Abstract
Global climate change and associated adverse abiotic stress conditions, such as drought, salinity, heavy metals, waterlogging, extreme temperatures, oxygen deprivation, etc., greatly influence plant growth and development, ultimately affecting crop yield and quality, as well as agricultural sustainability in general. Plant cells produce oxygen radicals and their derivatives, so-called reactive oxygen species (ROS), during various processes associated with abiotic stress. Moreover, the generation of ROS is a fundamental process in higher plants and employs to transmit cellular signaling information in response to the changing environmental conditions. One of the most crucial consequences of abiotic stress is the disturbance of the equilibrium between the generation of ROS and antioxidant defense systems triggering the excessive accumulation of ROS and inducing oxidative stress in plants. Notably, the equilibrium between the detoxification and generation of ROS is maintained by both enzymatic and nonenzymatic antioxidant defense systems under harsh environmental stresses. Although this field of research has attracted massive interest, it largely remains unexplored, and our understanding of ROS signaling remains poorly understood. In this review, we have documented the recent advancement illustrating the harmful effects of ROS, antioxidant defense system involved in ROS detoxification under different abiotic stresses, and molecular cross-talk with other important signal molecules such as reactive nitrogen, sulfur, and carbonyl species. In addition, state-of-the-art molecular approaches of ROS-mediated improvement in plant antioxidant defense during the acclimation process against abiotic stresses have also been discussed.
1. Introduction
Environmental stresses, including salinity, drought, extreme temperature, toxic metals/metalloids, flooding/waterlogging (WL), etc. are now prevalent due to drastic and harsh climate change [1,2]. The aggravation of such diverse abiotic stresses has become a major threat to sustainable crop production. Alongside, numerous detrimental effects lead to oxidative stress through the overaccumulation of reactive oxygen species (ROS) including free radicals (superoxide anion, O2•−; hydroperoxyl radical, HO2•; alkoxy radical, RO•; and hydroxyl radical, •OH) and nonradical molecules (hydrogen peroxide, H2O2 and singlet oxygen, 1O2) [3,4]. High-energy initiation or electron transfer reactions lead to atmospheric oxygen (O2) to the abovementioned partially reduced or activated forms of molecular oxygen [5]. The primary cellular ROS generation sites are chloroplasts, mitochondria, peroxisomes, apoplast, and plasma membranes [6]. Although ROS are formed in the plant as part of normal cellular metabolism, overaccumulation due to stress severely damages necessary cellular ingredients including carbohydrates, proteins, lipids, DNA, etc. because of their highly reactive nature [7].
Plants primarily deal with oxidative stress via an endogenous defensive mechanism consisting of different enzymatic (superoxide dismutase, SOD; catalase, CAT; ascorbate peroxidase, APX; glutathione reductase, GR; monodehydroascorbate reductase, MDHAR; dehydroascorbate reductase, DHAR; glutathione peroxidase, GPX; guaiacol peroxidase, GOPX; glutathione S-transferase, GST; Ferritin; nicotinamide adenine dinucleotide phosphate (NADPH) oxidase-like alternative oxidase, AOX; peroxiredoxins, PRXs; thioredoxins, TRXs; glutaredoxin, GRX; etc.) and nonenzymatic (ascorbic acid, AsA; glutathione, GSH; phenolic acids; alkaloids; flavonoids; carotenoids; α-tocopherol; nonprotein amino acids; etc.) antioxidants [8,9,10]. In plant cells, the antioxidant defense system and ROS accumulation uphold a steady-state balance [9]. Maintaining an optimum ROS level in the cell enables proper redox biology reactions and the regulation of numerous processes essential for plants such as growth and development [11]. This intermediate level is maintained by the balance between ROS production and ROS scavenging [4]. However, during stress conditions, overgeneration of ROS demolishes the equilibrium and causes cellular damage, leading to programmed cell death (PCD) as well as decreasing plant productivity [7].
Besides their damaging activity, ROS are well known as secondary messengers or signaling molecules and transport the signal to the nucleus through redox reactions using mitogen-activated protein kinase (MAPK) pathway in a variety of cellular mechanisms to increase tolerance against diverse abiotic stresses [6]. Reactive oxygen species participate as major molecules in the acclimation process of plants under environmental stimuli. They principally act as signal transduction molecules, which control diverse pathways throughout the acclimation of the plant under stress conditions [5,12]. Several studies showed that ROS are essential for the success of numerous fundamental natural processes, including cellular proliferation and differentiation [11]. In addition, H2O2 is a critical component of stress response regulation in crop plants such as rice [13], wheat [14], maize [15], mung bean [16], soybean [17], cucumber [18], sour orange [19], strawberry [20], basil [21], and rapeseed [22]. Moreover, it is established that in addition to ROS, reactive nitrogen species (RNS), reactive sulfur species (RSS), and reactive carbonyl species (RCS) also play a key signaling role and are all involved in a cross-talk in plant abiotic stress tolerance [23]. Therefore, ROS play a crucial, dual role in plant biology, representing a fascinating area of research for plant biologists.
In this review, we summarize the recent progress of harmful effects of ROS, antioxidant defense system involved in ROS detoxification under different abiotic stresses, and also the cross-talk of RNS, RSS, and RCS with ROS. We also focus on progress in molecular approaches of ROS-mediated improvement in plant antioxidant defense during the acclimation process against abiotic stress.
