
Antioxidant Capacity, Lipid Oxidation, and Quality Traits of Slow- and Fast-Growing Meagre (Argyrosomus regius) Fillets During Cold Storage


 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Samples and Somatometric Traits
2.2. Proximate Composition and Fatty Acid Content
2.3. Total Antioxidant Capacity, SOD Activity, and Lipid Oxidation
2.4. Muscle Ceullularity
2.5. Chemicals
2.6. Statistical Analysis
3. Results and Discussion
3.1. Body Traits and Proximate Composition of Muscle Tissue
3.2. Fatty Acid (FA) Analysis
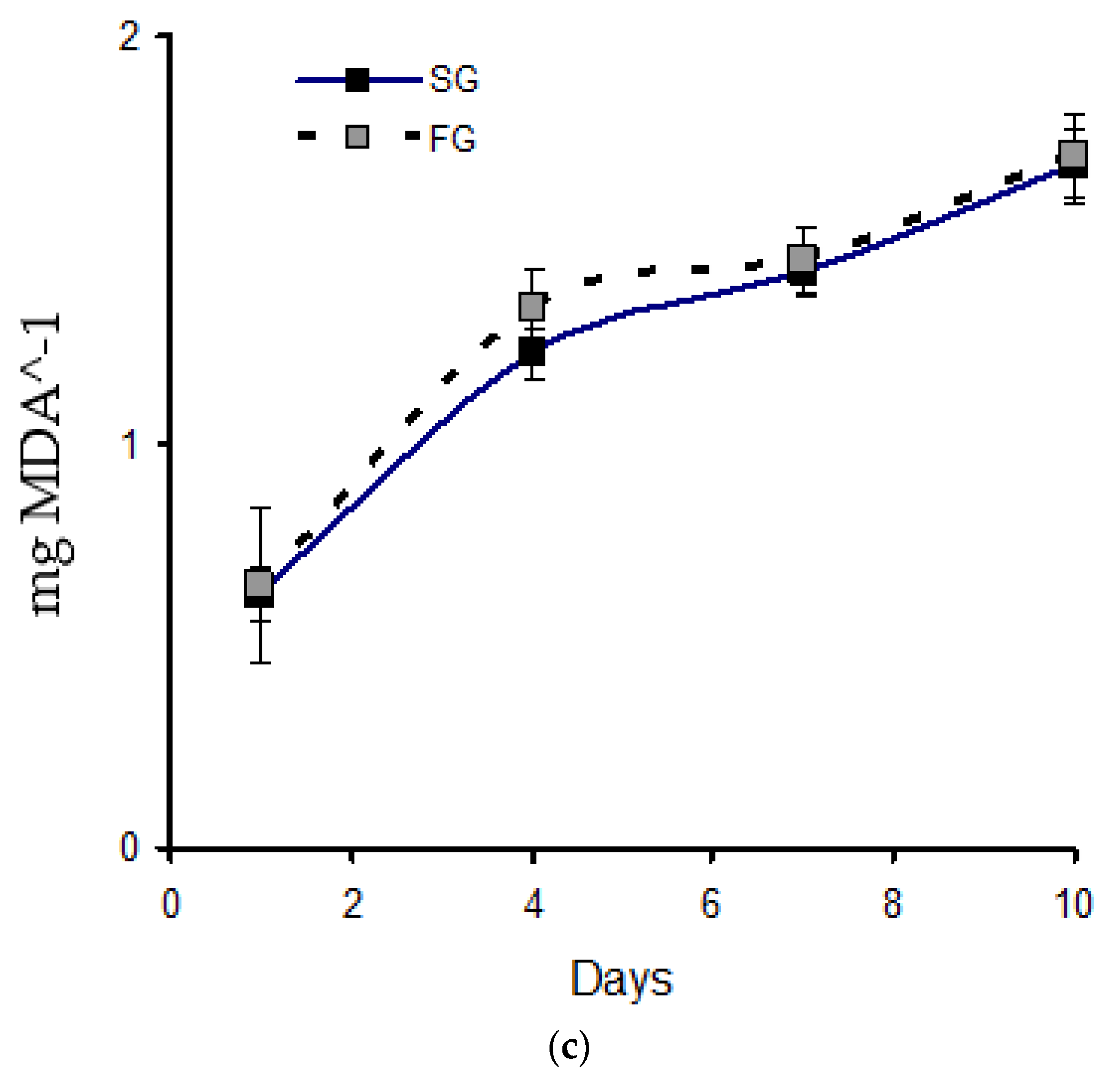
3.3. Total Antioxidant Capacity (TOAC), SOD Activity, and Lipid Oxidation
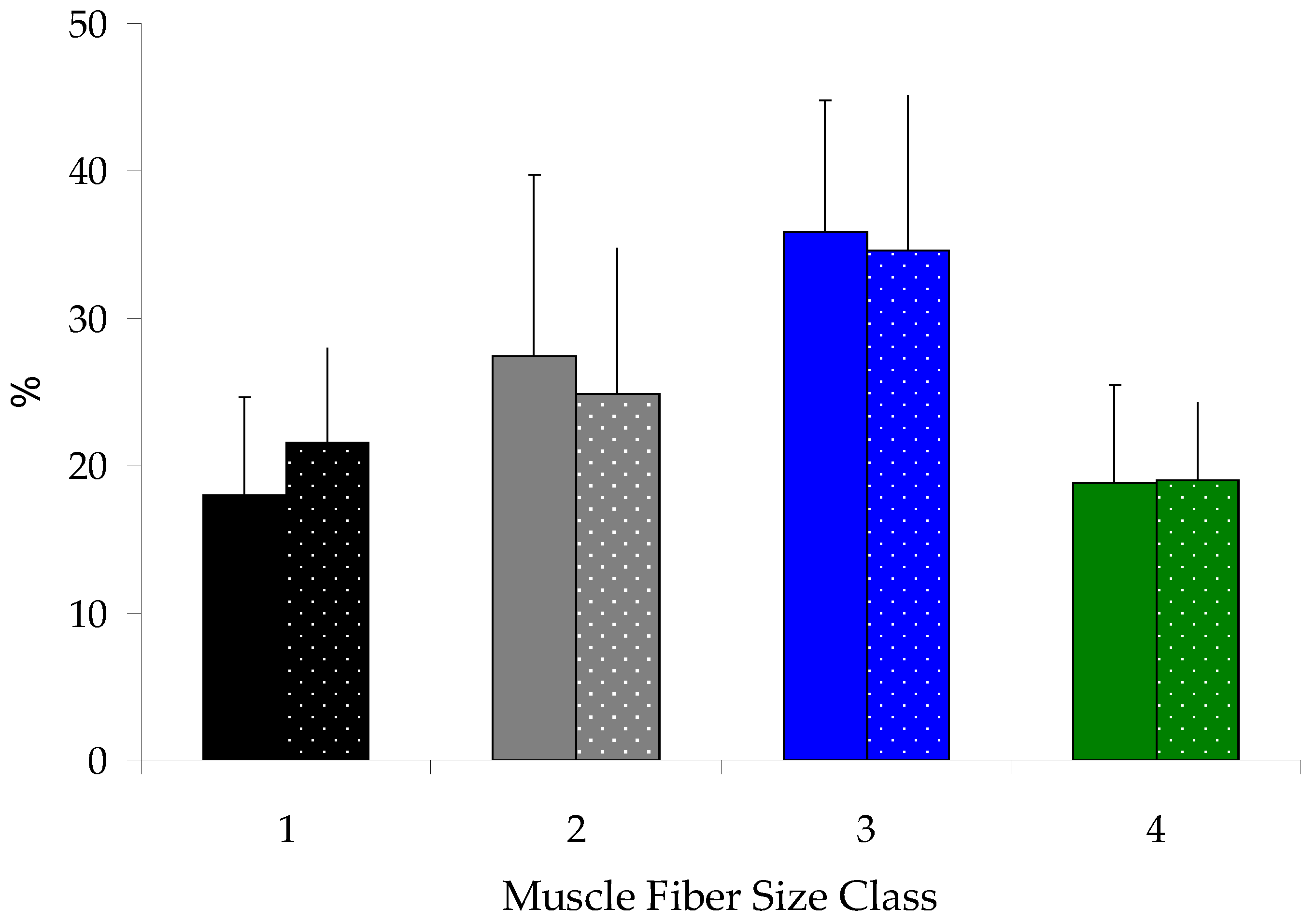
3.4. White Muscle Fiber Cellularity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giogios, I.; Grigorakis, K.; Kalogeropoulos, N. Organoleptic and Chemical Quality of Farmed Meagre (Argyrosomus regius) as Affected by Size. Food Chem. 2013, 141, 3153–3159. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.; Pereira, T.G.; Carvalho, L.M.; Pousão-Ferreira, P.; Grade, A.; Teixeira, B.; Quental-Ferreira, H.; Mendes, R.; Bandarra, N.; Gonçalves, A. Wild and Farmed Meagre, Argyrosomus regius: A Nutritional, Sensory and Histological Assessment of Quality Differences. J. Food Compos. Anal. 2017, 63, 8–14. [Google Scholar] [CrossRef]
- Fountoulaki, E.; Grigorakis, K.; Kounna, C.; Rigos, G.; Papandroulakis, N.; Diakogeorgakis, J.; Kokou, F. Growth Performance and Product Quality of Meagre (Argyrosomus regius) Fed Diets of Different Protein/Lipid Levels at Industrial Scale. Ital. J. Anim. Sci. 2017, 16, 685–694. [Google Scholar] [CrossRef]
- Vallecillos, A.; María-Dolores, E.; Villa, J.; Rueda, F.M.; Carrillo, J.; Ramis, G.; Soula, M.; Afonso, J.M.; Armero, E. Phenotypic and Genetic Components for Growth, Morphology, and Flesh-Quality Traits of Meagre (Argyrosomus regius) Reared in Tank and Sea Cage. Animals 2021, 11, 3285. [Google Scholar] [CrossRef]
- Vallecillos, A.; María-Dolores, E.; Villa, J.; Afonso, J.M.; Armero, E. Potential Use of Image Analysis in Breeding Programs for Growth and Yield Traits in Meagre (Argyrosomus regius). J. Mar. Sci. Eng. 2023, 11, 2067. [Google Scholar] [CrossRef]
- Benhaïm, D.; Péan, S.; Brisset, B.; Leguay, D.; Bégout, M.-L.; Chatain, B. Effect of Size Grading on Sea Bass (Dicentrarchus labrax) Juvenile Self-Feeding Behaviour, Social Structure and Culture Performance. Aquat. Living Resour. 2011, 24, 391–402. [Google Scholar] [CrossRef]
