Influence of Postharvest Temperatures on Carotenoid Biosynthesis and Phytochemicals in Mature Green Chili (Capsicum annuum L.)

Abstract
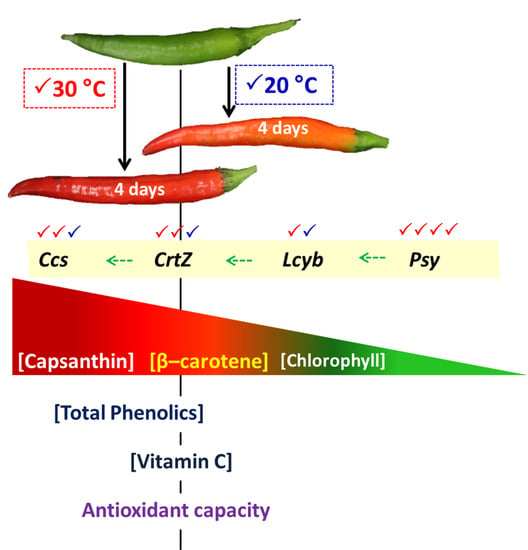
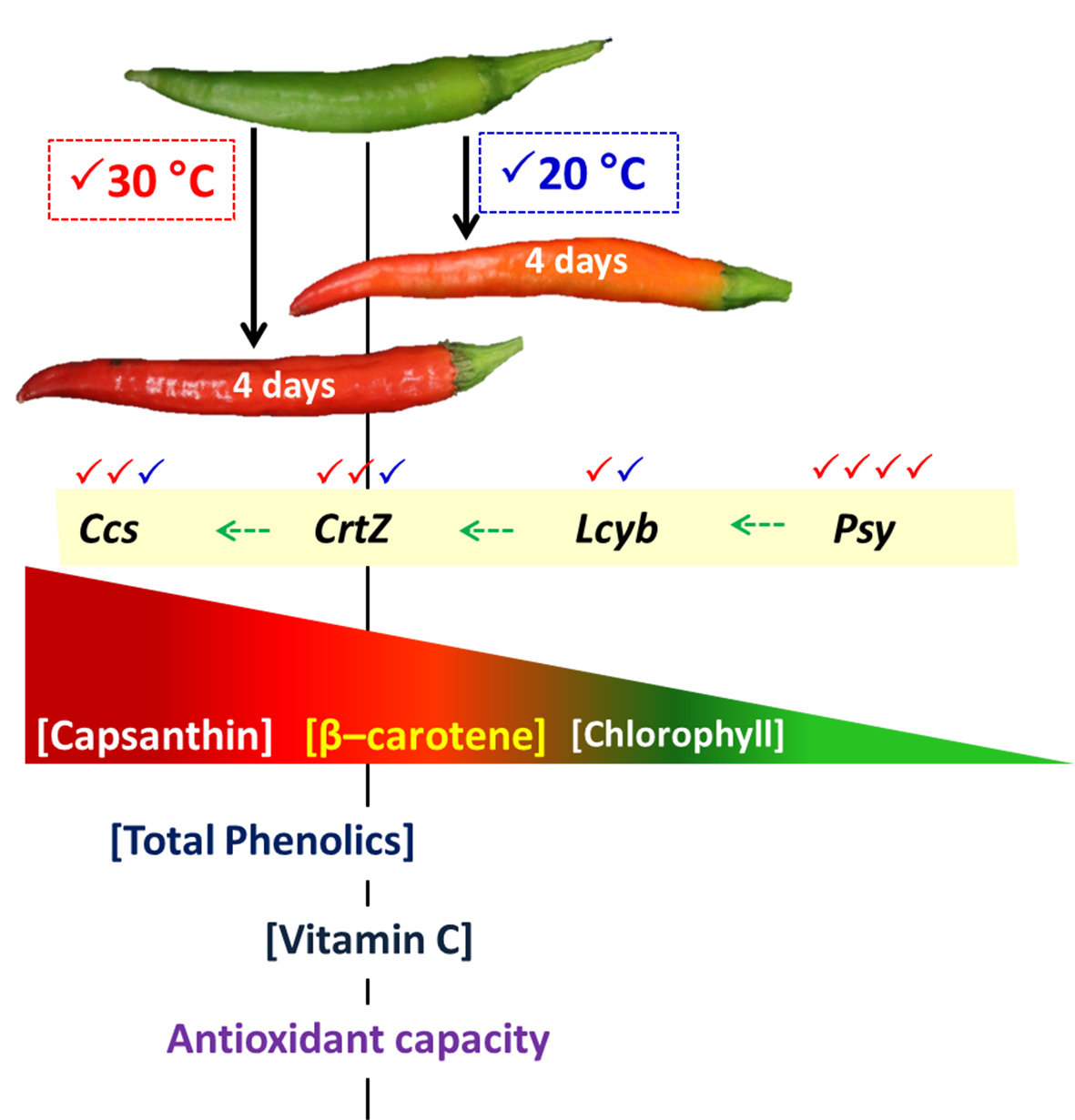
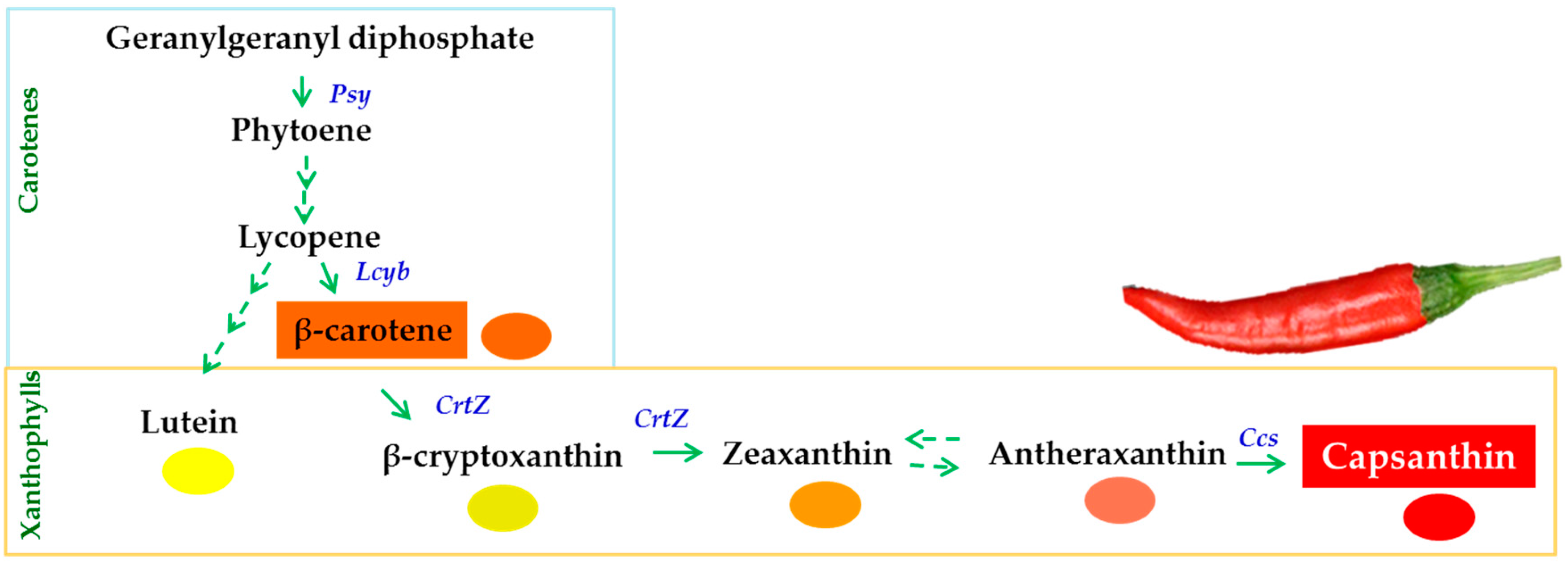
