.; Bian, Y.-H.; Yao, W.-X.; Xu, Z.; Wang, Y.-Y.; Li, X.-Q.; Leng, X.-J. Effects of replacing fishmeal with cottonseed protein concentrate on growth performance, flesh quality and gossypol deposition of largemouth bass (Micropterus salmoides). Aquaculture 2022, 548, 737551. [Google Scholar] [CrossRef]
- Liu, H.-K.; Yan, Q.-G.; Han, D.; Jin, J.-Y.; Zhu, X.-M.; Yang, Y.-X.; Xie, S.-Q. Effect of dietary cottonseed meal on growth performance, physiological response, and gossypol accumulation in pre-adult grass carp, Ctenopharyngodon idellus. Chin. J. Oceanol. Limnol. 2016, 34, 992–1003. [Google Scholar] [CrossRef]
- Yang, L.; Cai, M.; Zhong, L.; Shi, Y.; Xie, S.; Hu, Y.; Zhang, J. Effects of replacing soybean meal protein with Chlorella vulgaris powder on the growth and intestinal health of grass carp (Ctenopharyngodon idella). Animals 2023, 13, 2274. [Google Scholar] [CrossRef]
- Li, X. Histoembryology of Aquatic Animals; China Agriculture Press: Beijing, China, 2006. [Google Scholar]
- Luo, L.; Liu, S.-Y.; Chen, B.-P.; Li, F.-L.; Yongqiang Deng, Y.-Q.; Huang, X.-L.; Geng, Y.; Ouyang, P.; Chen, D.-F. Chronic ammonia stress caused disorder of intestinal microbiota and damaged intestinal structure and function in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2024, 581, 740428. [Google Scholar] [CrossRef]
- Tian, S.-J.; Wu, Y.; Yuan, J.; Zhang, Z.-Y.; Huang, D.; Zhou, H.H.; Zhang, W.-B.; Mai, K.-S. Replacement of dietary fishmeal by cottonseed protein concentrate on growth performance, feed utilization and protein metabolism of large yellow croaker Larimichthys crocea. Aquac. Rep. 2022, 26, 101313. [Google Scholar] [CrossRef]
- Liu, X.-R.; Han, B.; Xu, J.; Zhu, J.-Y.; Hu, J.-T.; Wan, W.-L.; Miao, S.-Y. Replacement of fishmeal with soybean meal affects the growth performance, digestive enzymes, intestinal microbiota and immunity of Carassius auratus gibelio ♀ × Cyprinus carpio ♂. Aquac. Rep. 2020, 18, 100472. [Google Scholar] [CrossRef]
- Fronte, B.; Licitra, R.; Bibbiani, C.; Casini, L.; De Zoysa, M.; Miragliotta, V.; Sagona, S.; Coppola, F.; Brogi, L.; Abramo, F. Fishmeal replacement with Hermetia illucens meal in aquafeeds: Effects on zebrafish growth performances, intestinal morphometry, and enzymology. Fishes 2021, 6, 28. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, Z.; Lu, S.; Liu, Y.; Han, S.; Jiang, H.; Yang, Y.; Liu, H. Physiological, mutritional and transcriptomic responses of sturgeon (Acipenser schrenckii) to complete substitution of fishmeal with cottonseed protein concentrate in aquafeed. Biology 2023, 12, 490. [Google Scholar]
- Lee, J.-W.; Jo, A.-H.; Lee, D.-C.; Choi, C.-Y.; Kang, J.-C.; Kim, J.-H. Review of cadmium toxicity effects on fish: Oxidative stress and immune responses. Environ. Res. 2023, 236, 116600. [Google Scholar] [CrossRef]