- Batzina, A.; Drossos, I.-P.; Giannoudaki, K.; Karakatsouli, N. Effects of Size Variability on Individual Growth and Feeding Behavior of European Seabass. Appl. Anim. Behav. Sci. 2020, 225, 104963. [Google Scholar] [CrossRef]
- Karipoglou, C.; Nathanailides, C. Growth Rate and Feed Conversion Efficiency of Intensively Cultivated European Eel (Anguilla anguilla L.). Int. J. Fish Aquacult. 2009, 1, 11–13. [Google Scholar]
- Duan, Z.; Zhou, Y.; Liu, W.; Shi, C.C.; Li, L.; Dong, Y.; Gao, Q.; Dong, S. Variations in Flavor According to Fish Size in Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2020, 526, 735398. [Google Scholar] [CrossRef]
- Morais, C.A.R.S.; Santana, T.P.; Santos, C.A.; Passetti, R.A.C.; Melo, J.F.B.; de Macedo, F.d.A.F.; Vieira, J.S.; Teixeira, A.J.C.; Del Vesco, A.P. Effect of Slaughter Weight on the Quality of Nile Tilapia Fillets. Aquaculture 2020, 520, 734941. [Google Scholar] [CrossRef]
- Saavedra, M.; Pereira, T.G.; Candeias-Mendes, A.; Carvalho, L.; Pousão-Ferreira, P.; Conceição, L.E.C. Effect of Increased Dietary Protein Level in Meagre (Argyrosomus regius) Juvenile Growth and Muscle Cellularity. Aquac. Nutr. 2018, 24, 1153–1159. [Google Scholar] [CrossRef]
- Skałecki, P.; Kaliniak-Dziura, A.; Domaradzki, P.; Florek, M.; Kępka, M. Fatty Acid Composition and Oxidative Stability of the Lipid Fraction of Skin-On and Skinless Fillets of Prussian Carp (Carassius gibelio). Animals 2020, 10, 778. [Google Scholar] [CrossRef]
- Matulić, D.; Blažina, M.; Pritišanac, E.; Čolak, S.; Bavčević, L.; Barić, R.; Križanac, S.; Vitlov, B.; Šuran, J.; Strunjak Perović, I. Growth, Fatty Acid Profile and Malondialdehyde Concentration of Meagre Argyrosomus regius Fed Diets with Different Lipid Content. Appl. Sci. 2024, 14, 4842. [Google Scholar] [CrossRef]
- Vallecillos, A.; Marín, M.; Bortoletti, M.; López, J.; Afonso, J.M.; Ramis, G.; Arizcun, M.; María-Dolores, E.; Armero, E. Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.). Animals 2021, 11, 2889. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.D.; Santaella, M.; Martínez, C.; Periago, M.J.; Blanco, A.; Vázquez, J.M.; Albors, O.L. Muscle Tissue Structure and Flesh Texture in Gilthead Sea Bream, Sparus aurata L., Fillets Preserved by Refrigeration and by Vacuum Packaging. LWT-Food Sci. Technol. 2011, 44, 1098–1106. [Google Scholar] [CrossRef]
