
Ubiquinone (Coenzyme Q-10) Supplementation Influences Exercise-Induced Changes in Serum 25(OH)D3 and the Methyl-Arginine Metabolites: A Double-Blind Randomized Controlled Trial

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
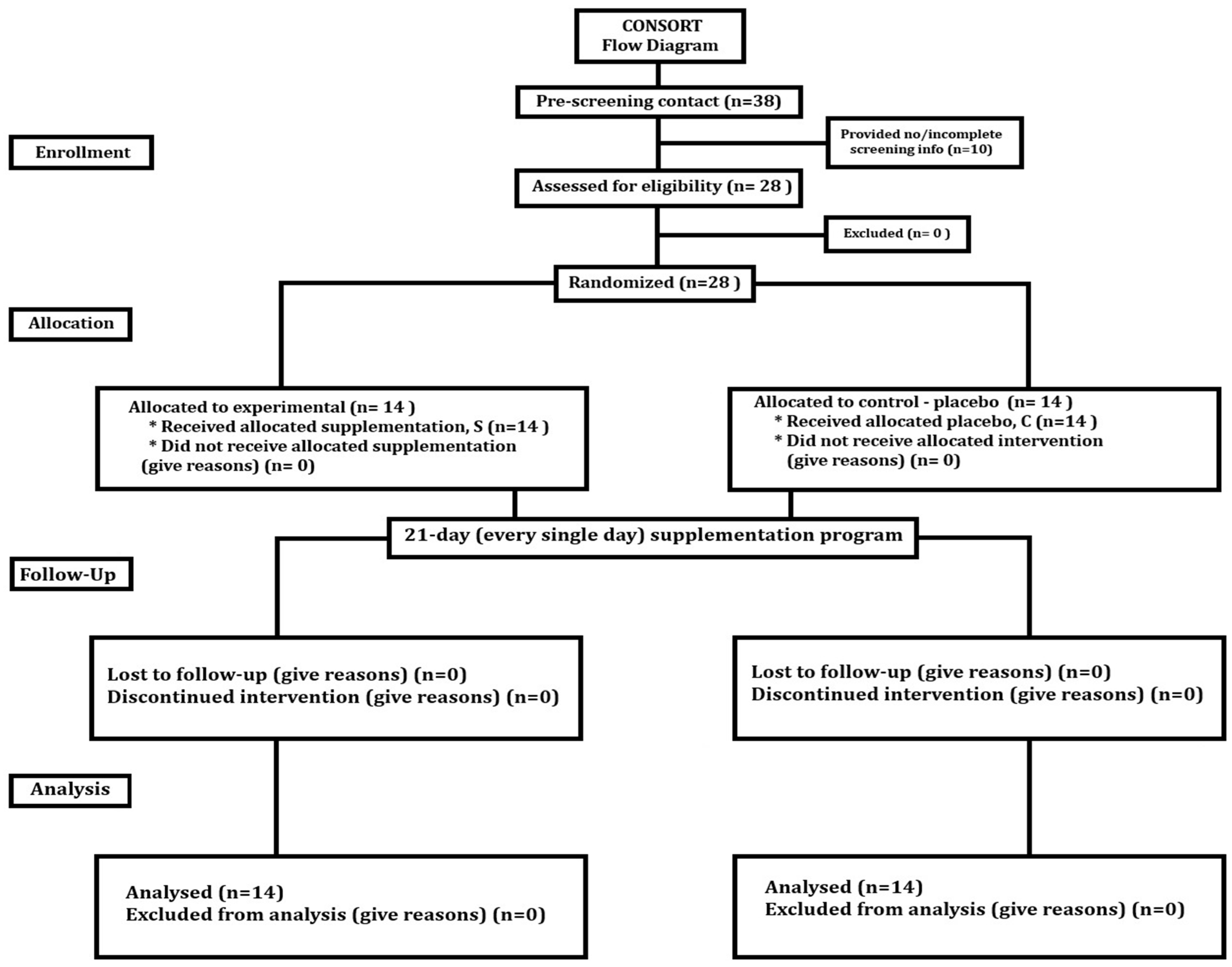
2.2. Experimental Overview
2.3. Participants
2.4. Measurement of Aerobic Components of Fitness: Bruce Treadmill Test
2.5. Supplementation
2.6. Sample Collection and Measurements of CoQ10, Vitamin D, ADMA, Symmetric Dimethylarginine (SDMA), MMA, Dimethylamine (DMA), Arginine (Arg), Citrulline (Cit), and Ornithine (Orn)
2.7. Statistical Analysis
3. Results
3.1. Anthropometric Characteristics and Performance
3.2. Resting Values before and after the 21-Day Supplementation Period
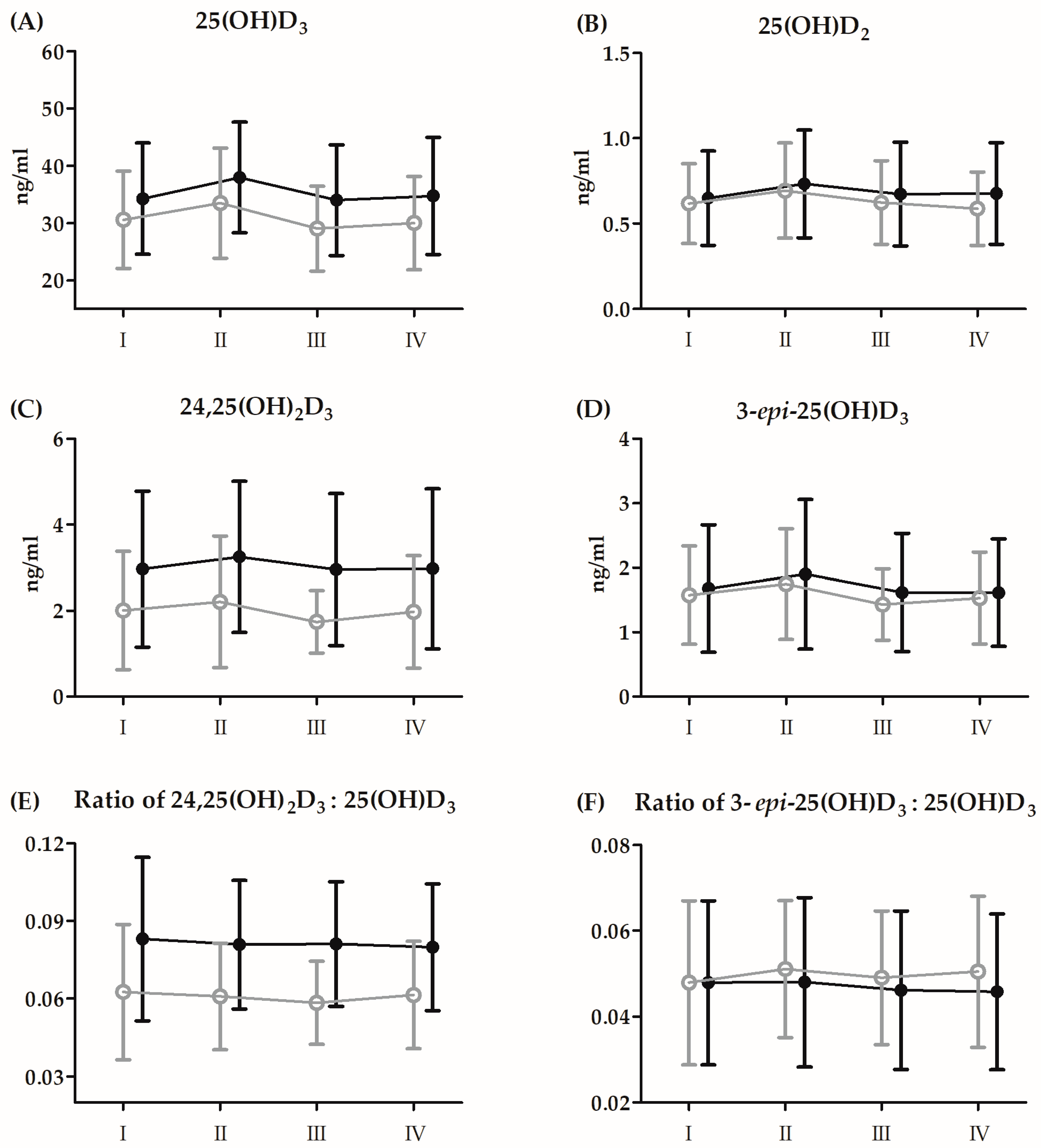
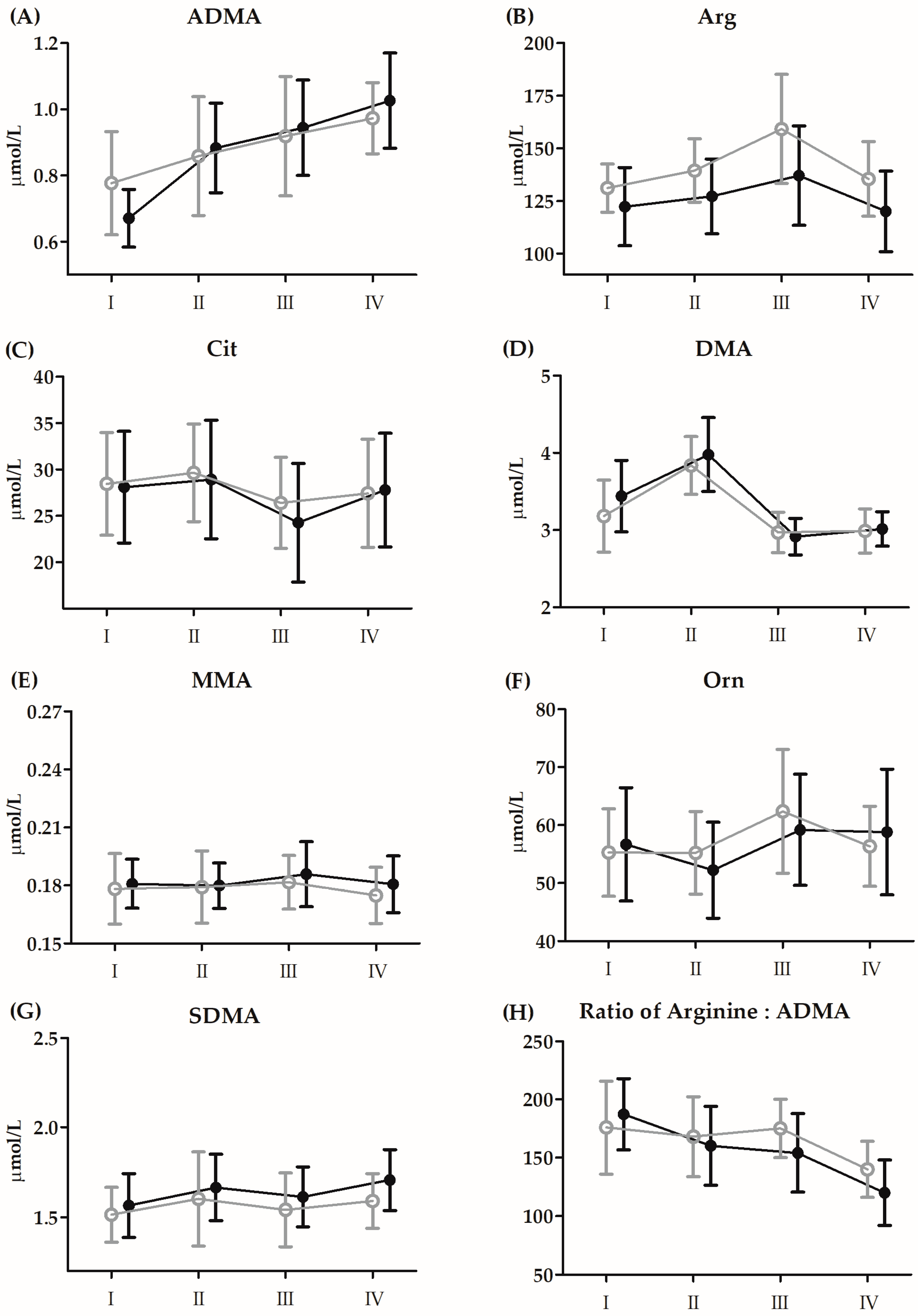
3.3. Changes in Vitamin D and Amino Acid Metabolites before the 21-Day Supplementation Period
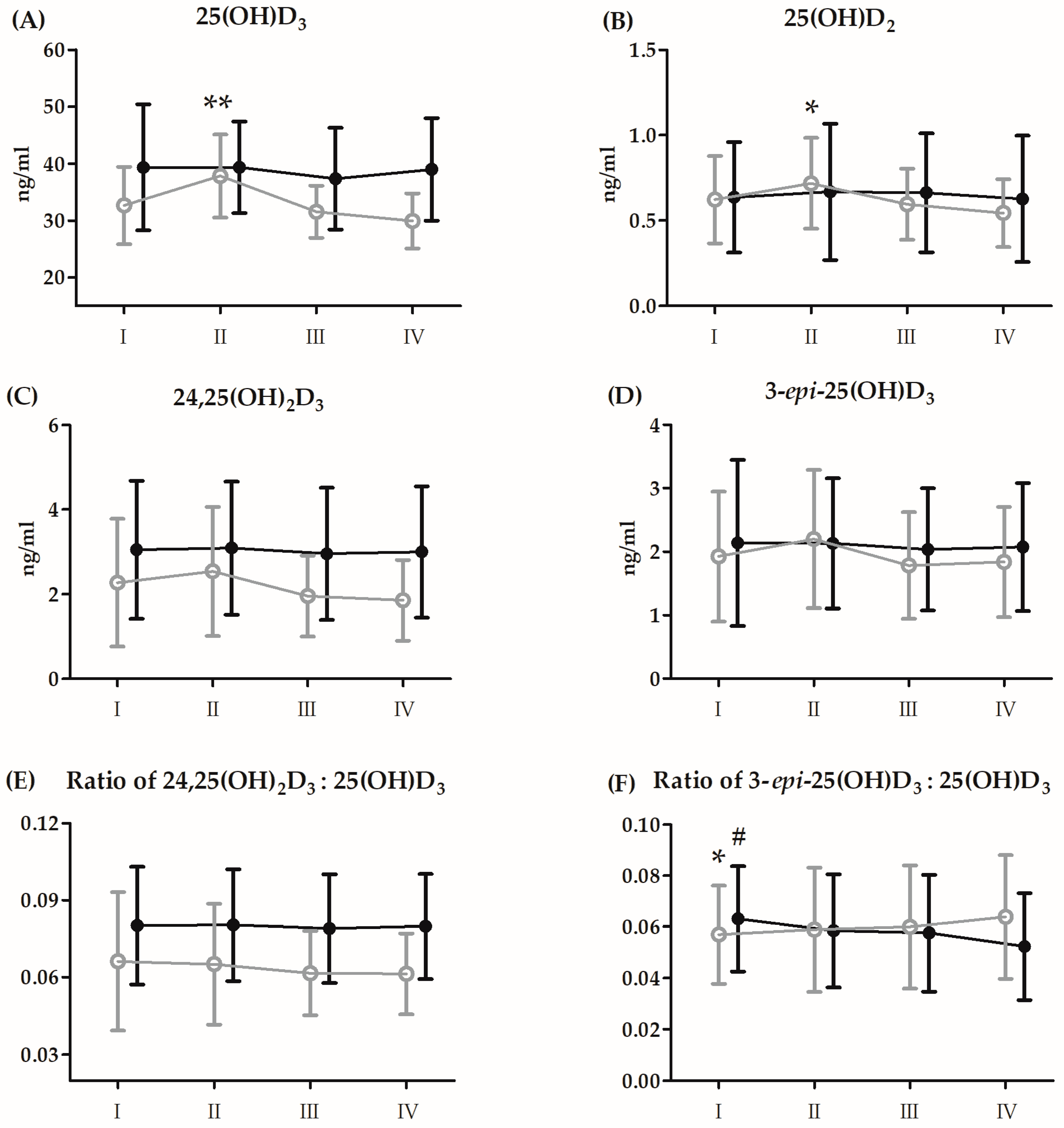
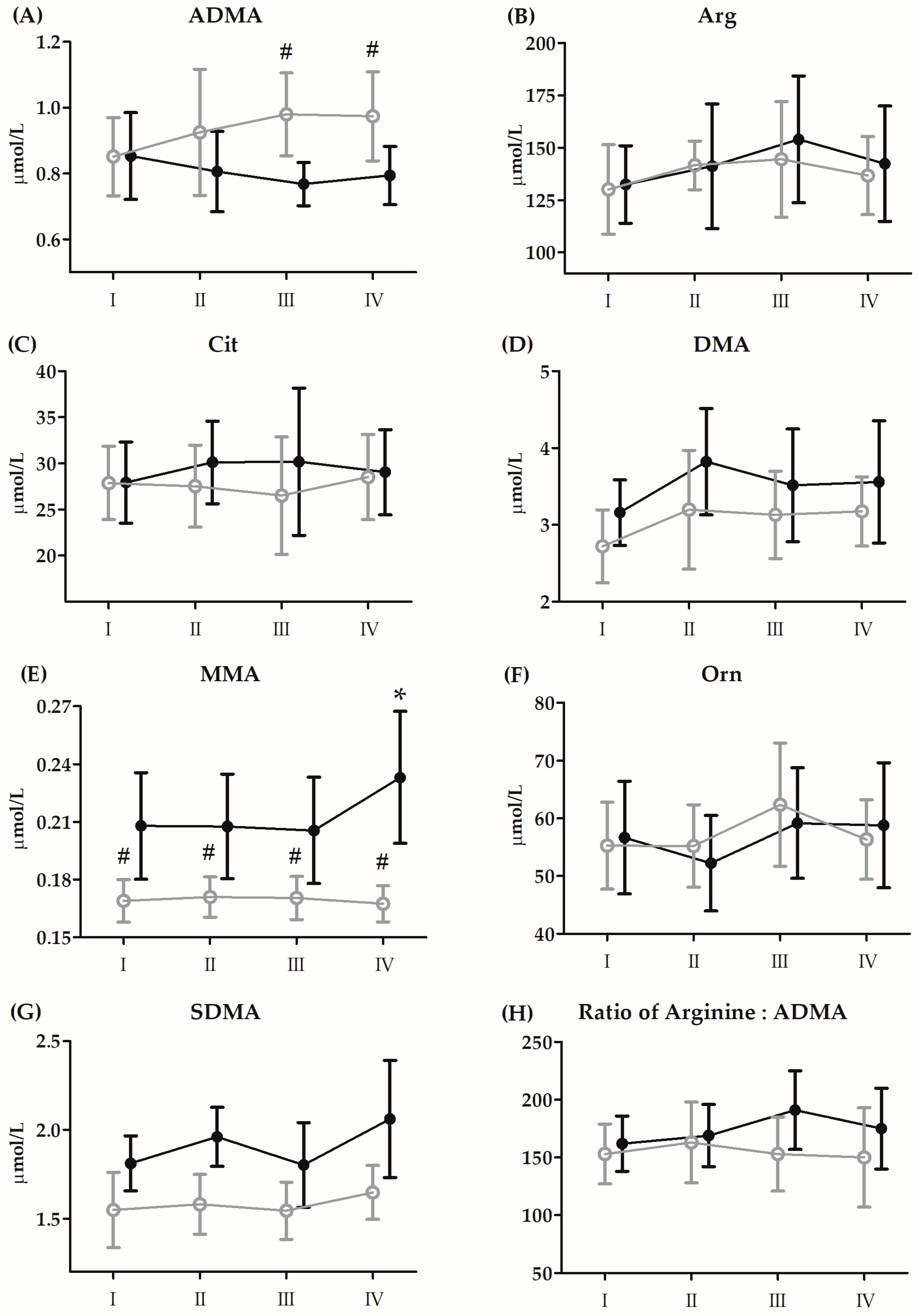
3.4. Changes in Vitamin D and Amino Acid Metabolites after the 21-Day Supplementation Period
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agudelo, L.Z.; Femenia, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal muscle PGC-1alpha1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Belardinelli, R.; Mucaj, A.; Lacalaprice, F.; Solenghi, M.; Seddaiu, G.; Principi, F.; Tiano, L.; Littarru, G.P. Coenzyme Q10 and exercise training in chronic heart failure. Eur. Heart J. 2006, 27, 2675–2681. [Google Scholar] [CrossRef]
- Orlando, P.; Silvestri, S.; Galeazzi, R.; Antonicelli, R.; Marcheggiani, F.; Cirilli, I.; Bacchetti, T.; Tiano, L. Effect of ubiquinol supplementation on biochemical and oxidative stress indexes after intense exercise in young athletes. Redox Rep. 2018, 23, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Gu, B.; Gu, Y.; Groome, L.J.; Sun, J.; Wang, Y. Activation of vitamin D receptor