Oxidative Status of Goats with Different CSN1S1 Genotypes Fed ad Libitum with Fresh and Dry Forages

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection and Assay
2.3. Statistical Analysis
3. Results
3.1. Redox Balance
3.2. Enzymatic Activity
3.3. Metabolic Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Celi, P.; Di Trana, A.; Claps, S. Effects of plane of nutrition on oxidative stress in goats during the peripartum period. Vet. J. 2010, 184, 95–99. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Wang, Y.; Chun, O.K.; Song, W.O. Plasma and dietary antioxidant status as cardiovascular disease risk factors: A review of human studies. Nutrients 2013, 5, 2969–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. Redox Biology in Transition Periods of Dairy Cattle: Role in the Health of Periparturient and Neonatal Animals. Antioxidants 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliotti, L.; Lai, O.; Cavallina, R.; Alfieri, L.; Rabusca, G.; Benvenuti, M.N. Changes in physiological and immunological parameters during the peripartum period in Zerasca sheep. Pol. J. Vet. Sci. 2014, 17, 737–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecchini, S.; Piccione, G.; Saoca, C.; Giangrosso, G.; Caputo, A.R.; Fazio, F. Redox status, lipid peroxidation and protein oxidation levels in small ruminants. J. Anim. Feed Sci. 2018, 27, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Ognik, K.; Patkowski, K.; Gruszecki, T.; Kostro, K. Redox status in the blood of ewes in the perinatal period and during lactation. Bull. Vet. Inst. Pulawy 2015, 59, 557–561. [Google Scholar] [CrossRef] [Green Version]
- Celi, P.; Di Trana, A.; Quaranta, A. Metabolic profile and oxidative status in goats during the peripartum period. Aust. J. Exp. Agric. 2008, 48, 1004–1008. [Google Scholar] [CrossRef]
- Karapehlivan, M.; Kaya, I.; Sag, A.; Akin, S.; Ozcan, A. Effects of Early and Late Lactation Period on Plasma Oxidant/Antioxidant Balance of Goats. Kafkas Univ. Vet. Fak. Derg. 2013, 19, 529–533. [Google Scholar] [CrossRef]
- Radin, L.; Simpraga, M.; Vince, S.; Kostelic, A.; Milinkovic-Tur, S. Metabolic and oxidative status of Saanen goats of different parity during the peripartum period. J. Dairy Res. 2015, 82, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Bharti, V.K.; Giri, A.; Vivek, P.; Kalia, S. Health and productivity of dairy cattle in high altitude cold desert environment of Leh-Ladakh: A review. Ind. J. Anim. Sci. 2017, 87, 3–10. [Google Scholar]