- Circu, M.-L.; Aw, T.-Y. Intestinal redox biology and oxidative stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, D.; Li, J.; Zhang, Y.-Y.; Cui, Z.-Y.; Li, T.-B.; Zheng, X.-H.; Liu, H.-B.; Wang, L.-S.; Li, H.-Q. Assessment of Fish Protein Hydrolysates in Juvenile Largemouth Bass (Micropterus salmoides) Diets: Effect on Growth, Intestinal Antioxidant Status, Immunity, and Microflora. Front. Nutr. 2020, 9, 816341. [Google Scholar] [CrossRef]
- Lian, X.; Shi, M.; Liang, Y.; Lin, Q.; Zhang, L. The Effects of Unconventional Feed Fermentation on Intestinal Oxidative Stress in Animals. Antioxidants 2024, 13, 305. [Google Scholar] [CrossRef]
- Zhu, Z.; Kou, S.; Zhang, X.; Lin, Y.; Chi, S.; Yang, Q.; Tan, B. Evaluation of corn distillers dried grains with solubles (DDGS) replacement for fishmeal in the diet for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Aquac. Rep. 2022, 25, 101224. [Google Scholar] [CrossRef]
- Mohammadi, M.; Imani, A.; Farhangi, M.; Gharaei, A.; Hafezieh, M. Replacement of fishmeal with processed canola meal in diets for juvenile Nile tilapia (Oreochromis niloticus): Growth performance, mucosal innate immunity, hepatic oxidative status, liver and intestine histology. Aquaculture 2020, 518, 734824. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, D.; Li, C.-H.; Li, J.-N.; Liu, Q.-W.; Wang, L.-S. Effects of bitter melon Momordica charantia saponins on growth, digestion and hepatopancreas health of juvenile Songpu mirror carp (Cyprinus carpio Songpu) fed high-carbohydrate diets. Chin. J. Fish. 2023, 36, 29–37. [Google Scholar]
- Dinu, D.; Marinescu, D.; Munteanu, M.C.; Staicu, A.C.; Costache, M.; Dinischiotu, A. Modulatory effects of deltamethrin onantioxidant defense mechanisms and lipid peroxidation in Carassius auratus gibelio liver and intestine. Archives of Environ. Contam. Toxicol. 2010, 58, 757–764. [Google Scholar] [CrossRef]
- Yin, B.; Liu, H.-Y.; Tan, B.-P.; Dong, X.-H.; Chi, S.-Y.; Yang, Q.-H.; Zhang, S.; Chen, L.-Q. Cottonseed protein concentrate (CPC) suppresses immune function in different intestinal segments of hybrid grouper ♀ Epinephelus fuscoguttatus × ♂Epinephelus lanceolatu via TLR-2/MyD88 signaling pathways. Fish Shellfish Immunol. 2018, 81, 318–328. [Google Scholar] [CrossRef]
- Zhou, J.-H.; Liu, N.; Yang, Y. Keap1-Nrf2/Are signaling pathway and its antioxidant regulation in teleost fish: A review. Chin. J. Fish. 2021, 35, 117–124. [Google Scholar]
- Medina-F’elix, D.; Garibay-Valdez, E.; Vargas-Albores, F.; Martínez-Porchas, M. Fish disease and intestinal microbiota: A close and indivisible relationship. Rev. Aquac. 2023, 15, 820–839. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Li, B.-S.; Sun, Y.-Z.; Wang, C.-Q.; Huang, B.-S. Research progress of plant protein-induced enteritis of fish and Its repair. J. Guangdong Ocean Univ. 2023, 43, 37–46. [Google Scholar]