promotes VEGF and CuZn-SOD expression in endothelial cells. J. Steroid Biochem. Mol. Biol. 2014, 140, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Cadenas, E.; Davies, K.J. Free radicals and exercise: An introduction. Free Radic. Biol. Med. 2008, 44, 123–125. [Google Scholar] [CrossRef]
- Davies, K.J.A. Cardiovascular Adaptive Homeostasis in Exercise. Front. Physiol. 2018, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Palm, F.; Onozato, M.L.; Luo, Z.; Wilcox, C.S. Dimethylarginine dimethylaminohydrolase (DDAH): Expression, regulation, and function in the cardiovascular and renal systems. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3227–H3245. [Google Scholar] [CrossRef] [PubMed]
- Poderoso, J.J.; Helfenberger, K.; Poderoso, C. The effect of nitric oxide on mitochondrial respiration. Nitric Oxide 2019, 88, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Chun, R.F.; Liu, P.T.; Modlin, R.L.; Adams, J.S.; Hewison, M. Impact of vitamin D on immune function: Lessons learned from genome-wide analysis. Front. Physiol. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Parsanathan, R.; Levine, S.N.; Bocchini, J.A.; Holick, M.F.; Vanchiere, J.A. The potential link between inherited G6PD deficiency, oxidative stress, and vitamin D deficiency and the racial inequities in mortality associated with COVID-19. Free Radic. Biol. Med. 2020, 161, 84–91. [Google Scholar] [CrossRef]
- Parsanathan, R.; Jain, S.K. Glutathione deficiency alters the vitamin D-metabolizing enzymes CYP27B1 and CYP24A1 in human renal proximal tubule epithelial cells and kidney of HFD-fed mice. Free Radic. Biol. Med. 2019, 131, 376–381. [Google Scholar] [CrossRef]