- Silanikove, N.; Koluman, N. Impact of climate change on the dairy industry in temperate zones: Predications on the overall negative impact and on the positive role of dairy goats in adaptation to earth warming. Small Rumin. Res. 2015, 123, 27–34. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Rathwa, S.D.; Vasava, A.A.; Pathan, M.M.; Madhira, S.P.; Patel, Y.G.; Pande, A.M. Effect of season on physiological, biochemical, hormonal, and oxidative stress parameters of indigenous sheep. Vet. World 2017, 10, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Di Trana, A.; Celi, P.; Claps, S.; Fedele, V.; Rubino, R. The effect of hot season and nutrition on the oxidative status and metabolic profile in dairy goats during mid lactation. Anim. Sci. 2006, 82, 717–722. [Google Scholar] [CrossRef]
- Gabai, G.; Testoni, S.; Piccinini, R.; Marinelli, L.; Stradaioli, G. Oxidative stress in primiparous cows in relation to dietary starch and the progress of lactation. Anim. Sci. 2004, 79, 99–108. [Google Scholar] [CrossRef]
- Sgorlon, S.; Stradaioli, G.; Gabai, G.; Stefanon, B. Variation of starch and fat in the diet affects metabolic status and oxidative stress in ewes. Small Rumin. Res. 2008, 74, 123–129. [Google Scholar] [CrossRef]
- Di Grigoli, A.; Di Trana, A.; Alabiso, M.; Maniaci, G.; Giorgio, D.; Bonanno, A. Effects of Grazing on the Behaviour, Oxidative and Immune Status, and Production of Organic Dairy Cows. Animals 2019, 9, 371. [Google Scholar] [CrossRef] [Green Version]
- Tsiplakou, E.; Mitsiopoulou, C.; Mavrommatis, A.; Karaiskou, C.; Chronopoulou, E.G.; Mavridis, G.; Sotirakoglou, K.; Labrou, N.E.; Zervas, G. Effect of under- and overfeeding on sheep and goat milk and plasma enzymes activities related to oxidation. J. Anim. Physiol. Anim. Nutr. 2018, 102, E288–E298. [Google Scholar] [CrossRef]
- Oh, J.; Wall, E.H.; Bravo, D.M.; Hristov, A.N. Host-mediated effects of phytonutrients in ruminants: A review. J. Dairy Sci. 2017, 100, 5974–5983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colitti, M.; Stefanon, B.; Gabai, G.; Gelain, M.E.; Bonsembiante, F. Oxidative Stress and Nutraceuticals in the Modulation of the Immune Function: Current Knowledge in Animals of Veterinary Interest. Antioxidants 2019, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, T.; Tan, B.; Yin, Y.L.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanon, B.; Sgorlon, S.; Gabai, G. Usefulness of nutraceutics in controlling oxidative stress in dairy cows around parturition. Vet. Res. Commun. 2005, 29, 387–390. [Google Scholar] [CrossRef]
- Yesilbag, D.; Biricik, H.; Cetin, I.; Kara, C.; Meral, Y.; Cengiz, S.S.; Orman, A.; Udum, D. Effects of juniper essential oil on growth performance, some rumen protozoa, rumen fermentation and antioxidant blood enzyme parameters of growing Saanen kids. J. Anim. Physiol. Anim. Nutr. 2017, 101, e67–e76. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Todaro, M.; Alabiso, M.; Vitale, F.; Di Trana, A.; Giorgio, D.; Settanni, L.; Gaglio, R.; Laddomada, B.; et al. Improvement of Oxidative Status, Milk and Cheese Production, and Food Sustainability Indexes by Addition of Durum Wheat Bran to Dairy Cows’ Diet. Animals 2019, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Ollier, S.; Chauvet, S.; Martin, P.; Chilliard, Y.; Leroux, C. Goat’s alpha(S1)-casein polymorphism affects gene expression profile of lactating mammary gland. Animal 2008, 2, 566–573. [Google Scholar] [CrossRef]