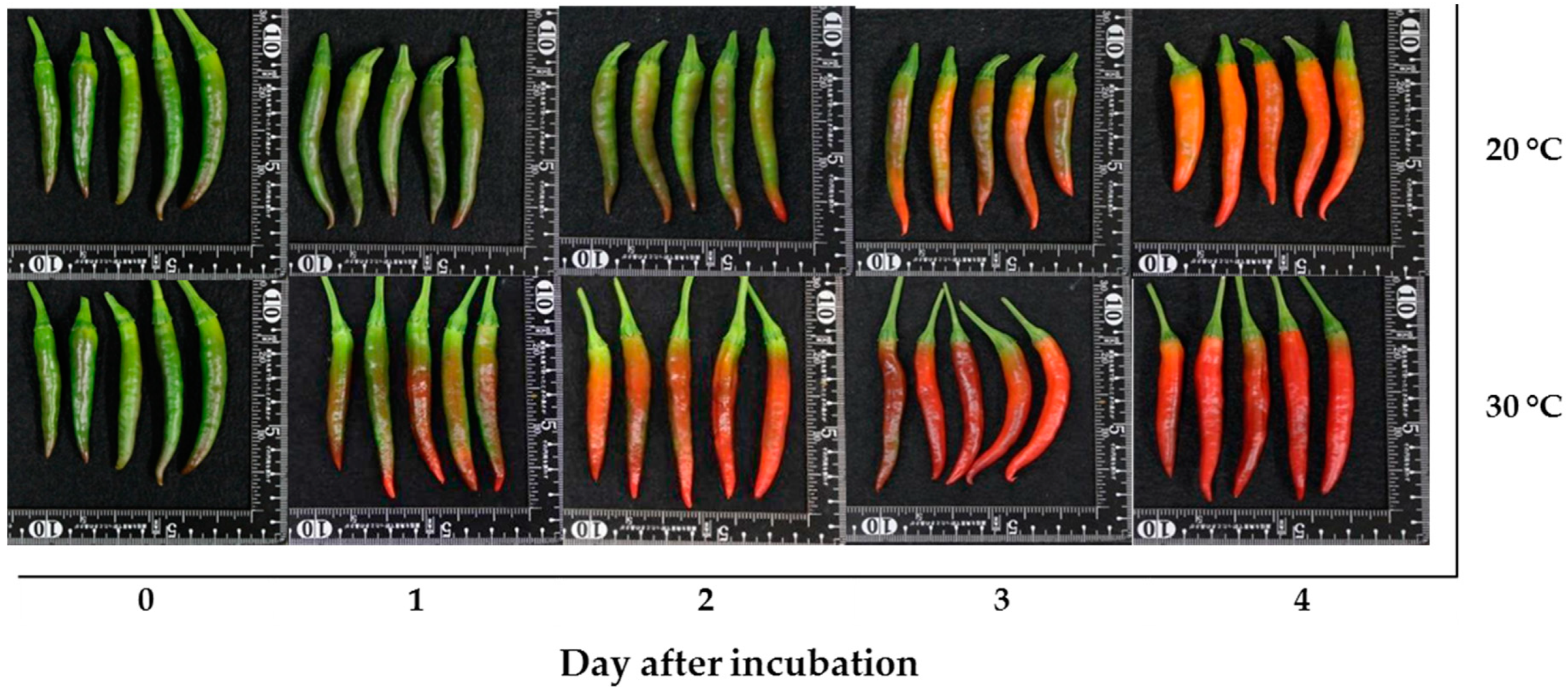
:
{kind=link}
{kind=link}
{kind=link}
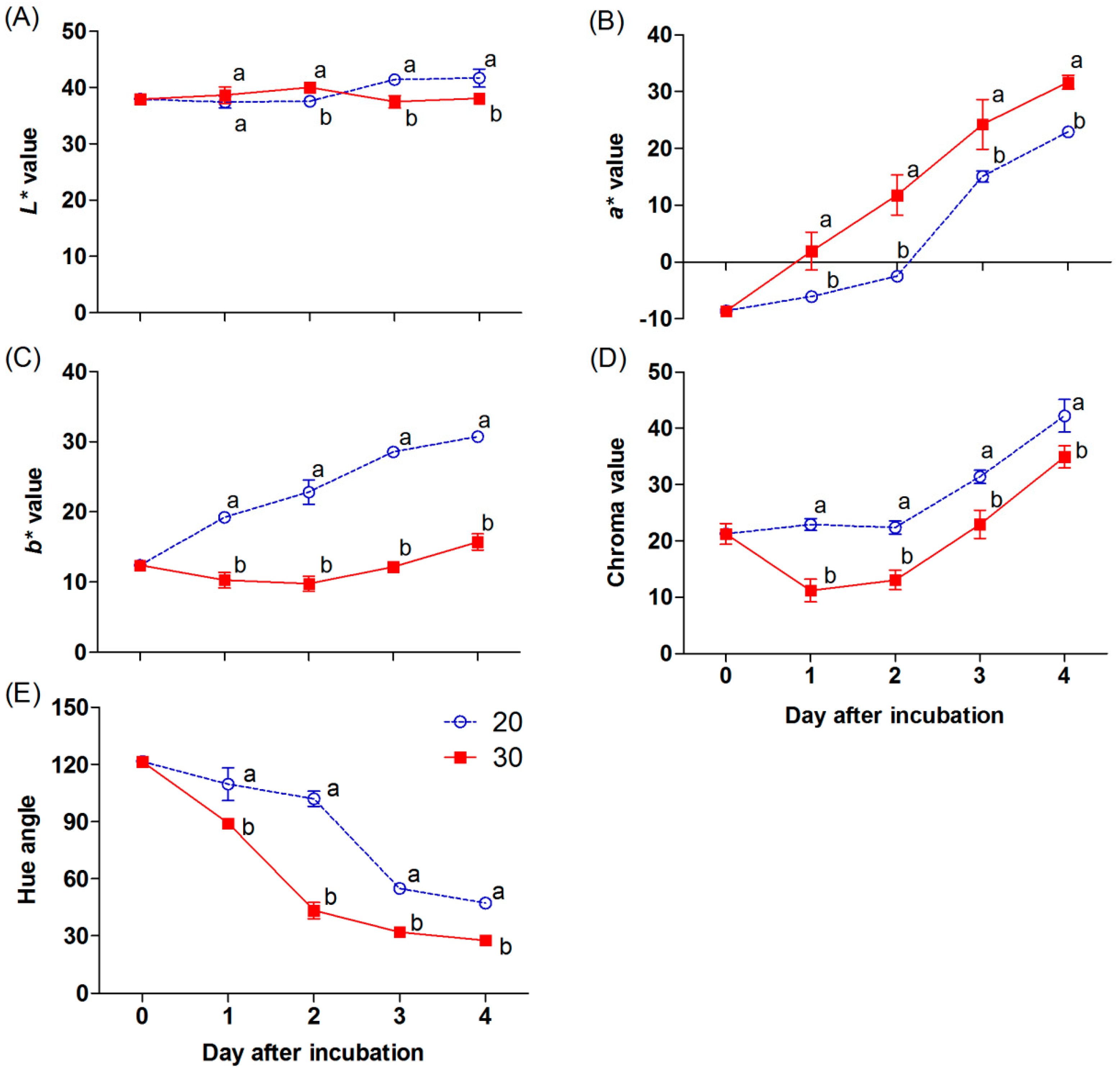{kind=link}
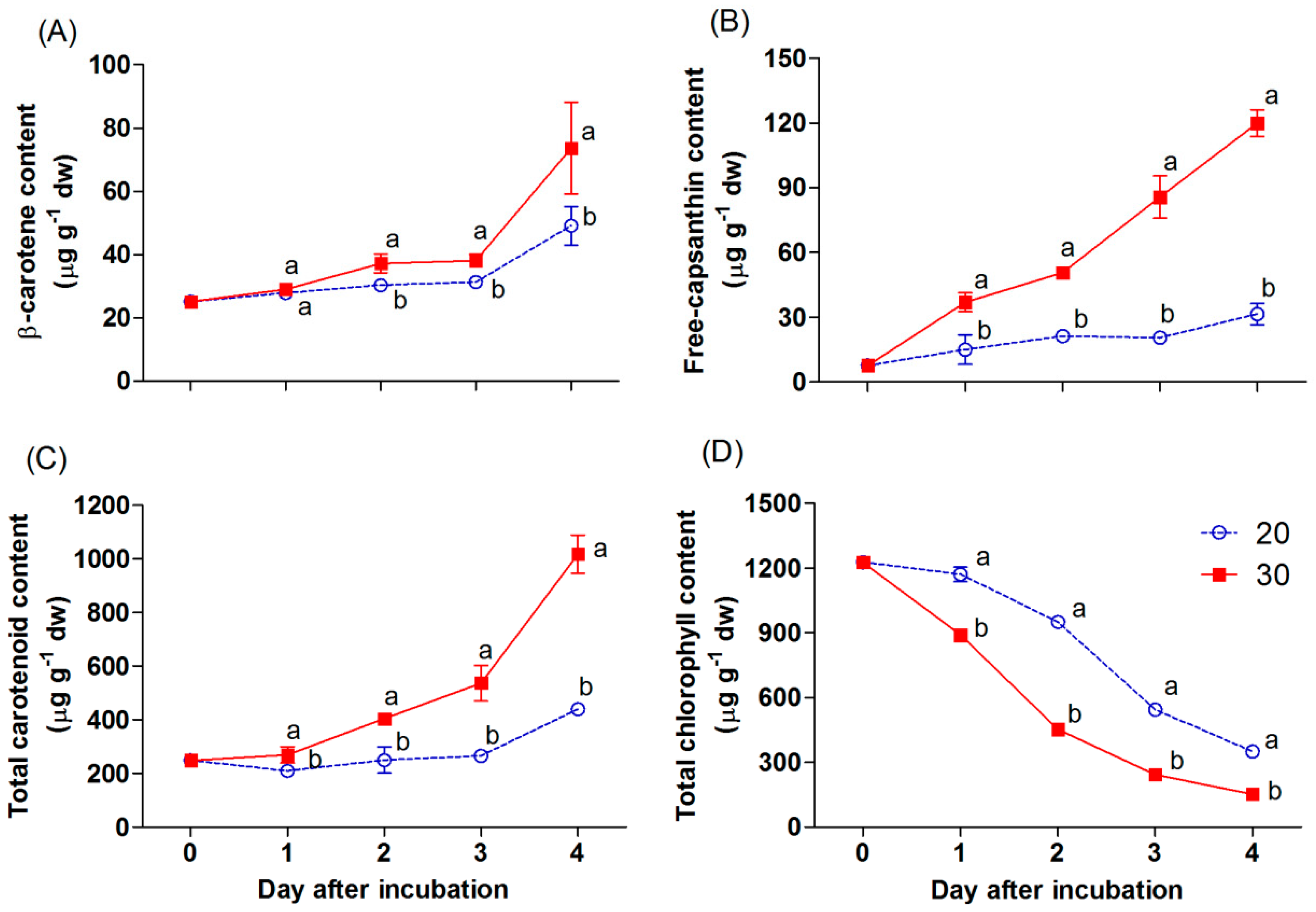{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
2.2. Surface Color Analysis
2.3. Total Phenolic Content (TPC) Determination
2.4. Vitamin C Content Determination
2.5. Antioxidant Capacity Determination
2.5.1. The 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
2.5.2. The Ferric-Reducing Antioxidant Power (FRAP) Assay
2.6. Pigments Determination
2.6.1. Total Chlorophyll Content
2.6.2. Carotenoid Content
2.7. Carotenoid Biosynthesis-Related Genes Expression Analysis
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Manikharda. Physicochemical properties, favor characteristics, and biological functions of the chili pepper Shimatogarashi (Capsicum frutescens). Ph.D. Thesis, The United Graduate School of Agricultural Sciences, Kagoshima University, Kagoshima, Japan, 26 April 2018.
