
Citrus Pomace as a Source of Plant Complexes to Be Used in the Nutraceutical Field of Intestinal Inflammation

 , , , ,
, , , ,  ,
,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Phytochemical Analyses
2.2.1. Proton Nuclear Magnetic Resonance (1H-NMR) Analysis
2.2.2. Secondary Metabolites Screening by Colorimetric Assays
Total Phenolic Compounds (TPC)
Total Flavonoid Compounds (TFC)
Vanillin Index
Proanthocyanidins
2.2.3. LC-DAD-ESI-MS Analysis
2.3. In Vitro Simulated Gastrointestinal Digestion
2.4. In Vitro Antioxidant and Anti-Inflammatory Assays
2.4.1. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
2.4.2. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
2.4.3. Ferric-Reducing Antioxidant Power (FRAP) Assay
2.4.4. ORAC
2.4.5. β-Carotene Bleaching (BCB) Assay
2.4.6. Iron-Chelating Activity (ICA) Assay
2.4.7. Heat-Induced Bovine Serum Albumin Denaturation (ADA)
2.4.8. Protease-Inhibitory Activity (PIA)
2.5. Antioxidant and Anti-Inflammatory Cell-Based Assays
2.5.1. Cell Culture and Treatments
2.5.2. Cell Viability
2.5.3. Cell Proliferation by IncuCyte
2.5.4. Transepithelial Electric Resistance (TEER) Measurement
2.5.5. Immunofluorescence
2.5.6. Intracellular Reactive Oxygen Species (ROS) Levels
2.5.7. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.6. Statistical Analyses
3. Results
3.1. Standardization and Titration of OE and LE
3.2. Phytochemical Characterization
3.2.1. 1H-NMR Profiling
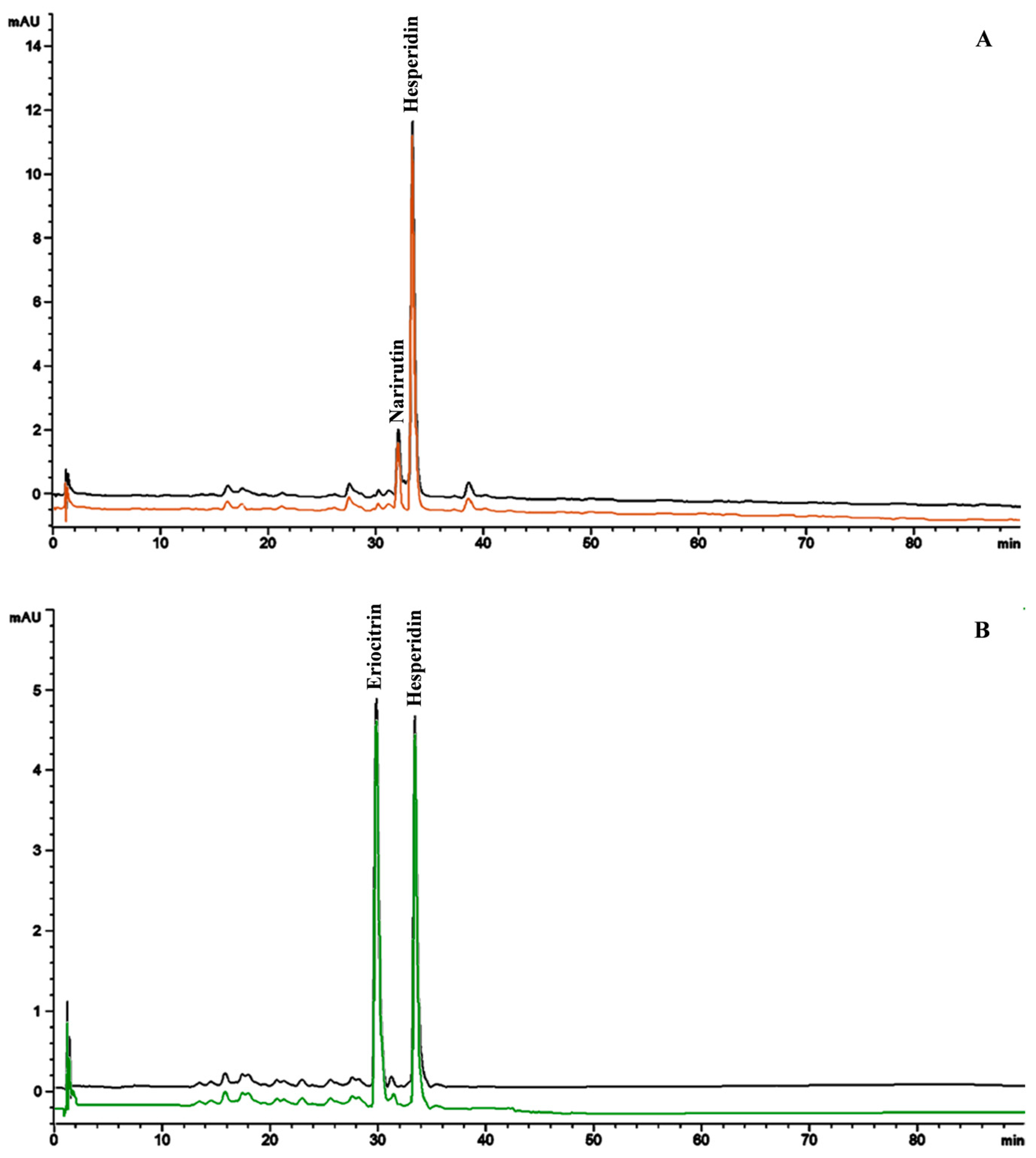
3.2.2. Secondary Metabolites: Phytochemical Screening and LC-DAD-ESI-MS Analysis
3.3. Intestinal Bioaccessibility
3.4. Antioxidant and Anti-Inflammatory Activity
3.4.1. In Vitro Cell-Free Assays
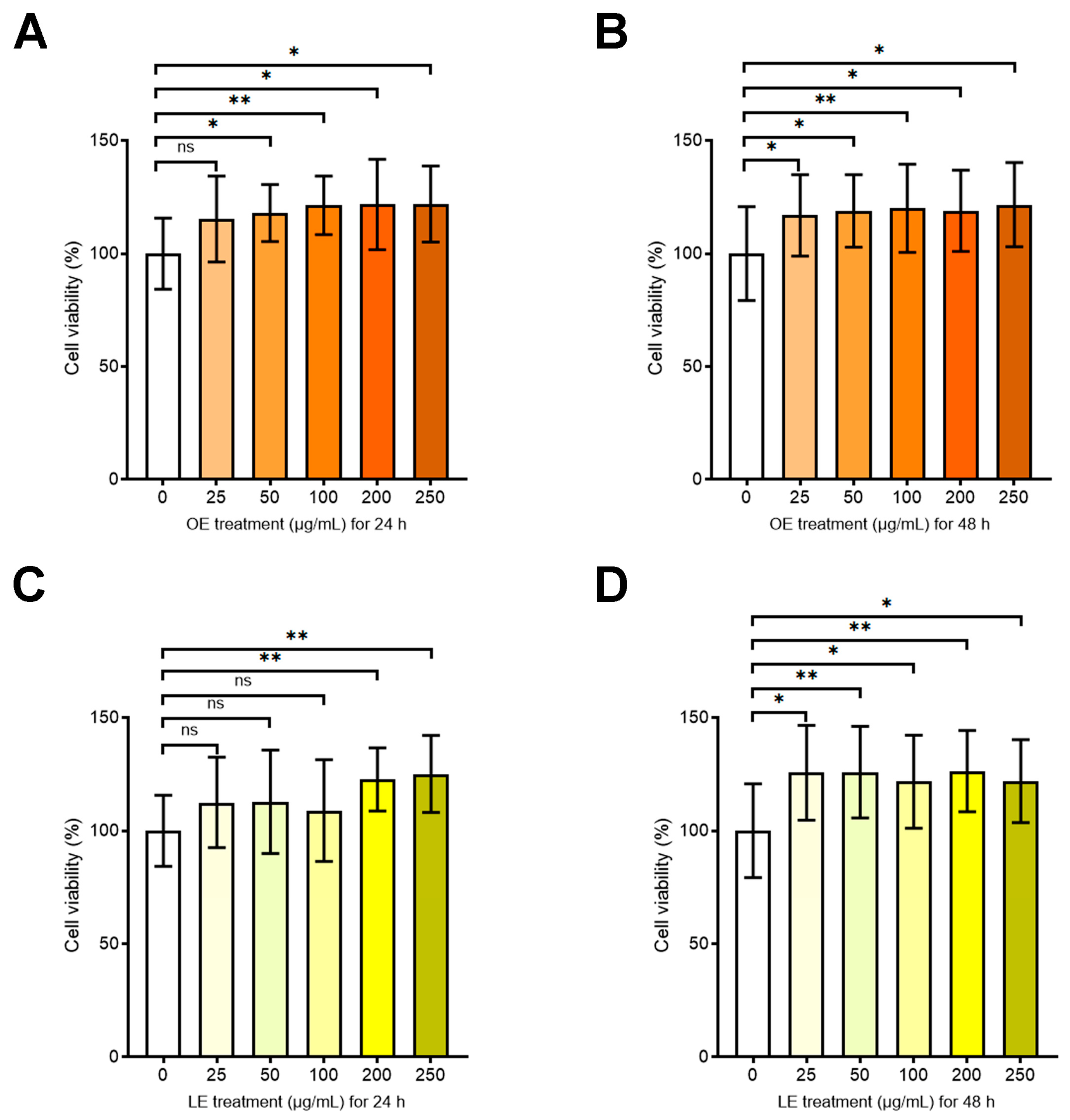
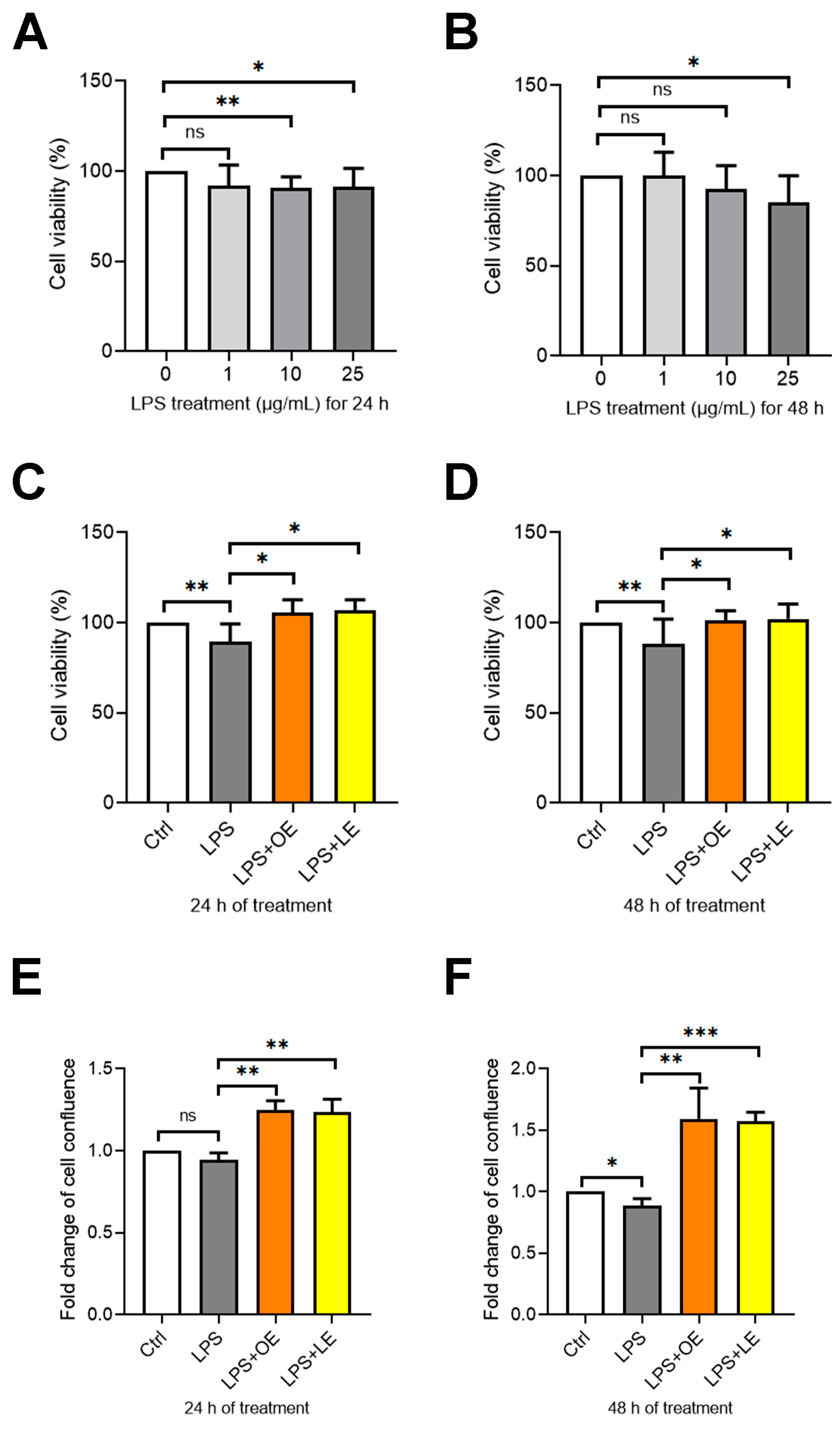
3.4.2. Effects of OE and LE on Cell Viability and Proliferation
3.4.3. Effects of OE and LE on Intestinal Barrier Permeability
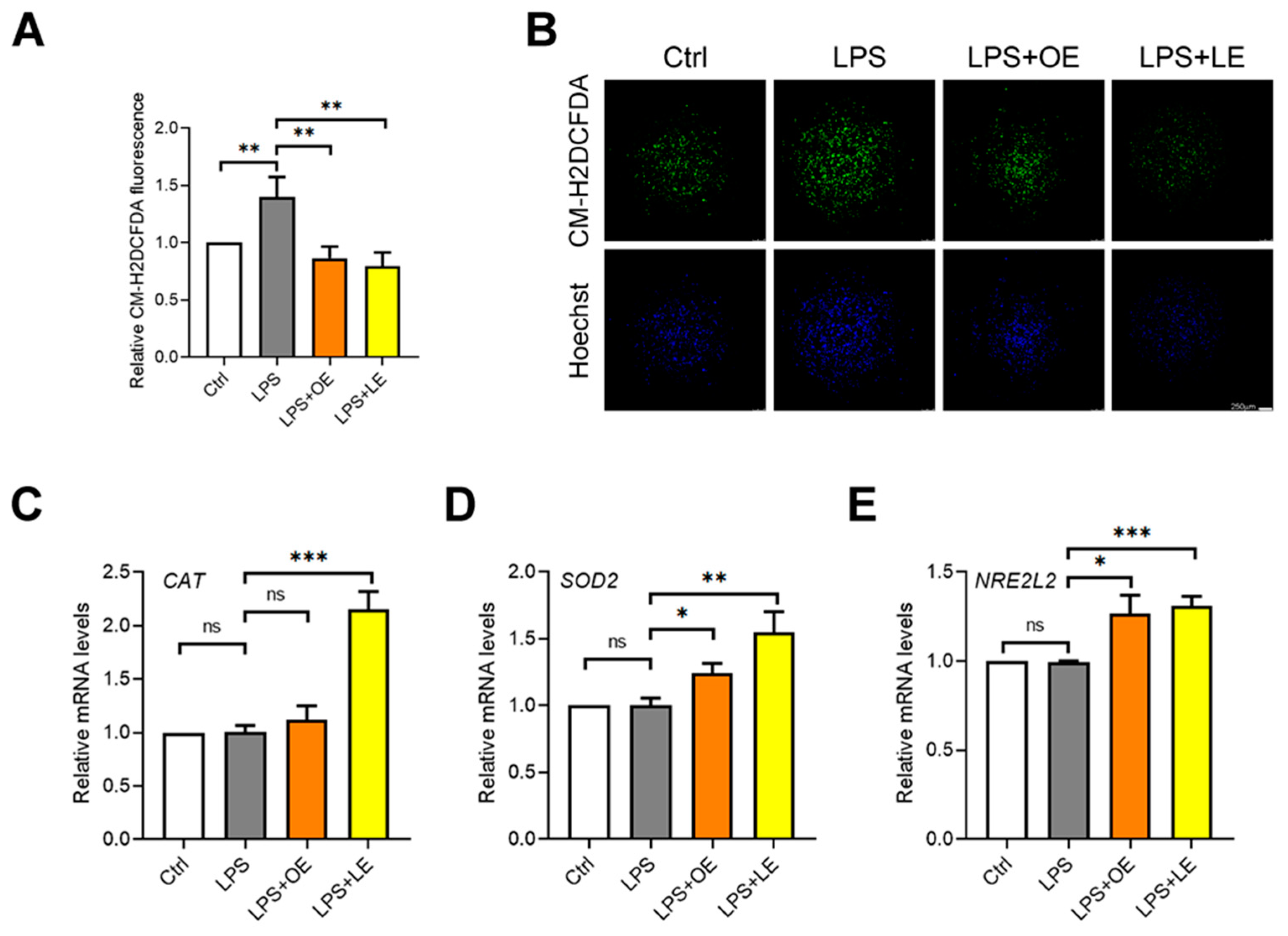
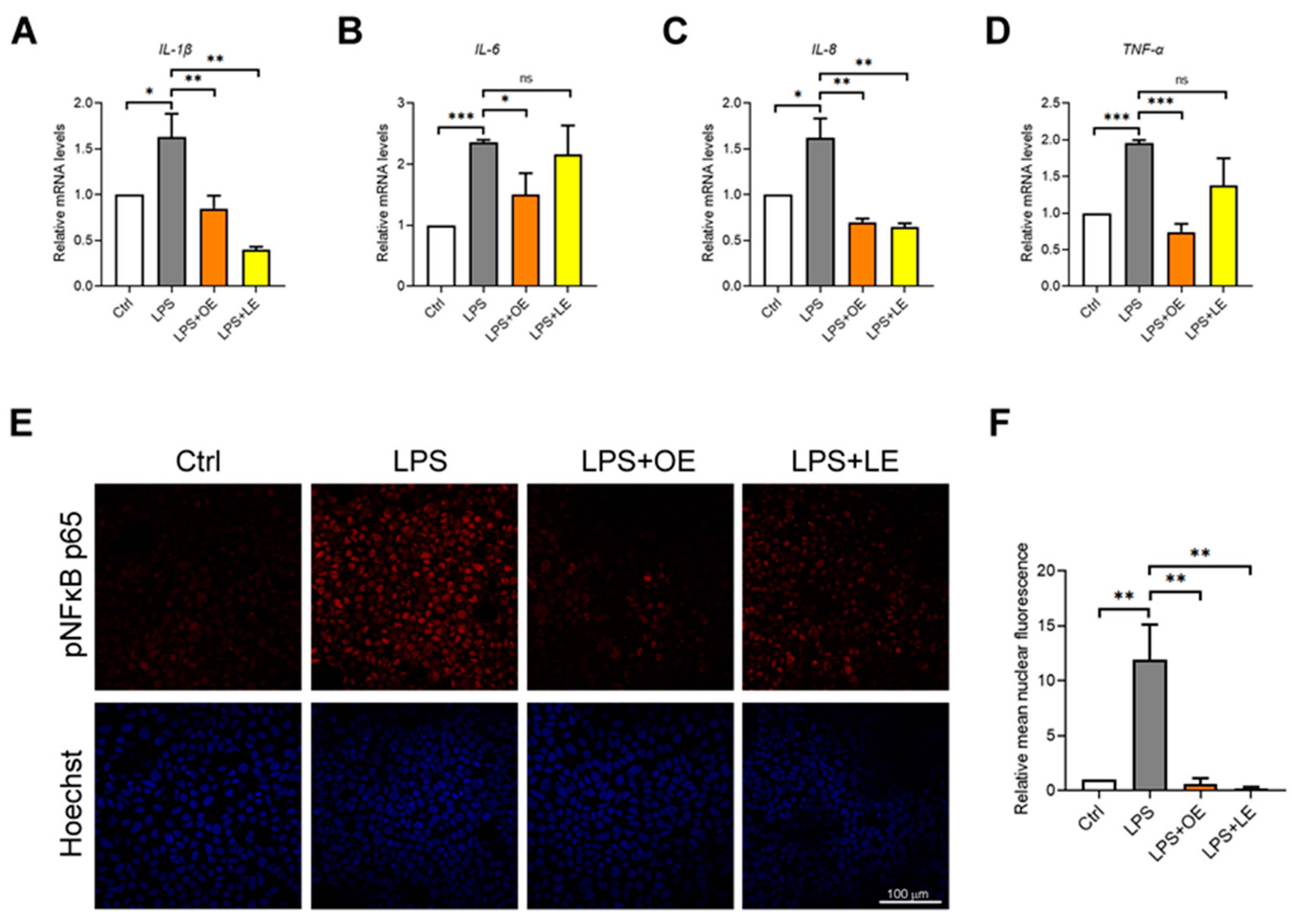