- Nyborg, C.; Bonnevie-Svendsen, M.; Melsom, H.S.; Melau, J.; Seljeflot, I.; Hisdal, J. Reduced L-Arginine and L-Arginine-ADMA-Ratio, and Increased SDMA after Norseman Xtreme Triathlon. Sports 2021, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Rola, R.; Kowalski, K.; Bienkowski, T.; Studzinska, S. Improved sample preparation method for fast LC-MS/MS analysis of vitamin D metabolites in serum. J. Pharm. Biomed. Anal. 2020, 190, 113529. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewicz, D.; Eickelberg, O. From arginine methylation to ADMA: A novel mechanism with therapeutic potential in chronic lung diseases. BMC Pulm. Med. 2009, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; McClung, H.L.; McClung, J.P.; Urso, M.L.; Pikosky, M.A.; Cloutier, G.J.; Fielding, R.A.; Young, A.J. Molecular responses to moderate endurance exercise in skeletal muscle. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhu, A.; Gutman, J.; Fu, Q.; Liu, S. Cysteine-Rich Whey Protein Isolate (CR-WPI) Ameliorates Erectile Dysfunction by Diminishing Oxidative Stress via DDAH/ADMA/NOS Pathway. Oxid. Med. Cell. Longev. 2022, 2022, 8151917. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Toxic Dimethylarginines: Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA). Toxins 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- MacAllister, R.J.; Rambausek, M.H.; Vallance, P.; Williams, D.; Hoffmann, K.H.; Ritz, E. Concentration of dimethyl-L-arginine in the plasma of patients with end-stage renal failure. Nephrol. Dial. Transpl. 1996, 11, 2449–2452. [Google Scholar] [CrossRef] [PubMed]
- Surdacki, A.; Nowicki, M.; Sandmann, J.; Tsikas, D.; Boeger, R.H.; Bode-Boeger, S.M.; Kruszelnicka-Kwiatkowska, O.; Kokot, F.; Dubiel, J.S.; Froelich, J.C. Reduced urinary excretion of nitric oxide metabolites and increased plasma levels of asymmetric dimethylarginine in men with essential hypertension. J. Cardiovasc. Pharmacol. 1999, 33, 652–658. [Google Scholar] [CrossRef]