- Acedo, A.L. International cooperator’s guide of postharvest technology for fresh chili pepper in Cambodia, Laos, and Vietnam. AVRDC World Veg. Cent. 2010, 10–735, 1–6. [Google Scholar]
- Menichini, F.; Tundis, R.; Bonesi, M.; Loizzo, M.R.; Conforti, F.; Statti, G.; Cindio, B.D.; Houghton, P.J.; Menichini, F. The influence of fruit ripening on the phytochemical content and biological activity of Capsicum chinense jacq. cv habanero. Food Chem. 2009, 114, 553–560. [Google Scholar] [CrossRef]
- Márkus, F.; Daood, H.G.; Kapitány, J.; Biass, P.A. Change in the carotenoid and antioxidant content of spice red pepper (paprika) as a function of ripening and some technological factors. J. Agric. Food Chem. 1999, 47, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Camara, B.; Brangeon, J. Carotenoid metabolism during chloroplast to chromoplast transformation in Capsicum annuum fruit. Planta 1981, 151, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.; Harel, S.; Palevitch, D.; Akiri, B.; Menagem, E.; Kanner, J. Carotenoid pigments and β-carotene in paprika fruits (Capsicum spp.) with different genotypes. J. Agric. Food Chem. 1995, 43, 362–366. [Google Scholar] [CrossRef]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction of paprika carotenoids, capsanthin and capsorubin, with reactive oxygen species. J. Agric. Food Chem. 2016, 64, 4786–4792. [Google Scholar] [CrossRef]
- Pola, W.; Sugaya, S.; Photchanachai, S. Color development and phytochemical changes in mature green chili (Capsicum annuum L.) exposed to red and blue light-emitting diodes. J. Agric. Food Chem. 2020, 68, 59–66. [Google Scholar] [CrossRef]
- Gómez-García, M.R.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 17, 19025–19053. [Google Scholar] [CrossRef] [Green Version]
- Thomus, P.; Janave, M.T. Effects of gamma irradiation and storage temperature on carotenoids and ascorbic acid content of mangoes on ripening. J. Sci. Food Agric. 1975, 26, 1503–1512. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ikoma, Y.; Kato, M.; Nakajima, N.; Hasegawa, Y. Effect of postharvest temperature and ethylene on carotenoid accumulation in the flavedo and juice sacs of Satsuma mandarin (Citrus unshiu Marc.) fruit. J. Agric. Food Chem. 2009, 57, 4724–4732. [Google Scholar] [CrossRef]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Kubo, Y. Ethylene, oxygen, carbon dioxide, and temperature in postharvest physiology. In Abiotic Stress Biology in Horticultural Plants; Kanayama, Y., Kochetov, A., Eds.; Springer: Tokyo, Japan, 2015; pp. 17–34. [Google Scholar]
- Yamauchi, N. Postharvest chlorophyll degradation and oxidative stress. In Abiotic Stress Biology in Horticultural Plants; Kanayama, Y., Kochetov, A., Eds.; Springer: Tokyo, Japan, 2015; pp. 101–113. [Google Scholar]
- Arnnok, P.; Ruangviriyachai, C.; Mahachai, R.; Techawongstien, S.; Chanthai, S. Determination of total phenolics and anthocyanin contents in the pericarp of hot chilli pepper (Capsicum annuum L.). Int. Food Res. J. 2012, 19, 235–243. [Google Scholar]
- Roe, J.H.; Milles, M.B.; Oesterling, M.J.; Damron, C.M. The determination of diketo-l-gulonic acid, dehydro-l-ascorbic acid and l-ascorbic acid in the same tissue extract by the 2,4-dinitrophenylhydrazine method. J. Biol. Chem. 1948, 174, 201–208. [Google Scholar] [PubMed]