- Pagano, R.I.; Pennisi, P.; Valenti, B.; Lanza, M.; Di Trana, A.; Di Gregorio, P.; De Angelis, A.; Avondo, M. Effect of CSN1S1 genotype and its interaction with diet energy level on milk production and quality in Girgentana goats fed ad libitum. J. Dairy Res. 2010, 77, 245–251. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Di Trana, A.; Di Gregorio, P.; Tornambe, G.; Bellina, V.; Claps, S.; Maggio, G.; Todaro, M. Influence of fresh forage-based diets and alphas(1)-casein (CSN1S1) genotype on nutrient intake and productive, metabolic, and hormonal responses in milking goats. J. Dairy Sci. 2013, 96, 2107–2117. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Montalbano, M.; Bellina, V.; Mazza, F.; Todaro, M. Effects of diet on casein and fatty acid profiles of milk from goats differing in genotype for α S1-casein synthesis. Eur. Food Res. Technol. 2013, 237, 951–963. [Google Scholar] [CrossRef] [Green Version]
- Valenti, B.; Pagano, R.I.; Avondo, M. Effect of diet at different energy levels on milk casein composition of Girgentana goats differing in CSN1S1 genotype. Small Rumin. Res. 2010, 105, 135–139. [Google Scholar] [CrossRef]
- Di Trana, A.; Di Gregorio, P.; Maggio, G.; Di Napoli, M.A.; Claps, S.; Giorgio, D. Effect of genotype at CSN1S1 per diet interaction on milk fatty acid profile in Derivata di Siria lactating goats. In Proceedings of the XI INTERNATIONAL CONFERENCE ON GOATS, Gran Canaria, Spain, 23–27 September 2012; p. 241. [Google Scholar]
- Bowling, A.T.; Stott, M.L.; Bickel, L. Silent Blood Chimerism in a Mare Confirmed by DNA Marker Analysis of Hair Bulbs. Anim. Genet. 1993, 24, 323–324. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, G.; Illario, R.; Rando, A.; di Gregorio, P.; Masina, P.; Ramunno, L. Molecular characterization of the goat CSN1S1(01) allele. J. Dairy Res. 2003, 70, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.J.; Leroux, C.; Bonastre, A.S.; Martin, P. Occurrence of a LINE sequence in the 3’ UTR of the goat alpha s1-casein E-encoding allele associated with reduced protein synthesis level. Gene 1994, 147, 179–187. [Google Scholar] [CrossRef]
- Ramunno, L.; Cosenza, G.; Pappalardo, M.; Pastore, N.; Gallo, D.; Di Gregorio, P.; Masina, P. Identification of the goat CSN1S1F allele by means of PCR-RFLP method. Anim Genet. 2000, 31, 342–343. [Google Scholar] [CrossRef]