- Wu, G.-D.; Lan, K.-P.; Chen, X.; Wang, Y.; Zhou, C.-P.; Lin, H.-Z.; Ma, Z.-H.; Wang, J. Effects of replacement of fish meal by fermented cottonseed meal ongrowth performance, feed utilization and intestinal bacteria community of juvenile golden pompano (Trachinotus ovatus). South China Fish. Sci. 2023, 19, 126–138. [Google Scholar]
- Bernardi, S.; Del Bo’, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andr´es-Lacueva, C.; Nicole Hidalgo-Liberona, N.; Peron, G.; Gonzalez-Dominguez, R.; Kroon, P.; et al. Polyphenols and intestinal permeability: Rationale and future perspectives. J. Agric. Food Chem. 2020, 68, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- KöNig, J.; Wells, J.; Cani, P.-D.; García-Ródenas, C.-L.; Macdonald, T.; Mercenier, A.; Jacqueline, W.; Freddy, T.; Robert-Jan, B. Human intestinal barrier function in health and disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef]
- He, G.; Zhang, T.; Zhou, X.; Liu, X.; Sun, H.; Chen, Y.; Tan, B.; Lin, S. Effects of cottonseed protein concentrate on growth performance, hepatic function and intestinal health in juvenile largemouth bass, Micropterus salmoides. Aquac. Rep. 2022, 23, 101052. [Google Scholar] [CrossRef]
- Zhang, B.-Y.; Yang, H.-L.; Wang, P.; Sun, Y.-Z. Advances in the interactions between intestinal microorganisms and host immune system in fish. Acta Microbiol. Sin. 2021, 61, 3046–3058. [Google Scholar]
- Ingerslev, H.-C.; Strube, M.-L.; Jørgensen, L.-G.; Dalsgaard, I.; Boye, M.; Madsen, L. Diet type dictates the gut microbiota and the immune response against Yersinia ruckeri in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2014, 40, 624–633. [Google Scholar] [CrossRef]
- Xie, X.; Wang, J.; Guan, Y.; Xing, S.-J.; Liang, X.-F.; Xue, M.; Wang, J.-J.; Chang, Y.; Leclecg, E. Cottonseed protein concentrate as fishmeal alternative for largemouth bass (Micropterus salmoides) supplemented a yeast-based paraprobiotic: Effects on growth performance, gut health and microbiome. Aquaculture 2022, 551, 737898. [Google Scholar] [CrossRef]
- Feng, J.-C.; Liu, S.-S.; Zhu, C.-J.; Cai, Z.-L.; Cui, W.-S.; Chang, X.-L.; Yan, X.; Qin, C.-B.; Zhang, J.-X.; Nie, G.-X. The effects of dietary Lactococcus spp. on growth performance, glucose absorption and metabolism of common carp, Cyprinus carpio L. Aquaculture 2022, 546, 737394. [Google Scholar] [CrossRef]
- Lam, M.-M.-C.; Seemann, T.; Bulach, D.-M.; Gladman, S.-L.; Chen, H.; Haring, V.; Moore, R.-J.; Ballard, S.; Grayson, M.-L.; Johnson, P.-D.-R.; et al. Comparative analysis of the first complete Enterococcus faecium genome. J. Bacteriol. 2012, 194, 2334–2341. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | CPC0 | CPC25 | CPC50 | CPC75 | CPC100 |
|---|---|---|---|---|---|
| Soybean meal a | 260 | 260 | 260 | 260 | 260 |
| Wheat middlings a | 417 | 415.5 | 414.3 | 413 | 411.4 |
| Fishmeal a | 100 | 75 | 50 | 25 | 0 |
| Soybean protein concentrate a | 50 | 50 | 50 | 50 | 50 |