- Wang, Z.; Qiao, F.; Zhang, W.; Parisi, G.; Du, Z.-Y.; Zhang, M. The Flesh Texture of Teleost Fish: Characteristics and Interventional Strategies. Rev. Aquac. 2023, 16, 508–535. [Google Scholar] [CrossRef]
- Nathanailides, C.; Panopoulos, S.; Kakali, F.; Karipoglou, C.; Lenas, D. Antemortem and Postmortem Biochemistry, Drip Loss and Lipid Oxidation of European Sea Bass Muscle Tissue. Procedia Food Sci. 2011, 1, 1099–1104. [Google Scholar] [CrossRef]
- Palanisamy, S.; Singh, A.; Zhang, B.; Zhao, Q.; Benjakul, S. Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices. Foods 2024, 13, 1238. [Google Scholar] [CrossRef]
- Grigorakis, K. Fillet Proximate Composition, Lipid Quality, Yields, and Organoleptic Quality of Mediterranean-Farmed MarineFish: A Review with Emphasis on New Species. Crit. Rev. Food Sci. Nutr. 2017, 57, 2956–2969. [Google Scholar] [CrossRef]
- Gao, X.; Zhai, H.; Peng, Z.; Yu, J.; Yan, L.; Wang, W.; Ren, T.; Han, Y. Comparison of Nutritional Quality, Flesh Quality, Muscle Cellularity, and Expression of Muscle Growth-Related Genes between Wild and Recirculating Aquaculture System (RAS)-Farmed Black Rockfish (Sebastes schlegelii). Aquac. Int. 2023, 31, 2263–2280. [Google Scholar] [CrossRef]
- Tsoupras, A.; Brummell, C.; Kealy, C.; Vitkaitis, K.; Redfern, S.; Zabetakis, I. Cardio-Protective Properties and Health Benefits of Fish Lipid Bioactives; The Effects of Thermal Processing. Mar. Drugs 2022, 20, 187. [Google Scholar] [CrossRef]
- Pazos, M.; González, M.J.; Gallardo, J.M.; Torres, J.L.; Medina, I. Preservation of the endogenous antioxidant system of fish muscle by grape polyphenols during frozen storage. Eur. Food Res. Technol. 2005, 220, 514–519. [Google Scholar] [CrossRef]
- Celano, R.; Campone, L.; Piccinelli, A.L.; Acernese, F.; Nabavi, S.M.; Di Bella, G.; Rastrelli, L. Fatty acid composition, antioxidant levels and oxidation products development in the muscle tissue of Merluccius merluccius and Dicentrarchus labrax during ice storage. LWT-Food Sci. Technol. 2016, 73, 654–662. [Google Scholar] [CrossRef]
- Nathanailides, C.; Lopez-Albors, O.; Lenas, D.; Athanassopoulou, F.; Grigorakis, K. Changes in Cytochrome C Oxidase (CcO) and Lactate Dehydrogenase (LDH) Enzyme Activity of the White Epaxial Muscle Tissue of Sea Bass (Dicentrarchus labrax L.) as Shelf-Life Predictive Indices during Ice-Storage. Ital. J. Food Sci. 2010, 22, 200–206. [Google Scholar]