3.4.4. Effects of OE and LE on Oxidative Stress and Inflammatory Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maiuolo, J.; Bosco, F.; Guarnieri, L.; Nucera, S.; Ruga, S.; Oppedisano, F.; Tucci, L.; Muscoli, C.; Palma, E.; Giuffrè, A.M.; et al. Protective Role of an Extract Waste Product from Citrus bergamia in an In Vitro Model of Neurodegeneration. Plants 2023, 12, 2126. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Antioxidant, Anti-Inflammatory and Anti-Angiogenic Properties of Citrus lumia Juice. Front. Pharmacol. 2020, 11, 593506. [Google Scholar] [CrossRef] [PubMed]
- Caggia, C.; Palmeri, R.; Russo, N.; Timpone, R.; Randazzo, C.L.; Todaro, A.; Barbagallo, S. Employ of Citrus By-product as Fat Replacer Ingredient for Bakery Confectionery Products. Front. Nutr. 2020, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Šafranko, S.; Šubarić, D.; Jerković, I.; Jokić, S. Citrus By-Products as a Valuable Source of Biologically Active Compounds with Promising Pharmaceutical, Biological and Biomedical Potential. Pharmaceuticals 2023, 16, 1081. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/markets-and-trade/commodities/citrus/en/ (accessed on 20 June 2024).
- Multari, S.; Licciardello, C.; Caruso, M.; Martens, S. Monitoring the changes in phenolic compounds and carotenoids occurring during fruit development in the tissues of four citrus fruits. Food Res. Int. 2020, 134, 109228. [Google Scholar] [CrossRef]
- Raimondo, M.; Caracciolo, F.; Cembalo, L.; Chinnici, G.; Pecorino, B.; D’Amico, M. Making Virtue Out of Necessity: Managing the Citrus Waste Supply Chain for Bioeconomy Applications. Sustainability 2018, 10, 4821. [Google Scholar] [CrossRef]
- Cirrincione, F.; Ferranti, P.; Ferrara, A.; Romano, A. A critical evaluation on the valorization strategies to reduce and reuse orange waste in bakery industry. Food Res. Int. 2024, 187, 114422. [Google Scholar] [CrossRef]
- Patsalou, M.; Samanides, C.G.; Protopapa, E.; Stavrinou, S.; Vyrides, I.; Koutinas, M. A Citrus Peel Waste Biorefinery for Ethanol and Methane Production. Molecules 2019, 24, 2451. [Google Scholar] [CrossRef]
- Denaro, M.; Smeriglio, A.; Trombetta, D. Antioxidant and Anti-Inflammatory Activity of Citrus Flavanones Mix and Its Stability after In Vitro Simulated Digestion. Antioxidants 2021, 10, 140. [Google Scholar] [CrossRef]