- Ramunno, L.; Cosenza, G.; Pappalardo, M.; Longobardi, E.; Gallo, D.; Pastore, N.; Di Gregorio, P.; Rando, A. Characterization of two new alleles at the goat CSN1S2 locus. Anim Genet. 2001, 32, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Ramunno, L.; Mariani, P.; Pappalardo, M.; Rando, A.; Capuano, M.; Di Gregorio, P.; Cosenza, G. A major-effect gene for β-casein content in goat milk. In Proceedings of the Atti XI Congresso Nazionale ASPA, Grado, Italy, 19–22 June 1995; pp. 19–22. [Google Scholar]
- Paris Institut National de la Recherche, A. Alimentation Des Bovins, Ovins & Caprins; INRA: Paris, France, 1988. [Google Scholar]
- Cesarone, M.R.; Belcaro, G.; Carratelli, M.; Cornelli, U.; De Sanctis, M.T.; Incandela, L.; Barsotti, A.; Terranova, R.; Nicolaides, A. A simple test to monitor oxidative stress. Int. Angiol. 1999, 18, 127–130. [Google Scholar]
- Alberti, A.; Bolognini, L.; Macciantelli, D.; Caratelli, M. The radical cation of N,N-diethyl-para-paraphenylendiamine: A possible indicator of oxidative stress in biological samples. Res. Chem. Intermed. 2000, 26, 253–267. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. Oxidative stress index (OSi) as a new tool to assess redox status in dairy cattle during the transition period. Animal 2013, 7, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Ranade, R.; Talukder, S.; Muscatello, G.; Celi, P. Assessment of oxidative stress biomarkers in exhaled breath condensate and blood of dairy heifer calves from birth to weaning. Vet. J. 2014, 202, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Trinder, P. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis. Methods 925.10, 65.17, 974.24, 992.16, 17th ed.; Association of Official Analytical Chemists International Gaithersburg: Rockville, MD, USA, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Tava, A.; De Benedetto, M.; Tedesco, D.; Di Miceli, G.; Piluzza, G. Proanthocyanidins from Hedysarum, Lotus and Onobrychis spp. growing in Sardinia and Sicily and their antioxidant activity. In Proceedings of the XX International Grassland Congress: Offered Papers, Dublin, Ireland, 26 June–1 July 2005. [Google Scholar]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Barbato, O.; Holmes, B.; Filipescu, I.-E.; Celi, P. Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows. Antioxidants 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Ciliberti, M.; Soccio, M.; Pastore, D.; Albenzio, M.; Sevi, A.; Caroprese, M. Antioxidant/oxidant balance: Application as a biomarker of the antioxidant status in plasma of ewes fed seaweed Ascophyllum nodosum and flaxseed under high ambient temperature. Small Rumin. Res. 2019, 170, 102–108. [Google Scholar] [CrossRef]