| Rapeseed meal a | 100 | 100 | 100 | 100 | 100 |
| Cottonseed protein concentrate a | 0 | 25 | 50 | 75 | 100 |
| Fish oil | 20 | 20.5 | 20.8 | 21.1 | 21.6 |
| Soybean oil | 20 | 20.5 | 20.8 | 21.1 | 21.6 |
| Vitamin premix b | 3 | 3 | 3 | 3 | 3 |
| Choline chloride | 3 | 3 | 3 | 3 | 3 |
| Dicalcium phosphate | 20 | 20 | 20 | 20 | 20 |
| Trace mineral premix c | 2 | 2 | 2 | 2 | 2 |
| Lysine | 3 | 3.4 | 3.9 | 4.4 | 4.9 |
| Methionine | 2 | 2.1 | 2.2 | 2.4 | 2.5 |
| Total | 1000.00 | 1000.00 | 1000.00 | 1000.00 | 1000.00 |
| Moisture | 104.7 | 104.6 | 105.0 | 105.0 | 104.9 |
| Crude protein | 305.7 | 305.5 | 305.4 | 305.2 | 305.0 |
| Crude lipid | 57.8 | 58.0 | 57.9 | 57.7 | 57.9 |
| Crude ash | 57.0 | 57.7 | 58.6 | 59.4 | 60.1 |
| Crude fibre | 58.7 | 57.0 | 55.3 | 53.6 | 51.8 |
| NFE d | 416.0 | 417.0 | 417.9 | 419.2 | 420.3 |
| Crude energy (kJ/g) e | 16.65 | 16.68 | 16.68 | 16.69 | 16.71 |
| Gene Name | Primer Sequence (5′-3′) | Serial Number |
|---|---|---|
| Growth axis | ||
| GH | F: ATCTTCCCTCTGTCTTTCTGC R: AAGTCGGCCAGCTTCTCA | M27000.1 |
| IGF1 | F: AGACAGCCCAAGGACAGCA R: TACAGTGGAGCACATCTCTGGAA | D83272.1 |
| Protein synthesis | ||
| PI3K | F:AAGACCTTCCTCATCACGAC R:CCTTCCACTACAACACTGCA | XM_042758409.1 |
| AKT | F:GGTGTGTTCAAGTTCACCGTCT R: TCCTCACCCAGCTCTCCA | XM_042722896.1 |
| TOR | F:CCACAACGCAGCCAACAA R:GCCACAGAATAGCAACCCT | AF119837.1 |
| S6K | F:GCCAATCTCAGCGTTCTCAAC R:CTGCCTAACATCATCCTCCTT | EF373664.1 |
| Antioxidation | ||
| Nrf2 | F: TTCCCGCTGGTTTACCTTAC R: CGTTTCTTCTGCTTGTCTTT | JX462955 |
| Keap1 | F: CTACAACCCCGAGAGACGA R: GGAGGAGATGAAGCTCCAGAC | JX470752 |
| CuMnSOD | F: TGGCGAAGAAGGCTGTTTGT R: TTCACTGGAGACCCGTCACT | JF342355 |
| CAT | F: CTGGAAGTGGAATCCGTTTG R: CGACCTCAGCGAAATAGTTG | JF411604 |
| GPX1a | F: GTGACGACTCTGTGTCCTTG R: AACCTTCTGCTGTATTCTCTTGA | JF411605 |
| GPX1b | F: TATGTCCGTCCTGGCAATGG R: ATCGCTGGGAATGGAAGTT | JF411606 |
| Immune barrier | ||
| occludin | F: ATCGGTTCAGTACAATCAGG R: GACAATGAAGCCCATAACAA | KF975606 |
| ZO-1 | F:GCCTGCCTACACTCAACCACAAC R:CTGCTTCGGCTGGAGGAGGAG | KY290394.1 |
| Claudin3 | F: GCACCAACTGTATCGAGGATG R: GGTTGTAGAAGTCCCGAATGG | JQ767157 |
| Claudin7 | F: CTTCTATAACCCCTTCACACCAG R: ACATGCCTCCACCCATTATG | JQ767155 |
| Claudin11 | F: TCGGAAGTGAACCAGAAAGC R: GAAGCCAAAGGACATCAAGC | JQ767158 |
| MLCK | F: AGCAGTGTGGGCATCAACCT R: CTCCAGCAGGGTCATGATGAG | XM_019076433.1 |
| Cytokines | ||
| IL-1β | F: AACTTCACACTTGAGGAT R: GACAGAACAATAACAACAAC | KC008576 |
| IL-6 | F: TAGGTTAATGAGCAAGAGGA R: AGAGACTGTTGATACTGGAA | AY102633.1 |
| IL-8 | F:AAACTGAGAGTCGACGCATTG R:TTTTCAATGACCTTCTTAACCCAG | EU011243.1 |
| IL-10 | F: GCCAGCATAAAGAACTCG R: CCAAATACTGCTCGATGT | JX524550.1 |
| TNF-α | F: AAGTCTCAGAACAATCAGGAA R: TGCCTTGGAAGTGACATT | AJ311800 |
| Internal reference | ||
| β-actin | F: GGCAGGTCATCACCATCGG R: TTGGCATACAGGTCTTTACGG | M24113.1 |
| Items | CPC0 | CPC25 | CPC50 | CPC75 | CPC100 |