- Prista, N.; Gordo, L.; Costa, J.L.; Costa, M.J.; Jones, C. Reproductive phase determination in male meagre (Argyrosomus regius, Sciaenidae): Testis development and histologic corroboration of a gross anatomical scale. Sci. Mar. 2014, 78, 65–80. [Google Scholar] [CrossRef]
- Follesa, M.C.; Carbonara, P. Atlas of the Maturity Stages of Mediterranean Fishery Resources. Gen. Fish. Comm. Mediterr. Stud. Rev. 2019, 99, 1–259. [Google Scholar]
- Ayala, M.D.; Hernández-Urcera, J.; Santaella, M.; Graciá, M.C.M.; López-Albors, O.; Cal, R. Influence of the Myotome Zone and Sex on the Muscle Cellularity and Fillet Texture of Diploid and Triploid Turbots Scophthalmus maximus L. Vet. Res. Forum 2020, 11, 105–112. [Google Scholar]
- Horowitz, W.; Latimer, G.W. Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 2006; Volume 18. [Google Scholar]
- Fernando, F.; Candebat, C.L.; Strugnell, J.M.; Andreakis, N.; Nankervis, L. Dietary Supplementation of Astaxanthin Modulates Skin Color and Liver Antioxidant Status of Giant Grouper (Epinephelus lanceolatus). Aquacult. Rep. 2022, 26, 101266. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Chatzigiannidou, I.; Feidantsis, K.; Kounna, C.; Chatzifotis, S. Effect of water temperature on cellular stress responses in meagre (Argyrosomus regius). Fish. Physiol. Biochem. 2020, 46, 1075–1091. [Google Scholar] [CrossRef]
- Nathanailides, C.; Lopez-Albors, O.; Abellán, E.; Vazquez, J.M.; Tyler, D.D.; Rowlerson, A.; Stickland, N.C. Muscle cellularity in relation to somatic growth in the European sea bass Dicentrarchus labrax (L.). Aquac. Res. 1996, 27, 885–889. [Google Scholar] [CrossRef]
- Grigorakis, K.; Fountoulaki, E.; Vasilaki, A.; Mittakos, I.; Nathanailides, C. Lipid quality and filleting yield of reared meagre (Argyrosomus regius). Int. J. Food Sci. Techn. 2011, 46, 711–716. [Google Scholar] [CrossRef]
- Kiessling, A.; Ruohonen, K.; Bjørnevik, M. Muscle Fibre Growth and Quality in Fish. Arch. Tierz. Dummerstorf. 2016, 49, 137–146. [Google Scholar]
- Saavedra, M.; Pereira, T.G.; Grade, A.; Barbeiro, M.; Ferreira, P.; Quental-Ferreira, H.; Nunes, M.L.; Bandara, N.; Goncalves, A. Farmed meagre, Argyrosomus regius of three different sizes: What are the differences in flesh quality and muscle cellularity? Inter. J. Food Sci. Tech. 2015, 50, 1311–1316. [Google Scholar] [CrossRef]