- Smeriglio, A.; Iraci, N.; Denaro, M.; Mandalari, G.; Giofrè, S.V.; Trombetta, D. Synergistic Combination of Citrus Flavanones as Strong Antioxidant and COX-Inhibitor Agent. Antioxidants 2023, 12, 972. [Google Scholar] [CrossRef]
- Mosca, A.; Crudele, A.; Smeriglio, A.; Braghini, M.R.; Panera, N.; Comparcola, D.; Alterio, A.; Sartorelli, M.R.; Tozzi, G.; Raponi, M.; et al. Antioxidant activity of Hydroxytyrosol and Vitamin E reduces systemic inflammation in children with paediatric NAFLD. Dig. Liver Dis. 2021, 53, 1154–1158. [Google Scholar] [CrossRef]
- Egbuna, C.; Awuchi, C.G.; Kushwaha, G.; Rudrapal, M.; Patrick-Iwuanyanwu, K.C.; Singh, O.; Odoh, U.E.; Khan, J.; Jeevanandam, J.; Kumarasamy, S.; et al. Bioactive Compounds Effective Against Type 2 Diabetes Mellitus: A Systematic Review. Curr. Top. Med. Chem. 2021, 21, 1067–1095. [Google Scholar] [CrossRef] [PubMed]
- Institute for Health Metrics and Evaluation. Available online: https://www.healthdata.org/research-analysis/library/global-regional-and-national-burden-inflammatory-bowel-disease-195 (accessed on 22 June 2024).
- Smeriglio, A.; Marcoccia, D.; Denaro, M.; Trombetta, D. Nutraceuticals in the Treatment of Inflammatory Bowel Disease: How the Panorama has Changed in the Last Decade? Curr. Med. Chem. 2023, 30, 2165–2190. [Google Scholar] [CrossRef]
- Seyedian, S.S.; Nokhostin, F.; Malamir, M.D. A review of the diagnosis, prevention, and treatment methods of inflammatory bowel disease. J. Med. Life 2019, 12, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Chen, D.; Zhong, R.; Peng, X. Animal models of inflammatory bowel disease: Category and evaluation indexes. Gastroenterol. Rep. 2024, 12, goae021. [Google Scholar] [CrossRef]
- Joshi, A.; Soni, A.; Acharya, S. In vitro models and ex vivo systems used in inflammatory bowel disease. Vitr. Model. 2022, 1, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.C. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The epidemiology of inflammatory bowel disease: East meets west. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef]
- Kobayashi, T.; Hibi, T. Improving IBD outcomes in the era of many treatment options. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 195. [Google Scholar] [CrossRef]