- Dzik, K.; Skrobot, W.; Flis, D.J.; Karnia, M.; Libionka, W.; Kloc, W.; Kaczor, J.J. Vitamin D supplementation attenuates oxidative stress in paraspinal skeletal muscles in patients with low back pain. Eur. J. Appl. Physiol. 2018, 118, 143–151. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Mieszkowski, J.; Stankiewicz, B.; Kochanowicz, A.; Niespodzinski, B.; Kowalik, T.; Zmijewski, M.; Kowalski, K.; Rola, R.; Bienkowski, T.; Antosiewicz, J. Ultra-Marathon-Induced Increase in Serum Levels of Vitamin D Metabolites: A Double-Blind Randomized Controlled Trial. Nutrients 2020, 12, 3629. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Z.B.; Taniguchi, H.; Tanisawa, K.; Higuchi, M. Effect of an Acute Bout of Endurance Exercise on Serum 25(OH)D Concentrations in Young Adults. J. Clin. Endocrinol. Metab. 2017, 102, 3937–3944. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Treadmill Speed | Treadmill Incline |
|---|---|---|
| 1 | 2.74 km/h (1.7 mph) | 10% grade |
| 2 | 4.03 km/h (2.5 mph) | 12% grade |
| 3 | 5.47 km/h (3.4 mph) | 14% grade |
| 4 | 6.76 km/h (4.2 mph) | 16% grade |
| 5 | 8.05 km/h (5.0 mph) | 18% grade |
| 6 | 8.86 km/h (5.5 mph) | 20% grade |
| 7 | 9.66 km/h (6.0 mph) | 22% grade |
| 8 | 10.46 km/h (6.5 mph) | 24% grade |
| 9 | 11.26 km/h (7.0 mph) | 26% grade |
| 10 | 12.07 km/h (7.5 mph) | 28% grade |
| Variable | Unit | Supplementation Group (n = 14) | Placebo Group (n = 14) | ||
|---|---|---|---|---|---|
| Before | After | Before | After | ||
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | ||
| Height | cm | 179.1 ± 7.3 | - | 181.2 ± 8.1 | - |
| Body mass | kg | 77.7 ± 7.1 | 76.0 ± 6.8 | 79.9 ± 7.8 | 79.4 ± 7.3 |
| Body fat percentage | % | 12.0 ± 4.6 | 11.5 ± 4.3 | 12.8 ± 5.1 | 12.4 ± 5.0 |
| Body mass index | kg/m2 | 23.5 ± 3.2 | 23.1 ± 3.0 | 24.0 ± 3.6 | 23.7 ± 3.6 |
| Maximal oxygen uptake | mL/min/kg | 57.2 ± 8.0 | 55.9 ± 7.9 | 51.3 ± 9.4 | 52.8 ± 7.5 |
| Respiratory exchange ratio | 1.26 ± 0.05 | 1.28 ± 0.07 | 1.29 ± 0.07 | 1.28 ± 0.06 | |
| Maximal heart rate | beats/min | 197.4 ± 7.5 | 197.7 ± 6.3 | 193.9 ± 5.2 | 192 ± 6.7 |