- Martelli, R.; Dalle Zotte, A.; Bonelli, A.; Paola Lupi, P.; Franci, O.; Parisi, G. Macronutrient and fatty acid profiles of meagre (Argyrosomus regius) fillets as influenced by harvesting time and boiling. Ital. J. Anim. Sci. 2013, 12, 538–545. [Google Scholar]
- Phalzgraff, T.; Borges, P.; Robaina, L.; Kaushok, S.; Izquierto, S. Essential fatty acid requirements of juvenile meagre (Argyrosomus regius). Aquaculture 2023, 572, 739532. [Google Scholar] [CrossRef]
- Makri, V.; Giantsis, I.A.; Nathanailides, C.; Feidantsis, K.; Antonopoulou, E.; Theodorou, J.A.; Michaelidis, B. Seasonal Energy Investment and Metabolic Patterns in a Farmed Fish. J. Therm. Biol. 2024, 123, 103894. [Google Scholar] [CrossRef]
- Carvalho, M.; Peres, H.; Saleh, R.; Fontanillas, R.; Rosenlund, G.; Oliva-Teles, A.; Izquierdo, M. Dietary Requirement for n-3 Long-Chain Polyunsaturated Fatty Acids for Fast Growth of Meagre (Argyrosomus regius, Asso 1801) Fingerlings. Aquaculture 2018, 488, 105–113. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Lutfi, E.; Sánchez-Moya, A.; Gutiérrez, J.; Capilla, E.; Navarro, I. Short-Term Responses to Fatty Acids on Lipid Metabolism and Adipogenesis in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2020, 21, 1623. [Google Scholar] [CrossRef]
- Omachi, D.O.; Aryee, A.N.; Onuh, J.O. Functional Lipids and Cardiovascular Disease Reduction: A Concise Review. Nutrients 2024, 16, 2453. [Google Scholar] [CrossRef]
- Das, P.; Dutta, A.; Panchali, T.; Khatun, A.; Kar, R.; Das, T.K.; Phoujdar, M.; Chakrabarti, S.; Ghosh, K.; Pradhan, S. Advances in Therapeutic Applications of Fish Oil: A Review. Meas. Food 2024, 13, 100142. [Google Scholar] [CrossRef]
- Goulas, A.E.; Kontominas, M.G. Effect of modified atmosphere packaging and vacuum packaging on the shelf-life of refrigerated chub mackerel (Scomber japonicus): Biochemical and sensory attributes. Eur. Food Res. Technol. 2007, 224, 545–553. [Google Scholar] [CrossRef]
- Secci, G.; Parisi, G. From Farm to Fork: Lipid Oxidation in Fish Products. A Review. Ital. J. Anim. Sci. 2016, 15, 124–136. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Costantini, D.; Cooke, S.J.; Willmore, W.G. A comparative and evolutionary approach to oxidative stress in fish: A review. Fish Fish. 2017, 18, 928–942. [Google Scholar] [CrossRef]