- Smeriglio, A.; Cornara, L.; Denaro, M.; Barreca, D.; Burlando, B.; Xiao, J.; Trombetta, D. Antioxidant and cytoprotective activities of an ancient Mediterranean citrus (Citrus lumia Risso) albedo extract: Microscopic observations and polyphenol characterization. Food Chem. 2019, 279, 347–355. [Google Scholar] [CrossRef]
- Sanna, C.; Chiocchio, I.; Mandrone, M.; Bonvicini, F.; Gentilomi, G.A.; Trincia, S.; Poli, F. Metabolomic analysis and bioactivities of Arbutus unedo leaves harvested across the seasons in different natural habitats of Sardinia (Italy). BMC Plant Biol. 2023, 23, 490. [Google Scholar] [CrossRef] [PubMed]
- Mandrone, M.; Chiocchio, I.; Barbanti, L.; Tomasi, P.; Tacchini, M.; Poli, F. Metabolomic study of sorghum (Sorghum bicolor) to interpret plant behavior under variable field conditions in view of smart agriculture applications. J. Agric. Food Chem. 2021, 69, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Ingegneri, M.; Smeriglio, A.; Rando, R.; Gervasi, T.; Tamburello, M.P.; Ginestra, G.; La Camera, E.; Pennisi, R.; Sciortino, M.T.; Mandalari, G.; et al. Composition and Biological Properties of Blanched Skin and Blanch Water Belonging to Three Sicilian Almond Cultivars. Nutrients 2023, 15, 1545. [Google Scholar] [CrossRef] [PubMed]
- Lenucci, M.S.; Cadinu, D.; Taurino, M.; Piro, G.; Dalessandro, G. Antioxidant composition in cherry and high-pigment tomato cultivars. J. Agric. Food Chem. 2006, 54, 2606–2613. [Google Scholar] [CrossRef]
- Smeriglio, A.; Ingegneri, M.; Germanò, M.P.; Miori, L.; Battistini, G.; Betuzzi, F.; Malaspina, P.; Trombetta, D.; Cornara, L. Pharmacognostic Evaluation of Monarda didyma L. Growing in Trentino (Northern Italy) for Cosmeceutical Applications. Plants 2024, 13, 112. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.L.; Rudaz, S.; Hae Choi, Y.; Kyong Kim, H. Plant metabolomics: From holistic data to relevant biomarkers. Curr. Med. Chem. 2013, 20, 1056–1090. [Google Scholar] [PubMed]
- Zenodo Data Repository. Available online: https://zenodo.org/records/12522369 (accessed on 24 June 2024).