| Variable | Unit | Supplementation Group (n = 14) | Placebo Group (n = 14) | ||
|---|---|---|---|---|---|
| Before | After | Before | After | ||
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | ||
| CoQ10 | ng/mL | 900 ± 321 | 2486 ± 1180 *† | 849 ± 187 | 827 ± 166 |
| 25(OH)D3 | ng/mL | 34.0 ± 9.0 | 39.5 ± 10.8 * | 30.5 ± 8.0 | 32.3 ± 7.0 |
| 25(OH)D2 | ng/mL | 0.6 ± 0.3 | 0.7 ± 0.3 | 0.6 ± 0.2 | 0.6 ± 0.3 |
| 24,25(OH)2D3 | ng/mL | 3.0 ± 1.8 | 3.3 ± 2.3 | 2.1 ± 1.4 | 2.3 ± 1.5 |
| 3-epi-25(OH)D3 | ng/mL | 1.7 ± 1.0 | 2.6 ± 1.4 * | 1.6 ± 0.8 | 1.9 ± 0.8 |
| Ratio 24,25(OH)2D3: 25(OH)D3 | 0.08 ± 0.03 | 0.08 ± 0.02 | 0.06 ± 0.02 | 0.06 ± 0.03 | |
| Ratio 3-epi-25(OH)D3: 25(OH)D3 | 0.04 ± 0.02 | 0.06 ± 0.02 * | 0.05 ± 0.02 | 0.06 ± 0.02 | |
| ADMA | µmol/L | 0.65 ± 0.08 | 0.85 ± 0.13 | 0.77 ± 0.15 | 0.85 ± 0.11 |
| Arg | µmol/L | 122.3 ± 18.5 | 136.9 ± 19.8 * | 131.2 ± 11.5 | 129.2 ± 20.9 |
| Cit | µmol/L | 28.4 ± 5.5 | 29.9 ± 4.2 | 28.4 ± 5.5 | 27.6 ± 3.9 |
| DMA | µmol/L | 3.4 ± 0.5 | 3.2 ± 0.4 | 3.2 ± 0.5 | 2.7 ± 0.5 |
| MMA | µmol/L | 0.17 ± 0.01 | 0.21 ± 0.03 *† | 0.17 ± 0.02 | 0.17 ± 0.02 |
| Orn | µmol/L | 60.0 ± 11.0 | 63.6 ± 11.1 | 55.3 ± 4.6 | 56.7 ± 8.5 |
| SDMA | µmol/L | 1.6 ± 0.2 | 1.8 ± 0.2 *† | 1.5 ± 0.2 | 1.5 ± 0.3 |
| Ratio Arg:ADMA | 183 ± 30.7 | 162.6 ± 24.8 | 175.9 ± 40.0 | 156.3 ± 26.8 | |
| Variable | Effect | F | Df | P | Effect Size (η2) | Post-Hoc Outcome |
|---|---|---|---|---|---|---|
| 25(OH)D3 | GR | 1.87 | 1, 26 | 0.18 | 0.06 | II > I, III, IV |
| RM | 11.78 | 3, 78 | <0.01 ** | 0.31 | ||
| GR × RM | 0.27 | 3, 78 | 0.84 | 0.01 | ||
| 25(OH)D2 | GR | 0.26 | 1, 26 | 0.61 | 0.01 | II > I, III, IV |
| RM | 14.57 | 3, 78 | <0.01 ** | 0.35 | ||
| GR × RM | 1.63 | 3, 78 | 0.18 | 0.05 | ||
| 24,25(OH)2D3 | GR | 3.32 | 1, 26 | 0.08 | 0.11 | II > III |
| RM | 3.49 | 3, 78 | 0.02 * | 0.12 | ||
| GR × RM | 0.43 | 3, 78 | 0.73 | 0.01 | ||
| 3-epi-25(OH)D3 | GR | 0.16 | 1, 26 | 0.68 | 0.01 | II > I, III, IV |