- Song, C.; Sun, C.; Liu, B.; Xu, P. Oxidative Stress in Aquatic Organisms. Antioxidants 2023, 12, 1223. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Jia, R.; Wang, L.; Hou, Y.; Feng, W.; Li, B.; Zhu, J. Effects of stocking density on the growth performance, physiological parameters, redox status and lipid metabolism of Micropterus salmoides in integrated rice–fish farming systems. Antioxidants 2022, 2022, 1215. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.L.; Han, C.; Huang, B.; Lei, J.L. Influence of Stocking Density on Growth Performance, Antioxidant Status, and Physiological Response of Juvenile Turbot, Scophthalmus maximu, Reared in Land-Based Recirculating Aquaculture System. J. World Aquac. Soc. 2016, 47, 587–599. [Google Scholar] [CrossRef]
- Jia, E.; Yan, Y.; Zhou, M.; Li, X.; Jiang, G.; Liu, W.; Zhang, D. Combined Effects of Dietary Quercetin and Resveratrol on Growth Performance, Antioxidant Capability and Innate Immunity of Blunt Snout Bream (Megalobrama amblycephala). Anim. Feed Sci. Technol. 2019, 256, 114268. [Google Scholar] [CrossRef]
- Diao, W.; Jia, R.; Hou, Y.; Dong, Y.; Li, B.; Zhu, J. (2023). Effects of stocking density on the growth performance, physiological parameters, antioxidant status and lipid metabolism of Pelteobagrus fulvidraco in the integrated rice-fish farming system. Animals 2023, 13, 1721. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, G.; Luan, Y.; Niu, J.; Li, Y.; Chen, H.; Liu, Y.; Zhu, S. Impact of Feeding Frequency on Growth Performance and Antioxidant Capacity of Litopenaeus vannamei in Recirculating Aquaculture Systems. Animals 2025, 15. [Google Scholar] [CrossRef]
- Mansour, A.T.; Goda, A.A.; Omar, E.A.; Khalil, H.S.; Esteban, M.Á. Dietary supplementation of organic selenium improves growth, survival, antioxidant and immune status of meagre, Argyrosomus regius, juveniles. Fish Shellfish. Immunol. 2017, 68, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, W.; Yin, Y.; Yang, L.; Chen, M.; Sun, Y.; Ye, J. Effects of Carnosine Addition in Low-Fishmeal Feed on the Growth Performance, Muscle Antioxidant Capacity and Flesh Quality of Orange-Spotted Grouper (Epinephelus coioides). Fishes 2024, 9, 467. [Google Scholar] [CrossRef]
- Harimanaa, Y.; Tanga, X.; Xuc, P.; Xuc, G.; Karangwab, E.; Zhangb, K.; Sun, Y.; Li, Y.; Ma, S.; Uriho, A.; et al. Effect of long-term moderate exercise on muscle cellularity and texture, antioxidant activities, tissue composition, freshness indicators and flavor characteristics in large mouth bass (Micropterus salmoides). Aquaculture 2019, 510, 100–108. [Google Scholar] [CrossRef]