- Wu, X.X.; Huang, X.L.; Chen, R.R.; Li, T.; Ye, H.J.; Xie, W.; Huang, Z.M.; Cao, G.Z. Paeoniflorin Prevents Intestinal Barrier Disruption and Inhibits Lipopolysaccharide (LPS)-Induced Inflammation in Caco-2 Cell Monolayers. Inflammation 2019, 42, 2215–2225. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; von der Weid, P.Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes 2020, 11, 421–432. [Google Scholar] [CrossRef]
- Leporini, M.; Loizzo, M.R.; Sicari, V.; Pellicanò, T.M.; Reitano, A.; Dugay, A.; Deguin, B.; Tundis, R. Citrus x Clementina Hort. Juice Enriched with Its By-Products (Peels and Leaves): Chemical Composition, In Vitro Bioactivity, and Impact of Processing. Antioxidants 2020, 9, 298. [Google Scholar] [CrossRef]
- Zema, D.A.; Calabrò, P.S.; Folino, A.; Tamburino, V.; Zappia, G.; Zimbone, S.M. Valorisation of Citrus Processing Waste: A Review. Waste Manag. 2018, 80, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Putnik, P.; Bursać Kovačević, D.; Režek Jambrak, A.; Barba, F.J.; Cravotto, G.; Binello, A.; Lorenzo, J.M.; Shpigelman, A. Innovative “Green” and Novel Strategies for the Extraction of Bioactive Added Value Compounds from Citrus Wastes—A Review. Molecules 2017, 22, 680. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green extraction of natural products: Concept and principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed]
- Londono-Londono, J.; Lima, V.R.; Lara, O.; Gil, A.; Pasa, T.B.C.; Arango, G.J.; Pineda, J.R.R. Clean recovery of antioxidant flavonoids from citrus peel: Optimizing an aqueous ultrasound-assisted extraction method. Food Chem. 2010, 119, 81–87. [Google Scholar] [CrossRef]
- Singanusong, R.; Nipornram, S.; Tochampa, W.; Rattanatraiwong, P. Low power ultrasound-assisted extraction of phenolic compounds from mandarin (Citrus reticulata Blanco cv. Sainampueng) and lime (Citrus aurantifolia) peels and the antioxidant. Food Anal. Methods 2015, 8, 1112–1123. [Google Scholar]
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M.; Bansode, R.R. Agricultural by-products as important food sources of polyphenols. In Polyphenols: Food Sources, Bioactive Properties, and Antioxidant Effects; Cobb, D.T., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2014; pp. 1–32. [Google Scholar]
- Yu, J.; Wang, L.; Walzem, R.L.; Miller, E.G.; Pike, L.M.; Patil, B.S. Antioxidant activity of citrus limonoids, flavonoids, and coumarins. J. Agric. Food Chem. 2005, 53, 2009–2014. [Google Scholar] [CrossRef]
- Yoo, K.M.; Hwang, I.K.; Park, J.H.; Moon, B. Major phytochemical composition of 3 native Korean citrus varieties and bioactive activity on V79-4 cells induced by oxidative stress. J. Food Sci. 2009, 74, 462–468. [Google Scholar] [CrossRef]
- Kim, H.G.; Kim, G.; Park, S.; Lee, J.H.; Seo, O.N.; Lee, S.J.; Kim, J.H.; Shim, J.; Abd El-Aty, A.M.; Jin, J.S. Flavonoid profiling in three citrus varieties native to the Republic of Korea using liquid chromatography coupled with tandem mass spectrometry: Contribution to overall antioxidant activity. Biomed. Chromatogr. 2012, 26, 464–470. [Google Scholar] [CrossRef]
- Hirata, T.; Fujii, M.; Akita, K.; Yanaka, N.; Ogawa, K.; Kuroyanagi, M.; Hongo, D. Identification and physiological evaluation of the components from Citrus fruits as potential drugs for anti-corpulence and anticancer. Bioorg Med. Chem. 2009, 17, 25–28. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, A.; Moriguchi, T.; Koyama, K.; Omura, M.; Akihama, T. Analysis of sucrose synthase genes in citrus suggests different roles and phylogenetic relationships. J. Exp. Bot. 2002, 53, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Albertini, M.V.; Carcouet, E.; Pailly, O.; Gambotti, C.; Luro, F.; Berti, L. Changes in organic acids and sugars during early stages of development of acidic and acidless citrus fruit. J. Agric. Food Chem. 2006, 54, 8335–8339. [Google Scholar] [CrossRef] [PubMed]
- Tezcan, F.; Gültekin-Özgüven, M.; Diken, T.; Özçelik, B.; Erim, F.B. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juices. Food Chem. 2009, 115, 873–877. [Google Scholar] [CrossRef]
- Wang, S.; Tu, H.; Wan, J.; Chen, W.; Liu, X.; Luo, J.; Xu, J.; Zhang, H. Spatio-temporal distribution and natural variation of metabolites in citrus fruits. Food Chem. 2016, 199, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Proch, J.; Bose, P. Determination of quantity and quality of polyphenol antioxidants in foods and beverages. Methods Enzym. 2001, 335, 103–114. [Google Scholar]