| RM | 9.27 | 3, 78 | <0.01 ** | 0.26 | ||
| GR × RM | 0.29 | 3, 78 | 0.83 | 0.01 | ||
| 24,25(OH)2D3:25(OH)D3 | GR | 5.56 | 1, 26 | 0.03 * | 0.17 | S > C |
| RM | 0.86 | 3, 78 | 0.46 | 0.03 | ||
| GR × RM | 0.41 | 3, 78 | 0.74 | 0.02 | ||
| 3-epi-25(OH)D3:25(OH)D3 | GR | 0.29 | 1, 26 | 0.59 | 0.01 | I, II, IV > III |
| RM | 4.31 | 3, 78 | <0.01 ** | 0.14 | ||
| GR × RM | 0.62 | 3, 78 | 0.60 | 0.02 |
| Variable | Effect | F | Df | P | Effect Size (η2) | Post-Hoc Outcome |
|---|---|---|---|---|---|---|
| ADMA | GR | 0.01 | 1, 26 | 0.98 | <0.01 | I < II, III, IV |
| RM | 25.88 | 3, 78 | <0.01 ** | 0.43 | ||
| GR × RM | 0.24 | 3, 78 | 0.07 | 0.06 | ||
| Arg | GR | 3.49 | 1, 26 | 0.07 | 0.11 | III > I, II, IV |
| RM | 15.60 | 3, 78 | <0.01 ** | 0.37 | ||
| GR × RM | 0.63 | 3, 78 | 0.59 | 0.02 | ||
| Cit | GR | 0.12 | 1, 26 | 0.72 | <0.01 | III > I, II, IV |
| RM | 10.81 | 3, 78 | <0.01 ** | 0.29 | ||
| GR × RM | 1.05 | 3, 78 | 0.37 | 0.04 | ||
| DMA | GR | 0.24 | 1, 26 | 0.42 | 0.02 | II > I > II, IV |
| RM | 99.03 | 3, 78 | <0.01 ** | 0.79 | ||
| GR × RM | 2.36 | 3, 78 | 0.07 | 0.08 | ||
| MMA | GR | 0.49 | 1, 26 | 0.48 | 0.01 | |
| RM | 1.71 | 3, 78 | 0.17 | 0.06 | ||
| GR × RM | 0.30 | 3, 78 | 0.82 | 0.01 | ||
| Orn | GR | 0.06 | 1, 26 | 0.80 | <0.01 | III > I, II |
| RM | 6.14 | 3, 78 | <0.01 ** | 0.15 | ||
| GR × RM | 1.50 | 3, 78 | 0.21 | 0.04 | ||
| SDMA | GR | 2.61 | 1, 26 | 0.11 | 0.09 | I < IV |
| RM | 2.69 | 3, 78 | 0.05 * | 0.09 | ||
| GR × RM | 0.21 | 3, 78 | 0.88 | 0.01 | ||
| Arg:ADMA | GR | 1.23 | 1, 26 | 0.27 | 0.05 | I, II, III > IV |
| RM | 19.64 | 3, 78 | <0.01 ** | 0.43 | ||
| GR × RM | 2.53 | 3, 78 | 0.07 | 0.08 |
| Variable | Effect | F | Df | P | Effect Size (η2) | Post-Hoc Outcome |
|---|---|---|---|---|---|---|
| 25(OH)D3 | GR | 4.56 | 1, 26 | 0.04 * | 0.14 | S > P |
| RM | 8.67 | 3, 78 | <0.01 ** | 0.25 | II > I, III | |
| GR × RM | 5.25 | 3, 78 | <0.01 ** | 0.16 | PII > PI, PIII, PIV | |