- Piluzza, G.; Bullitta, S. The dynamics of phenolic concentration in some pasture species and implications for animal husbandry. J. Sci. Food Agric. 2010, 90, 1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Riedl, K.M.; Jones, G.A.; Sovik, K.N.; Ritchard, N.T.; Hartzfeld, P.W.; Riechel, T.L. High molecular weight plant polyphenolics (tannins) as biological antioxidants. J. Agric. Food Chem. 1998, 46, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Riedl, K.M.; Hagerman, A.E. Tannin− protein complexes as radical scavengers and radical sinks. J. Agric. Food Chem. 2001, 49, 4917–4923. [Google Scholar] [CrossRef]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Kapusta, A.; Kuczyska, B.; Puppel, K.; Kamaszewski, M. The relationship between early stages of lactation and antioxidant capacity of milk and blood plasma of PHF cows. Anim. Sci. Pap. Rep. 2018, 36, 149–158. [Google Scholar]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Sgorlon, S.; Stradaioli, G.; Zanin, D.; Stefanon, B. Biochemical and molecular responses to antioxidant supplementation in sheep. Small Rumin. Res. 2006, 64, 143–151. [Google Scholar] [CrossRef]
- Jiang, Z.; Wan, Y.; Li, P.; Xue, Y.; Cui, W.; Chen, Q.; Chen, J.; Wang, F.; Mao, D. Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep. Animals 2019, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Van Zanden, J.J.; Geraets, L.; Wortelboer, H.M.; van Bladeren, P.J.; Rietjens, I.M.; Cnubben, N.H. Structural requirements for the flavonoid-mediated modulation of glutathione S-transferase P1-1 and GS-X pump activity in MCF7 breast cancer cells. Biochem. Pharmacol. 2004, 67, 1607–1617. [Google Scholar] [CrossRef]
- Fukao, T.; Hosono, T.; Misawa, S.; Seki, T.; Ariga, T. The effects of allyl sulfides on the induction of phase II detoxification enzymes and liver injury by carbon tetrachloride. Food Chem. Toxicol. 2004, 42, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Mudron, P.; Rehage, J.; Qualmann, K.; Sallmann, H.P.; Scholz, H. A study of lipid peroxidation and vitamin E in dairy cows with hepatic insufficiency. J. Vet. Med. Ser. A 1999, 46, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Dunshea, F.; Bell, A.; Trigg, T. Relations between plasma non-esterified fatty acid metabolism and body fat mobilization in primiparous lactating goats. Br. J. Nutr. 1989, 62, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Latruffe, N.; Menzel, M.; Delmas, D.; Buchet, R.; Lançon, A. Compared binding properties between resveratrol and other polyphenols to plasmatic albumin: Consequences for the health protecting effect of dietary plant microcomponents. Molecules 2014, 19, 17066–17077. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Diet | |||||
|---|---|---|---|---|---|
| DAF100 | DAF65 | FSF100 | FSF65 | ||
| Dry matter | % | 87.5 | 85.7 | 19.1 | 47.1 |
| Crude protein | % DM 1 | 15.0 | 15.1 | 17.2 | 15.5 |
| Ether extract | % DM | 2.1 | 2.3 | 3.2 | 2.8 |
| NDF 2 | % DM | 54.2 | 44.3 | 32.5 | 26.7 |