- Proteggente, A.R.; Pannala, A.S.; Paganga, G.; Van Buren, L.; Wagner, E.; Wiseman, S.; Van De Put, F.; Dacombe, C.; Rice-Evans, C.A. The antioxidant activity of regularly consumed fruit and vegetables reflects their phenolic and vitamin C composition. Free Radical. Res. 2002, 36, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Wilmsen, P.K.; Spada, D.S.; Salvador, M. Antioxidant activity of the flavonoid hesperidin in chemical and biological systems. J. Agric. Food Chem. 2005, 53, 4757–4761. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Martínez-Florensa, M.; Espín, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T. A Citrus extract containing flavanones represses plasminogen activator inhibitor-1 (PAI-1) expression and regulates multiple inflammatory, tissue repair, and fibrosis genes in human colon fibroblasts. J. Agric. Food Chem. 2009, 57, 9305–9315. [Google Scholar] [CrossRef]
- Benavente-García, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Yu, J.H. Hesperidin enhances intestinal barrier function in Caco-2 cell monolayers via AMPK-mediated tight junction-related proteins. FEBS Open Biol. 2023, 13, 532–544. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, M.; Li, Y.; Yu, H.; Wang, D.; Chen, Q.; Chen, Y.; Zhang, Y.; Wang, T. Flavonoids from Citrus aurantium ameliorate TNBS-induced ulcerative colitis through protecting colonic mucus layer integrity. Eur. J. Pharmacol. 2019, 857, 172456. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Li, Y.; Deng, L.; Ren, R.; Ge, B.; Liao, Z.; Xiang, S.; Zhou, B. Alisol B 23-Acetate Ameliorates Lipopolysaccharide-Induced Intestinal Barrier Dysfunction by Inhibiting TLR4-NOX1/ROS Signaling Pathway in Caco-2 Cells. Front. Pharmacol. 2022, 13, 911196. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Hanate, M.; Aw, W.; Itoh, H.; Saito, K.; Kobayashi, S.; Hachimura, S.; Fukuda, S.; Tomita, M.; Hasebe, Y.; et al. Eggshell membrane powder ameliorates intestinal inflammation by facilitating the restitution of epithelial injury and alleviating microbial dysbiosis. Sci. Rep. 2017, 7, 43993. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lin, T.Y.; Fan, C.W.; Maa, M.C.; Leu, T.H. Lipopolysaccharide-promoted proliferation of Caco-2 cells is mediated by c-Src induction and ERK activation. Biomedicine 2015, 5, 5. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, B.; Wei, X.; Ding, M.; Luo, Z.; Tan, X.; Zheng, Z. Daidzein Protects Caco-2 Cells against Lipopolysaccharide-Induced Intestinal Epithelial Barrier Injury by Suppressing PI3K/AKT and P38 Pathways. Molecules 2022, 27, 8928. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saha, S.; Pupo, E.; Zariri, A.; van der Ley, P. Lipid A heterogeneity and its role in the host interactions with pathogenic and commensal bacteria. Microlife 2022, 3, uqac011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Peng, J.; Li, H.; Olaolu, O.A.; Ibrahim, S.; Ibrahim, S.; Wang, S. Natural Products: A Dependable Source of Therapeutic Alternatives for Inflammatory Bowel Disease through Regulation of Tight Junctions. Molecules 2023, 28, 6293. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front. Endocrinol. 2023, 14, 1217165. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Topa, J.; Tańska, M.; Cieślińska, A.; Fiedorowicz, E.; Savelkoul, H.F.J.; Jarmołowska, B. Modulatory Effects of Osthole on Lipopolysaccharides-Induced Inflammation in Caco-2 Cell Monolayer and Co-Cultures with THP-1 and THP-1-Derived Macrophages. Nutrients 2020, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Fontana, G.; Bruno, M.; Sottile, F.; Badalamenti, N. The Chemistry and the Anti-Inflammatory Activity of Polymethoxyflavonoids from Citrus Genus. Antioxidants 2022, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Grohmann, K. Concentrations of hesperidin and other orange peel flavonoids in Citrus processing byproducts. J. Agric. Food Chem. 1996, 44, 811–814. [Google Scholar] [CrossRef]
- Bodduluru, L.N.; Kasala, E.R.; Madhana, R.M.; Barua, C.C.; Hussain, M.I.; Haloi, P.; Borah, P. Naringenin ameliorates inflammation and cell proliferation in benzo(a)pyrene induced pulmonary carcinogenesis by modulating CYP1A1, NFjB and PCNA expression. Int. Immunopharmacol. 2016, 30, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Guazelli, C.F.S.; Fattori, V.; Ferraz, C.R.; Borghi, S.M.; Casagrande, R.; Baracat, M.M.; Verri, W.A., Jr. Antioxidant and anti-inflammatory effects of hesperidin methyl chalcone in experimental ulcerative colitis. Chem. Biol. Interact. 2021, 333, 109315. [Google Scholar] [CrossRef] [PubMed]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Song, Y.; Xu, Y.; Xu, H. Manipulating Microbiota in Inflammatory Bowel Disease Treatment: Clinical and Natural Product Interventions Explored. Int. J. Mol. Sci. 2023, 24, 11004. [Google Scholar] [CrossRef]
- Turner, J. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Diagnostic Signal (δ, Multiplicity *) | LE (mg/g DW) | OE (mg/g DW) |
|---|---|---|---|
| alanine | 1.45, d | 3.6 ± 0.1 | 1.9 ± 0.1 |
| α-glucose | 5.20, d | 109.3 ± 1.4 | 108.2 ± 1.0 |