| 25(OH)D2 | GR | 0.75 | 1, 26 | 0.39 | 0.03 | |
| RM | 5.03 | 3, 78 | <0.01 ** | 0.16 | II > I, III, IV | |
| GR × RM | 3.00 | 3, 78 | 0.03 * | 0.11 | PII > PIV | |
| 24,25(OH)2D3 | GR | 4.96 | 1, 26 | 0.03 * | 0.16 | S > P |
| RM | 2.97 | 3, 78 | 0.04 * | 0.11 | II > III | |
| GR × RM | 1.28 | 3, 78 | 0.28 | 0.04 | ||
| 3-epi-25(OH)D3 | GR | 0.69 | 1, 26 | 0.41 | 0.02 | |
| RM | 4.13 | 3, 78 | <0.01 ** | 0.13 | II > III, IV | |
| GR × RM | 1.82 | 3, 78 | 0.15 | 0.06 | ||
| 24,25(OH)2D3:25(OH)D3 | GR | 5.05 | 1, 26 | 0.03 * | 0.16 | S > P |
| RM | 0.72 | 3, 78 | 0.53 | 0.02 | ||
| GR × RM | 0.34 | 3, 78 | 0.79 | 0.01 | ||
| 3-epi-25(OH)D3:25(OH)D3 | GR | 0.01 | 1, 26 | 0.97 | <0.01 | |
| RM | 0.46 | 3, 78 | 0.70 | 0.02 | ||
| GR × RM | 3.90 | 3, 78 | 0.01 * | 0.12 | SI > SIV; PI < PIV |
| Variable | Effect | F | Df | P | Effect Size (η2) | Post-Hoc Outcome |
|---|---|---|---|---|---|---|
| ADMA | GR | 17.59 | 1, 26 | <0.01 ** | 0.4 | S < P |
| RM | 0.39 | 3, 78 | 75 | 0.01 | ||
| GR × RM | 4.82 | 3, 78 | <0.01 ** | 0.15 | SIII, SIV < PIII, PIV | |
| Arg | GR | 0.31 | 1, 26 | 0.58 | 0.01 | I < III |
| RM | 3.51 | 3, 78 | 0.02 * | 0.11 | ||
| GR × RM | 1.63 | 3, 78 | 0.18 | 0.05 | ||
| Cit | GR | 2.79 | 1, 26 | 0.10 | 0.09 | |
| RM | 2.08 | 3, 78 | 0.10 | 0.07 | ||
| GR × RM | 0.11 | 3, 78 | 0.95 | <0.01 | ||
| DMA | GR | 4.78 | 1, 26 | 0.42 | 0.15 | S > P |
| RM | 8.66 | 3, 78 | <0.01 ** | 0.24 | I < II, III, IV | |
| GR × RM | 0.46 | 3, 78 | 0.07 | 0.01 | ||
| MMA | GR | 24.18 | 1, 26 | <0.01 ** | 0.48 | S > P |
| RM | 15.39 | 3, 78 | <0.01 ** | 0.37 | I, II, III < IV | |
| GR × RM | 24.51 | 3, 78 | <0.01 ** | 0.48 | SI–SIV > PI–PIV | |
| Orn | GR | 0.82 | 1, 26 | 0.37 | 0.03 | II < III |
| RM | 3.84 | 3, 78 | <0.01 ** | 0.13 | ||
| GR × RM | 0.83 | 3, 78 | 0.47 | 0.03 | ||
| SDMA | GR | 29.54 | 1, 26 | <0.01 ** | 0.53 | S > P |
| RM | 6.58 | 3, 78 | <0.01 ** | 0.2 | I < IV | |