| ADF 3 | % DM | 36.8 | 24.6 | 26.9 | 19.4 |
| ADL 4 | % DM | 12.9 | 6.7 | 4.9 | 3.5 |
| Ash | % DM | 11.3 | 10.4 | 9.6 | 6.71 |
| NFC 5 | % DM | 17.4 | 27.9 | 37.5 | 48.3 |
| UFL 6 | n/kg DM | 0.65 | 0.81 | 0.77 | 0.98 |
| Item | Diet (D) | SEM | Genotype (G) | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DAF100 | DAF65 | FSF100 | FSF65 | AA | AF | D | G | D × G | |||
| ROMs | 240.34 A | 260.80 A | 75.60 B | 71.79 B | 23.12 | 165.85 | 158.42 | 16.41 | 0.001 | 0.750 | 0.762 |
| BAP | 7.66 b | 7.94 a | 7.96 a | 8.01 a | 0.097 | 7.91 | 7.87 | 0.069 | 0.050 | 0.647 | 0.684 |
| OSI | 12.01 A | 9.92 A | 2.87 B | 2.44 B | 1.413 | 7.29 | 6.33 | 1.003 | 0.001 | 0.502 | 0.720 |
| SOD | 5.09 b | 5.35 a | 5.47 a | 5.37 a | 0.080 | 5.32 | 5.33 | 0.057 | 0.040 | 0.935 | 0.617 |
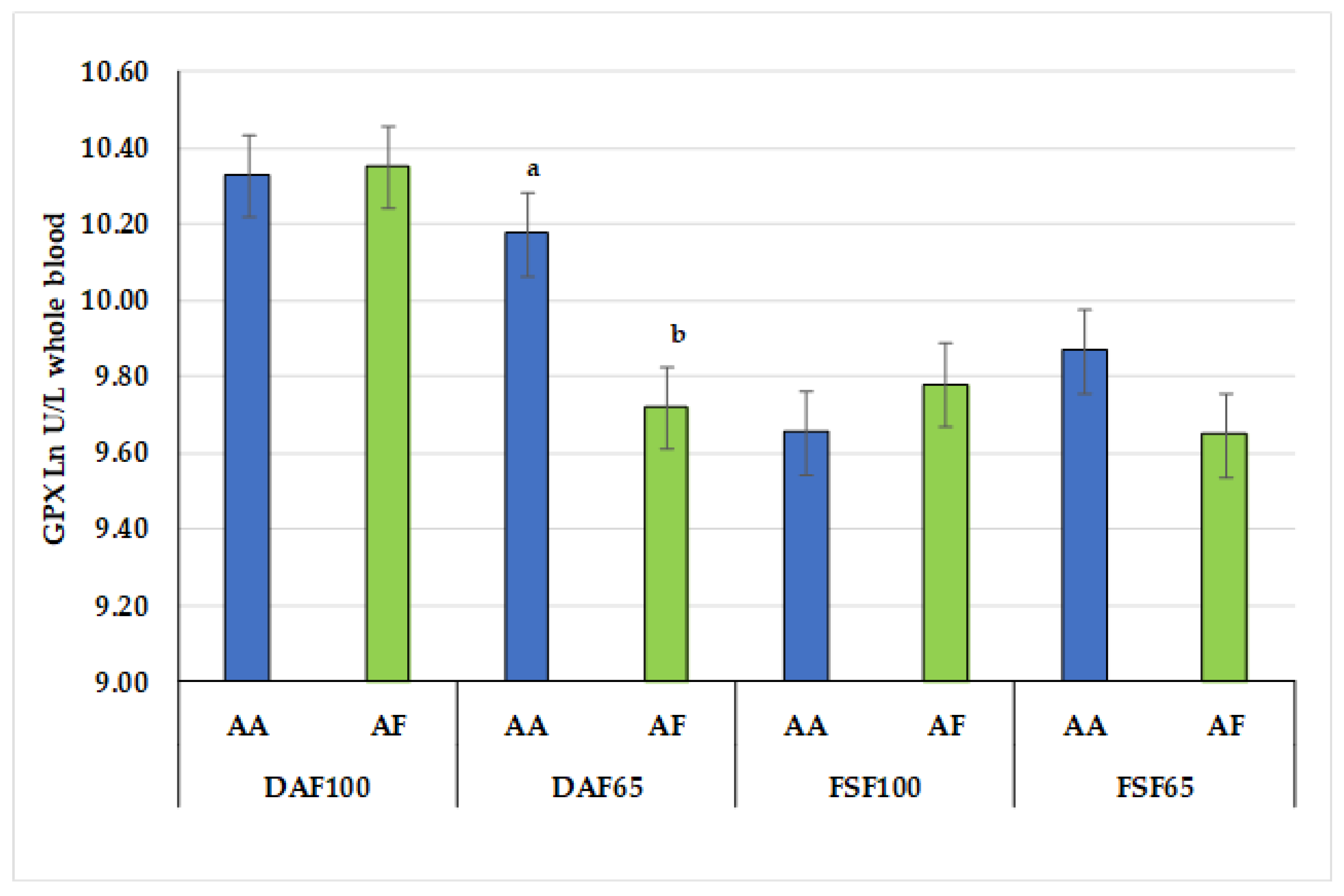
| GPX | 10.34 a | 9.95 b | 9.72 c | 9.76 c | 0.073 | 10.01 $ | 9.88 £ | 0.055 | 0.001 | 0.104 | 0.036 |
| Glucose | 51.03 | 49.59 | 51.63 | 53.00 | 1.56 | 50.79 | 50.83 | 1.11 | 0.093 | 0.977 | 0.443 |
| NEFA | 0.163 | 0.133 | 0.262 | 0.176 | 0.053 | 0.164 | 0.203 | 0.037 | 0.397 | 0.471 | 0.685 |
| BHBA | 0.377 | 0.340 | 0.287 | 0.512 | 0.064 | 0.425 | 0.333 | 0.045 | 0.150 | 0.166 | 0.296 |
| Cholesterol | 66.72 | 67.96 | 67.04 | 68.29 | 4.52 | 65.30 | 69.71 | 3.21 | 0.994 | 0.338 | 0.866 |
| Albumin | 3.31 b | 3.32 b | 3.86 a | 3.85 a | 0.02 | 3.58 | 3.59 | 0.01 | 0.001 | 0.665 | 0.395 |
| ROMs | BAP | SOD | Albumin | |
|---|---|---|---|---|
| GPX | 0.541 ** | −0.280 * | −0.219 * | −0.475 ** |
| SOD | −0.303 * | |||
| Albumin | −0.796 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorgio, D.; Di Trana, A.; Di Gregorio, P.; Rando, A.; Avondo, M.; Bonanno, A.; Valenti, B.; Di Grigoli, A. Oxidative Status of Goats with Different CSN1S1 Genotypes Fed ad Libitum with Fresh and Dry Forages. Antioxidants 2020, 9, 224. https://doi.org/10.3390/antiox9030224
Giorgio D, Di Trana A, Di Gregorio P, Rando A, Avondo M, Bonanno A, Valenti B, Di Grigoli A. Oxidative Status of Goats with Different CSN1S1 Genotypes Fed ad Libitum with Fresh and Dry Forages. Antioxidants. 2020; 9(3):224. https://doi.org/10.3390/antiox9030224
Chicago/Turabian StyleGiorgio, Daniela, Adriana Di Trana, Paola Di Gregorio, Andrea Rando, Marcella Avondo, Adriana Bonanno, Bernardo Valenti, and Antonino Di Grigoli. 2020. "Oxidative Status of Goats with Different CSN1S1 Genotypes Fed ad Libitum with Fresh and Dry Forages" Antioxidants 9, no. 3: 224. https://doi.org/10.3390/antiox9030224
APA StyleGiorgio, D., Di Trana, A., Di Gregorio, P., Rando, A., Avondo, M., Bonanno, A., Valenti, B., & Di Grigoli, A. (2020). Oxidative Status of Goats with Different CSN1S1 Genotypes Fed ad Libitum with Fresh and Dry Forages. Antioxidants, 9(3), 224. https://doi.org/10.3390/antiox9030224