- Arslan, D.; Özcan, M.M. Dehydration of red bell pepper (Capsicum annuum L.): Change in drying behavior, colour and antioxidant content. Food Bioprod. Process. 2011, 89, 504–513. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M.; Ornelas-Paz, J.J.; Victoria-Campos, C.I.; Ibarra-Junquera, V.; Pérez-Martínez, J.D.; Escalante-Minakata, P. Antioxidant activity and content of chlorophylls and carotenoids in raw and heat-processed Jalapeño peppers at intermediate stages of ripening. Food Chem. 2014, 146, 188–196. [Google Scholar] [CrossRef]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Welsch, R.; Medina, J.; Giuliano, G.; Beyer, P.; von Lintig, J. Structural and functional characterization of the phytoene synthase promoter from Arabidopsis thaliana. Planta 2003, 216, 523–534. [Google Scholar] [CrossRef]
- Pagamas, P.; Nawata, E. Sensitive stages of fruit and seed development of chili pepper (Capsicum annuum L. var. shishito) exposed to high-temperature stress. Sci. Hortic. 2008, 117, 21–25. [Google Scholar] [CrossRef]
- Tian, S.L.; Li, L.; Shah, S.N.M.; Gong, Z.H. The relationship between red fruit colour formation and key genes of capsanthin biosynthesis pathway in Capsicum annuum. Biol. Plant. 2015, 59, 507–513. [Google Scholar] [CrossRef]
- Deli, J.; Molnár, P.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of red paprika (Capsicum annuum var. lycopersiciforme rubrum) during ripening; biosynthesis of carotenoids in red paprika. J. Agric. Food Chem. 2001, 49, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.Q.; Yang, X.T.; Zhang, Z.Q. Chlorophyll degradation and its control in postharvest fruits. Stewart Postharvest Rev. 2008, 4, 1–4. [Google Scholar]
- Pinto, C.M.F.; dos Santos, I.C.; de Araujo, F.F.; da Silva, T.P. Pepper importance and growth (Capsicum spp.). In Production and Breeding of Chilli Peppers (Capsicum spp.); do Rêgo, E.R., do Rêgo, M.M., Finger, F.L., Eds.; Springer: Basel, Switzerland, 2016; pp. 1–26. [Google Scholar]
- Zhang, R.; Kramer, D.M.; Cruz, J.A.; Struck, K.R.; Sharkey, T.D. The effects of moderately high temperature on zeaxanthin accumulation and decay. Photosynth. Res. 2011, 108, 171–181. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.K.; Ali, Z.M.; Zainal, Z. Changes in ethylene production, carbohydrase activity and antioxidant status in pepper fruits during ripening. Sci. Hortic. 2012, 142, 23–31. [Google Scholar] [CrossRef]
- Sun, T.; Xu, Z.; Wu, C.T.; Janes, M.; Prinyawiwatkul, W.; No, H.K. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.). J. Food Sci. 2007, 72, S98–S102. [Google Scholar] [CrossRef]
- Mateos, R.M.; Jiménez, A.; Román, P.; Romojaro, F.; Bacarizo, S.; Leterrier, M.; Gómez, M.; Sevilla, F.; Del Río, L.A.; Corpas, F.J.; et al. Antioxidant systems from pepper (Capsicum annuum L.): Involvement in the response to temperature changes in ripe fruits. Int. J. Mol. Sci. 2013, 14, 9556–9580. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pola, W.; Sugaya, S.; Photchanachai, S. Influence of Postharvest Temperatures on Carotenoid Biosynthesis and Phytochemicals in Mature Green Chili (Capsicum annuum L.). Antioxidants 2020, 9, 203. https://doi.org/10.3390/antiox9030203
Pola W, Sugaya S, Photchanachai S. Influence of Postharvest Temperatures on Carotenoid Biosynthesis and Phytochemicals in Mature Green Chili (Capsicum annuum L.). Antioxidants. 2020; 9(3):203. https://doi.org/10.3390/antiox9030203
Chicago/Turabian StylePola, Wissanee, Sumiko Sugaya, and Songsin Photchanachai. 2020. "Influence of Postharvest Temperatures on Carotenoid Biosynthesis and Phytochemicals in Mature Green Chili (Capsicum annuum L.)" Antioxidants 9, no. 3: 203. https://doi.org/10.3390/antiox9030203
APA StylePola, W., Sugaya, S., & Photchanachai, S. (2020). Influence of Postharvest Temperatures on Carotenoid Biosynthesis and Phytochemicals in Mature Green Chili (Capsicum annuum L.). Antioxidants, 9(3), 203. https://doi.org/10.3390/antiox9030203