| β-glucose | 4.6, d | 183.6 ± 2.4 | 197.5 ± 2.0 |
| sucrose | 5.4, d | 76.5 ± 1.0 | 224.8 ± 2.3 |
| fructose | 3.87, dd | 318.1 ± 4.0 | 347.6 ± 3.2 |
| citric acid | 2.71, d | 210.7 ± 4.6 | 47.2 ± 1.0 |
| proline | 2.09, m | 8.0 ± 0.1 | 12.9 ± 0.3 |
| GABA | 3.02, t | 2.5 ± 0.1 | 4.42 ± 0.1 |
| asparagine | 2.95, dd | 21.5 ± 1.2 | n.d. § |
| aspartic acid | 2.6, dd | 23.9 ± 0.7 | n.d. |
| succinic acid | 2.47, s | n.d. | 0.4 ± 0.1 |
| tyrosine | 7.28, d | n.d. | 3.1 ± 0.1 |
| malic acid | 4.3, dd | n.d. | 15.7 ± 0.1 |
| Phytochemical Assay | OE | LE |
|---|---|---|
| Total phenols (g GAE a/100 g DE b) | 2.41 ± 0.16 | 2.46 ± 0.14 |
| Flavonoids (g RE c/100 g DE) | 1.36 ± 0.09 | 1.53 ± 0.08 |
| Vanillin index (mg CE d/100 g DE) | 0.36 ± 0.01 | 0.46 ± 0.02 ** |
| Proanthocyanidins (mg CyE e/100 g DE) | 0.003 ± 0.000 | 0.004 ± 0.000 |
| Polymerization index f | 139.17 | 111.41 ** |
| Compound Name | RT b (min) | Molecular Formula | Molecular Weight | [M−H]− (m/z) | [M+H]+ (m/z) | OE c | LE d |
|---|---|---|---|---|---|---|---|
| 6-Hydroxyapigenin (Scutellarein) a | 16.1 | C15H10O6 | 286 | 287 | − | + | |
| Luteolin-8-glucoside (Orientin) a | 16.2 | C21H20O11 | 448 | 447 | − | + | |
| Apigenin 6-C-glucoside 8-C-arabinoside | 16.8 | C26H28O14 | 564 | 565 | + | − | |
| Kaempferol 7-O-glucoside a | 17.4 | C21H20O11 | 448 | 449 | − | + | |
| Dihydroferulic acid 4-O-glucuronide | 17.5 | C16H20O10 | 372 | 371 | − | + | |
| Dihydrocaffeic acid dimer | 18.7 | C18H20O8 | 364 | 365 | − | + | |
| Heptyl caffeate | 19.4 | C16H22O4 | 278 | 277 | − | + | |
| Feruloylisocitric acid | 19.6 | C16H16O10 | 368 | 367 | + | − | |
| Naringenin 7-O-glucoside a | 19.8 | C21H22O10 | 434 | 433 | + | − | |
| Hydroxycaffeic acid | 20.0 | C9H8O5 | 196 | 197 | + | − | |
| Apigenin 7-O-rutinoside a | 20.4 | C27H30O14 | 578 | 579 | + | + | |
| Apigenin 7,4′-diglucoside | 21.4 | C27H30O15 | 594 | 595 | − | + | |
| Diosmetin 3′-O-glucuronide | 21.7 | C22H20O12 | 476 | 475 | + | − | |
| Quercetin 3-O-galactoside (Hyperoside) a | 22.6 | C21H20O12 | 464 | 463 | − | + | |
| Sakuranin | 22.7 | C22H24O10 | 448 | 449 | + | + | |
| Diosmetin 6,8-di-C-glucoside | 23.2 | C28H32O16 | 624 | 625 | + | + | |
| Diosmetin-7-O-glucoside a | 23.4 | C22H22O11 | 462 | 463 | − | + | |
| Chrysoeriol-C-glucoside | 25.1 | C22H22O11 | 462 | 463 | − | + | |
| Kaempferol 3-O-rhamnoside a | 25.2 | C21H20O10 | 432 | 433 | + | − | |
| Quercetin 3-rutinoside (Rutin) a | 26.4 | C27H30O16 | 610 | 611 | + | + | |
| Apigenin 6,8-C-diglucoside (Vicenin 2) | 27.4 | C27H30O15 | 594 | 595 | + | − | |
| Isosakuranetin-7-O-rutinoside (Didymin) a | 28.0 | C28H34O14 | 594 | 593 | 595 | + | + |
| Kaempferol-3-O-rutinoside a | 28.4 | C27H30O15 | 594 | 595 | + | − | |
| Perilloside A | 28.9 | C16H26O6 | 314 | 313 | − | + | |
| Isorhamnetin 3-O-rutinoside (Narcissin) a | 29.0 | C28H32O16 | 624 | 623 | 625 | − | + |
| Hesperetin-glucuronide-sulfate | 29.2 | C22H22O15S | 558 | 559 | + | − | |
| Quercetin-3-O-sophoroside (Baimaside) | 29.4 | C27H30O17 | 626 | 625 | − | + | |
| Limonin glucoside | 29.9 | C32H42O14 | 650 | 649 | + | − | |
| Sinigrin | 30.0 | C10H16KNO9S2 | 397 | 396 | + | − | |
| Eriodictyol-7-O-rutinoside (Eriocitrin) a | 30.2 | C27H32O15 | 596 | 595 | 597 | − | + |
| 3-Hydroxyphloretin 2′-O-xylosyl-glucoside | 30.5 | C26H32O15 | 584 | 585 | + | − | |
| Eriodictyol 7-O-neohesperidoside (Neoeriocitrin) a | 31.2 | C27H32O15 | 596 | 595 | 597 | − | + |
| Diosmetin-7-O-rutinoside (Diosmin) a | 31.4 | C28H32O15 | 608 | 607 | 609 | + | + |
| 5,3′,4′-Trihydroxy-3-methoxy-6,7-methylenedioxyflavone 4′-O-glucuronide | 31.9 | C23H20O14 | 520 | 519 | + | − | |
| Hesperetin-3′,5,7-tri-sulfate | 32.1 | C16H14O14S3 | 525 | 526 | + | − | |
| Limocitrin O-3-hdroxy-3-methylglutaryl (HMG)-glucoside | 32.2 | C29H32O17 | 652 | 653 | + | + | |
| Narirutin a | 32.4 | C27H32O14 | 580 | 579 | + | − | |
| Apigenin 7,4′-diglucoside | 32.9 | C27H30O15 | 594 | 593 | − | + | |
| Apigenin 7-sulfate | 33.0 | C15H10O8S | 350 | 351 | + | − | |
| Limonin | 33.2 | C26H30O8 | 470 | 471 | − | + | |
| Hesperetin-7-rutinoside (Hesperidin) a | 33.7 | C28H34O15 | 610 | 609 | 611 | + | + |
| Hesperetin-7-neohesperidoside (Neohesperidin) a | 34.6 | C28H34O15 | 610 | 609 | 611 | + | + |
| Naringenin hexosyl-deoxyhexoside | 35.0 | C27H32O14 | 580 | 581 | − | + | |
| Methyl-limonexic acid | 35.6 | C27H32O10 | 516 | 517 | + | − | |
| Diosmetin-7-O-neohesperidoside (Neodiosmin) a | 35.9 | C28H32O15 | 608 | 607 | 609 | − | + |
| Kaempferol 3-O-(6″-acetyl-galactoside) 7-O-rhamnoside | 36.1 | C29H32O16 | 636 | 637 | + | − | |
| Nomilin hexoside | 36.4 | C34H46O15 | 694 | 693 | + | + | |
| Quercetin 3-rhamnoside (Quercitrin) a | 36.6 | C21H20O11 | 448 | 447 | 449 | + | + |
| Nomilinic acid-O-hexoside | 38.2 | C34H48O16 | 712 | 711 | + | + | |