| GR × RM | 1.45 | 3, 78 | 0.23 | 0.05 | ||
| Arg/ADMA | GR | 5.63 | 1, 26 | 0.03 * | 0.18 | S > P |
| RM | 1.09 | 3, 78 | 0.35 | 0.04 | ||
| GR × RM | 1.54 | 3, 78 | 0.20 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mieszkowski, J.; Kochanowicz, A.; Brzezińska, P.; Kochanowicz, M.; Żołądkiewicz, K.; Stankiewicz, B.; Niespodziński, B.; Reczkowicz, J.; Kowalski, K.; Antosiewicz, J. Ubiquinone (Coenzyme Q-10) Supplementation Influences Exercise-Induced Changes in Serum 25(OH)D3 and the Methyl-Arginine Metabolites: A Double-Blind Randomized Controlled Trial. Antioxidants 2024, 13, 760. https://doi.org/10.3390/antiox13070760
Mieszkowski J, Kochanowicz A, Brzezińska P, Kochanowicz M, Żołądkiewicz K, Stankiewicz B, Niespodziński B, Reczkowicz J, Kowalski K, Antosiewicz J. Ubiquinone (Coenzyme Q-10) Supplementation Influences Exercise-Induced Changes in Serum 25(OH)D3 and the Methyl-Arginine Metabolites: A Double-Blind Randomized Controlled Trial. Antioxidants. 2024; 13(7):760. https://doi.org/10.3390/antiox13070760
Chicago/Turabian StyleMieszkowski, Jan, Andrzej Kochanowicz, Paulina Brzezińska, Magdalena Kochanowicz, Katarzyna Żołądkiewicz, Błażej Stankiewicz, Bartłomiej Niespodziński, Joanna Reczkowicz, Konrad Kowalski, and Jędrzej Antosiewicz. 2024. "Ubiquinone (Coenzyme Q-10) Supplementation Influences Exercise-Induced Changes in Serum 25(OH)D3 and the Methyl-Arginine Metabolites: A Double-Blind Randomized Controlled Trial" Antioxidants 13, no. 7: 760. https://doi.org/10.3390/antiox13070760
APA StyleMieszkowski, J., Kochanowicz, A., Brzezińska, P., Kochanowicz, M., Żołądkiewicz, K., Stankiewicz, B., Niespodziński, B., Reczkowicz, J., Kowalski, K., & Antosiewicz, J. (2024). Ubiquinone (Coenzyme Q-10) Supplementation Influences Exercise-Induced Changes in Serum 25(OH)D3 and the Methyl-Arginine Metabolites: A Double-Blind Randomized Controlled Trial. Antioxidants, 13(7), 760. https://doi.org/10.3390/antiox13070760