|---|---|---|---|---|---|
| TP (g/L) | 27.00 ± 1.98 c | 42.62 ± 2.41 ab | 48.53 ± 0.28 a | 42.68 ± 0.89 ab | 40.04 ± 3.31 b |
| BUN (mmol/L) | 5.78 ± 0.14 a | 3.75 ± 0.31 c | 3.53 ± 0.33 c | 4.69 ± 0.23 b | 5.99 ± 0.30 a |
| ALT (U/gprot) | 19.28 ± 0.54 c | 19.01 ± 0.39 c | 20.45 ± 0.18 ab | 21.27 ± 0.18 a | 19.64 ± 0.18 bc |
| AST (U/gprot) | 63.55 ± 2.73 c | 69.94 ± 2.31 c | 81.8 ± 2.93 b | 77.8 ± 2.58 b | 109.57 ± 0.72 a |
| TC (mmol/L) | 6.02 ± 0.10 d | 7.57 ± 0.12 a | 7.05 ± 0.02 b | 5.76 ± 0.76 d | 6.46 ± 0.17 c |
| TG (mmol/L) | 7.12 ± 0.31 a | 5.46 ± 0.65 b | 5.13 ± 0.22 b | 7.07 ± 0.43 a | 7.83 ± 0.34 a |
| HDL-C (mmol/L) | 0.57 ± 0.23 e | 1.15 ± 0.23 b | 1.44 ± 0.01 a | 0.91 ± 0.24 c | 0.65 ± 0.22 d |
| LDL-C (mmol/L) | 4.9 ± 0.03 b | 4.49 ± 0.52 b | 4.34 ± 0.2 b | 5.04 ± 0.39 b | 7.9 ± 0.98 a |
| Groups | ACE | CHAO | SOBS | SHANNON | SIMPSON | COVERAGE (%) |
|---|---|---|---|---|---|---|
| CPC0 | 472.83 ± 42.22 | 479.58 ± 40.79 | 426.50 ± 50.14 | 3.59 ± 0.45 | 0.12 ± 0.06 | 99.76 ± 0.01 |
| CPC50 | 459.59 ± 91.13 | 463.59 ± 91.52 | 426.83 ± 87.77 | 3.91 ± 0.87 | 0.09 ± 0.08 | 99.81 ± 0.02 |
| CPC100 | 460.82 ± 57.07 | 462.3 ± 60.32 | 402.83 ± 56.27 | 3.52 ± 0.48 | 0.11 ± 0.05 | 99.74 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Z.; Ge, K.; Wu, D.; Wang, L.; Li, J.; Li, C.; Zhou, M.; Zhang, H.; Miao, L.; Ge, X. Suitable Cottonseed Protein Concentrate Supplementation in Common Carp (Cyprinus carpio) Serves as an Effective Strategy for Fish Meal Sparing Based on Improvement in Intestinal Antioxidant Capacity, Barrier and Microbiota Composition. Antioxidants 2024, 13, 436. https://doi.org/10.3390/antiox13040436
Fan Z, Ge K, Wu D, Wang L, Li J, Li C, Zhou M, Zhang H, Miao L, Ge X. Suitable Cottonseed Protein Concentrate Supplementation in Common Carp (Cyprinus carpio) Serves as an Effective Strategy for Fish Meal Sparing Based on Improvement in Intestinal Antioxidant Capacity, Barrier and Microbiota Composition. Antioxidants. 2024; 13(4):436. https://doi.org/10.3390/antiox13040436
Chicago/Turabian StyleFan, Ze, Kaibo Ge, Di Wu, Liansheng Wang, Jinnan Li, Chenhui Li, Meng Zhou, Haitao Zhang, Linghong Miao, and Xianping Ge. 2024. "Suitable Cottonseed Protein Concentrate Supplementation in Common Carp (Cyprinus carpio) Serves as an Effective Strategy for Fish Meal Sparing Based on Improvement in Intestinal Antioxidant Capacity, Barrier and Microbiota Composition" Antioxidants 13, no. 4: 436. https://doi.org/10.3390/antiox13040436
APA StyleFan, Z., Ge, K., Wu, D., Wang, L., Li, J., Li, C., Zhou, M., Zhang, H., Miao, L., & Ge, X. (2024). Suitable Cottonseed Protein Concentrate Supplementation in Common Carp (Cyprinus carpio) Serves as an Effective Strategy for Fish Meal Sparing Based on Improvement in Intestinal Antioxidant Capacity, Barrier and Microbiota Composition. Antioxidants, 13(4), 436. https://doi.org/10.3390/antiox13040436