| Pelargonidin 3-O-(6″-succinyl-glucoside) | 38.7 | C25H24O13 | 533 | 534 | + | − | |
| Isosakuranetin-7-O-neohesperidoside (Poncirin) a | 39.2 | C28H34O14 | 594 | 593 | + | − | |
| Cyanidin 3-O-xylosyl-rutinoside | 41.8 | C32H38O19 | 727 | 726 | 728 | + | − |
| Kaempferol O-synapoyl-caffeoyl-sophoroside-O-hexoside | 42.9 | C53H56O28 | 1141 | 1139 | + | − | |
| Kaempferol-isorhamninoside-rhamnoside | 43.7 | C39H50O23 | 886 | 887 | + | + | |
| Ichangin | 44.5 | C26H32O9 | 488 | 487 | + | − | |
| Acacetin (di-deoxyhexosyl)-hexoside | 45.7 | C34H42O18 | 738 | 737 | + | − | |
| Hydroxy-pentamethoxyflavanone (Norcitromitin) | 46.2 | C20H22O8 | 390 | 389 | 391 | + | − |
| Naringin a | 47.6 | C27H32O14 | 580 | 581 | + | + | |
| Feruloylquinic acid | 48.6 | C17H20O9 | 368 | 369 | + | + | |
| 4′,5,6,7,8-Pentamethoxyflavone (Tangeretin) a | 49.3 | C20H20O7 | 372 | 373 | + | − | |
| Nomilinic acid | 50.0 | C28H36O10 | 532 | 531 | + | + | |
| Peonidin 3-O-glicoside | 51.2 | C22H22O11 | 462 | 463 | + | − | |
| Hesperetin-7-sulfate-3′,5-di-glucuronide | 52.1 | C28H26O20S | 718 | 719 | + | − | |
| 6-Demethoxytangeretin | 55.2 | C19H18O6 | 342 | 343 | + | + | |
| Sinapic acid a | 55.3 | C11H12O5 | 224 | 223 | − | + | |
| Pelargonidin 3-O-sambubioside | 56.4 | C26H28O14 | 565 | 566 | + | − | |
| 3,7-Di-O-methylquercetin | 56.5 | C17H14O7 | 330 | 331 | − | + | |
| Eriodictyol a | 58.1 | C15H12O6 | 288 | 289 | + | + | |
| Deacetylnomilin | 58.8 | C26H32O8 | 472 | 473 | − | + | |
| 4′-O-Methylkaempferol (Kaempferide) a | 62.0 | C16H12O6 | 300 | 301 | + | + | |
| 5,6-Dihydroxy-7,8,3′,4′-tetramethoxyflavone (Pebrellin) | 63.6 | C19H18O8 | 374 | 375 | + | − | |
| Homoeriodictyol chalcone a | 63.7 | C16H14O6 | 302 | 303 | − | + | |
| Desmethyltangeretin (Gardenin B) | 66.1 | C19H18O7 | 358 | 359 | + | + | |
| Desmethylnobiletin | 66.4 | C20H20O8 | 388 | 389 | − | + | |
| 4′,5-dihydroxy-6,7,8-trimethoxyflavone (Xanthomicrol) | 70.6 | C18H16O7 | 344 | 343 | − | + | |
| Naringenin-sulfate | 71.2 | C15H10O9S | 366 | 367 | + | − | |
| Citrusin III | 71.4 | C36H53N7O9 | 727 | 728 | − | + | |
| 5-O-Methylmikanin | 75.0 | C19H18O7 | 358 | 359 | + | + | |
| Kaempferol 3,5-dimethyl ether | 80.3 | C17H14O6 | 314 | 315 | + | + | |
| Naringenin-4′-methylether (Isosakuranetin) a | 80.6 | C16H14O5 | 286 | 285 | + | + |
| Test | OE | LE | OE | LE | RS |
|---|---|---|---|---|---|
| g RSE a/100 g DE | g RSE a/100 g DE | IC50 (µg/mL) | |||
| DPPH | 0.77 ± 0.10 *** | 1.61 ± 0.16 | 3810.09 (2231.51–4505.38) *** | 1015.23 (803.66–1282.50) | 11.62 (9.82–13.75) |
| TEAC | 3.95 ± 0.29 | 2.44 ± 0.45 | 127.26 (105.28–153.81) | 182.49 (152.76–218.00) | 3.73 (1.51–9.24) |
| FRAP | 1.80 ± 0.09 | 1.71 ± 0.12 | 119.80 (112.66–354.32) | 199.15 (119.14–332.89) | 3.68 (1.61–8.46) |
| ORAC | 15.13 ± 1.10 | 15.19 ± 0.22 | 7.36 (5.81–9.34) | 5.98 (4.93–7.25) | 0.67 (0.20–1.16) |
| BCB | 0.82 ± 0.03 ** | 1.33 ± 0.05 | 76.65 (54.72–107.36) * | 41.60 (32.68–52.96) | 0.32 (0.15–0.55) |
| ICA | 39.92 ± 3.01 *** | 50.27 ± 0.58 | 2140.12 (1329.71–3444.44) *** | 535.88 (424.92–675.82) | 5.65 (2.50–7.75) |
| ADA | 2.28 ± 0.21 *** | 11.29 ± 1.01 | 3825.66 (2961.86–4941.34) *** | 785.09 (188.93–1362.44) | 29.67 (17.56–50.14) |
| PIA | 79.63 ± 1.77 ** | 92.17 ± 4.88 | 210.15 (184.46–239.41) | 151.14 (95.83–238.38) | 28.75 (14.41–57.34) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingegneri, M.; Braghini, M.R.; Piccione, M.; De Stefanis, C.; Mandrone, M.; Chiocchio, I.; Poli, F.; Imbesi, M.; Alisi, A.; Smeriglio, A.; et al. Citrus Pomace as a Source of Plant Complexes to Be Used in the Nutraceutical Field of Intestinal Inflammation. Antioxidants 2024, 13, 869. https://doi.org/10.3390/antiox13070869
Ingegneri M, Braghini MR, Piccione M, De Stefanis C, Mandrone M, Chiocchio I, Poli F, Imbesi M, Alisi A, Smeriglio A, et al. Citrus Pomace as a Source of Plant Complexes to Be Used in the Nutraceutical Field of Intestinal Inflammation. Antioxidants. 2024; 13(7):869. https://doi.org/10.3390/antiox13070869
Chicago/Turabian StyleIngegneri, Mariarosaria, Maria Rita Braghini, Michela Piccione, Cristiano De Stefanis, Manuela Mandrone, Ilaria Chiocchio, Ferruccio Poli, Martina Imbesi, Anna Alisi, Antonella Smeriglio, and et al. 2024. "Citrus Pomace as a Source of Plant Complexes to Be Used in the Nutraceutical Field of Intestinal Inflammation" Antioxidants 13, no. 7: 869. https://doi.org/10.3390/antiox13070869
APA StyleIngegneri, M., Braghini, M. R., Piccione, M., De Stefanis, C., Mandrone, M., Chiocchio, I., Poli, F., Imbesi, M., Alisi, A., Smeriglio, A., & Trombetta, D. (2024). Citrus Pomace as a Source of Plant Complexes to Be Used in the Nutraceutical Field of Intestinal Inflammation. Antioxidants, 13(7), 869. https://doi.org/10.3390/antiox13070869