- Berri, C.; Le Bihan-Duval, E.; Debut, M.; Santé-Lhoutellier, V.; Baéza, E.; Gigaud, V.; Jégo, Y.; Duclos, M.J. Consequence of Muscle Hypertrophy on Characteristics of Pectoralis Major Muscle and Breast Meat Quality of Broiler Chickens. J. Anim. Sci. 2007, 85, 2005–2011. [Google Scholar] [CrossRef]
- López-Albors, O.; Abdel, I.; Periago, M.J.; Ayala, M.D.; Alcázar, A.G.; Graciá, C.M.; Nathanailides, C.; Vázquez, J.M. Temperature Influence on the White Muscle Growth Dynamics of the Sea Bass Dicentrarchus labrax, L. Flesh Quality Implications at Commercial Size. Aquaculture 2008, 277, 39–51. [Google Scholar] [CrossRef]
- Strzelczak, A.; Balejko, J.; Szymczak, M.; Witczak, A. Effect of Protein Denaturation Temperature on Rheological Properties of Baltic Herring (Clupea harengus Membras) Muscle Tissue. Foods 2021, 10, 829. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Traits and Muscle Tissue Proximate Composition | SG | FG | p-Value |
|---|---|---|---|
| Body weight (BW, g) | 459.09 (21.01) | 1148.8 (215.32) | * |
| Gonadosomatic (GSI, %) | 0.03 (0.04) | 0.12 (0.05) | * |
| Hepatosomatic (HSI, %) | 1.2 (0.24) | 1.22 (0.25) | Nsd |
| Viscerosomatic (VSI, %) | 4.69 (2.01) | 4.78 (0.57) | Nsd |
| Perivisceral fat (PVF, %) | 2.08 (0.25) | 0.54 (0.33) | * |
| Filleting yield (FY, %) | 31.53 (3.16) | 33.46(3.53) | * |
| Moisture (%) | 78.29 (0.78) | 80.40(1.40) | * |
| Proteins (%) | 19.0 (0.44) | 18.92(0.61) | Nsd |
| Lipids (%) | 0.78 (0.05) | 0.27(0.04) | * |
| Ash (%) | 1.10 (0.02) | 1.02(0.04) | Nsd |
| Fatty Acid | Chemical Type | SG | FG | p-Value |
|---|---|---|---|---|
| Myristic acid | C14:0 | 2.91 (0.75) | 3.63 (1.03) | Nsd |
| Palmitic acid | C16:0 | 12.1 (1.54) | 16.54 (0.92) | * |
| Stearic acid | C18:0 | 4.19 (1.26) | 7.62 (0.79) | * |
| Palmitoleic acid | C16:1 (ω-7) | 0.21 (0.19) | 3.89 (0.20) | *** |
| cis-11-octadecenoic acid | C18:1 (ω-7) | 0.28 (0.25) | 0.63 (0.55) | Nsd |
| Oleic acid | C18:1 (ω-9) | 19.57 (1.48) | 18.88 (2.54) | Nsd |
| Linoleic acid (LA) | C18:2 (ω-6) | 9.83 (1.18) | 8.27 (2.10) | Nsd |
| Linoleni cacid (LNA) | C18:3 (ω-3) | 0.38 (0.11) | 0.97 (0.88 | Nsd |
| Eicosadienoic acid | C20:2 (ω-6) | 0.25 (0.21) | 0.82 (0.71) | Nsd |
| Dihomo-γ-linolenic acid (DGLH) | C20:3 (ω-6) | 0.19 (0.18) | 2.24 (0.46) | ** |
| Εicosatrienoic acid | C20:3 (ω-3) | 2.66 (0.52) | 4.37 (0.48) | * |
| Arachidonic acid | C20:4 (ω-6) | 3.43 (0.88) | 3.71 (1.77) | Nsd |
| Eicosapentaenoic acid (EPA) | C20:5 (ω-3) | 7.75 (8.20) | 3.69 (0.45) | Nsd |
| Docosahexaenoic acid (DHA) | C22:6 (ω-3) | 13.90 (3.31) | 7.47 (1.11) | * |
| Nervonic acid | C24:1 (ω-9) | 7.81 (0.79) | 7.25 (1.24) | Nsd |
| Lipid Quality Indices | SG | FG | p-Value |
|---|---|---|---|
| ω-3 | 24.30 (1.45) | 15.52 (1.26) | ** |
| ω-6 | 13.70 (1.39) | 15.04 (3.00) | nsd |
| ω-3/ω-6 | 1.78 (0.14) | 1.07 (0.30) | * |
| SFA | 19.21 (2.66) | 27.79 (0.95) | ** |
| ΣMUFA | 27.88 (1.72) | 29.86 (1.37) | nsd |
| ΣPUFA | 38.38 (31.53) | 31.53 (1.22) | * |
|
Myofiber Characteristics | SG | FG | p-Value |
|---|---|---|---|
| MCSA (μm2) | 1516.42 (±63.7) [0.0015 mm2] | 1667.44 (±448.9) [0.0017 mm2] [0.0017 mm2] | nsd |
| MFd (number/mm2) | 510.48(±131.8) | 449.68 (±106.8) | nsd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mittakos, I.; Nathanailides, C.I.; Kokokiris, L.E.; Barbouti, A.; Bitchava, K.; Gouva, E.; Kolygas, M.N.; Terzidis, M.A.; Kontominas, M.G. Antioxidant Capacity, Lipid Oxidation, and Quality Traits of Slow- and Fast-Growing Meagre (Argyrosomus regius) Fillets During Cold Storage. Antioxidants 2025, 14, 124. https://doi.org/10.3390/antiox14020124
Mittakos I, Nathanailides CI, Kokokiris LE, Barbouti A, Bitchava K, Gouva E, Kolygas MN, Terzidis MA, Kontominas MG. Antioxidant Capacity, Lipid Oxidation, and Quality Traits of Slow- and Fast-Growing Meagre (Argyrosomus regius) Fillets During Cold Storage. Antioxidants. 2025; 14(2):124. https://doi.org/10.3390/antiox14020124
Chicago/Turabian StyleMittakos, Ioannis, Cosmas I. Nathanailides, Lambros E. Kokokiris, Alexandra Barbouti, Konstantina Bitchava, Evangelia Gouva, Markos N. Kolygas, Michael A. Terzidis, and Michael G. Kontominas. 2025. "Antioxidant Capacity, Lipid Oxidation, and Quality Traits of Slow- and Fast-Growing Meagre (Argyrosomus regius) Fillets During Cold Storage" Antioxidants 14, no. 2: 124. https://doi.org/10.3390/antiox14020124
APA StyleMittakos, I., Nathanailides, C. I., Kokokiris, L. E., Barbouti, A., Bitchava, K., Gouva, E., Kolygas, M. N., Terzidis, M. A., & Kontominas, M. G. (2025). Antioxidant Capacity, Lipid Oxidation, and Quality Traits of Slow- and Fast-Growing Meagre (Argyrosomus regius) Fillets During Cold Storage. Antioxidants, 14(2), 124. https://doi.org/10.3390/antiox14020124