2. Chemistry of Reactive Oxygen Species
Atmospheric O2 is a free molecule that exists in the ground state (triplet oxygen, 3O2) having two unpaired parallel spin electrons with the same spin numbers, which drop off its reactivity. However, additional energy from some biochemical reactions, electron transport chains (ETC), ultraviolet-B, and ionizing irradiations assist 3O2 to get rid of the spin restriction and thus becoming ROS (Figure 1) [24].
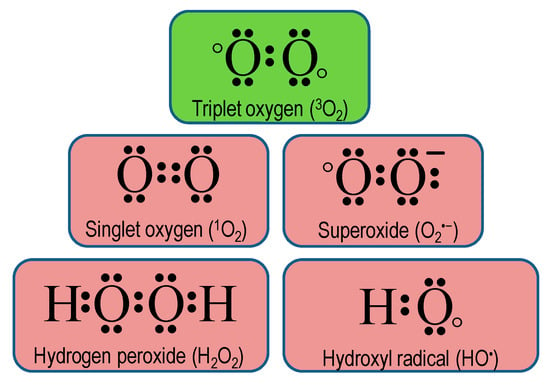
Figure 1.
Lewis dot structure of triplet oxygen and reactive oxygen species.
In plant cells, ROS can be formed in many compartments including chloroplasts, mitochondria, peroxisomes, and plasma membrane [25]. In the chloroplast, the chlorophyll (chl) pigments absorb light quanta and become excited to their triplet state. If this triplet chl is not quenched efficiently, a charge recombination occurs leading 3O2 to excited 1O2 [25]. Although its lifetime is very short (3.1–3.9 μs) and diffusion distance is low (190 nm), 1O2 diffuses outside the chloroplast to reach the cell wall, targets plasma membrane, tonoplast, or even cytosolic signaling cascades [26]. Furthermore, 3O2 can receive electrons from ETC or nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity producing O2•−, which has a half-life of 1–1000 μs [4]. In addition, O2•− reacts with H+ producing HO2•−, which is far more reactive, stable, and permeable through biological membranes. Similarly, H2O2 can be produced through the dismutation of O2•−/HO2•− by SOD isoforms, NADPH oxidases, and heme-containing class III peroxidases (POX) activity [27,28]. Chemically, H2O2 acts as a weak acid that is highly diffusible and stable, having a lifetime of <1 s, and could cross the plasma membrane via aquaporins [29]. Another important ROS—•OH, can be produced by the Fenton reaction, hydroperoxides activity during sunlight, and inner-sphere electron transfer. Moreover, specific proteins, such as heme oxygenases, cytochrome P450s, superoxide reductases, and some photosystem II (PSII) proteins, also generate •OH [30]. The computed half-life of •HO is approximately 1 ns and has a short diffusibility (<1 nm).
Some reactions of production and conversions of ROS in the biological system:
O2 + e− → O2•−
O2•− + H+ ⇄ HO2•−
H2O2 + HO• ⇄ HO2•− + H2O
•OH + •OH ⇄ O• + H2O
O2•− + 2H+ + e− → H2O2
O2•− + HO2•− + H2O → H2O2 + O2 + •OH
HOOH → HO• + •OH
ROOH → RH• + •OH
Fe3+/Cu2+/Mn3+ + H2O2 → Fe2+/Cu+/Mn2+ OH + •OH
Fe2+/Cu+/Mn2+ + H2O2 → Fe3+/Cu2+/Mn3+ + HO2•− + H+
Cellular ROS are comprised of both free radical and nonradicals (Figure 2). Among the free radicals, O2•−, •OH, RO•, and peroxyl radical (ROO•) and nonradicals, H2O2, 1O2, and ozone (O3) are common [31]. However, some other nonradical ROS are also found in plants such as hypochlorous acid (HOCl), hydroperoxides (ROOH), and excited carbonyls (RO*) [32]. Moreover, reactive oxygen intermediates (ROI) are also classified as reactive oxygen molecules formed by incomplete reduction of O2; therefore, ROS include all types of ROI as well as O3 and 1O2. In addition, some acids like hypobromous acid (HOBr), hypoiodous acid (HOI), and HOCl and radicals like carbonate radical (CO3•−) and semiquinone (SQ•−) are also incorporated into ROS [33,34,35].
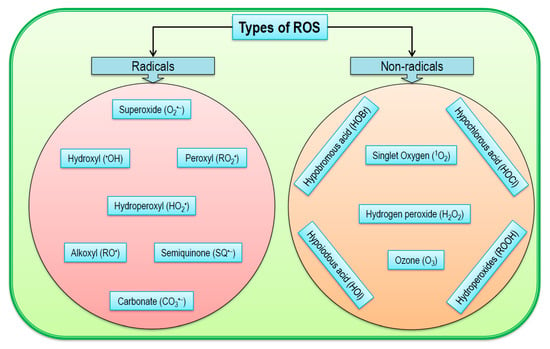
Figure 2.
Types of reactive oxygen species in plants.
Among ROS, O2•− predominantly acts as a reducing agent forming strong oxidants. Moreover, O2•− reacts with nitric oxide (NO) producing RNSs, RSSs, and RCSs. These compounds also promote oxidative stress, and are involved in “shaping” the intra- and extracellular redox signal [36,37].
3. Localization and Processes of the Generation of ROS in Plant Cells
In plant cells, chloroplasts, mitochondria, peroxisomes, plasma membrane, and cell wall are the major locations of ROS generation [38,39]. Therefore, compartmental ROS generation amounts to its overall production in plants (Figure 3) [6,40].
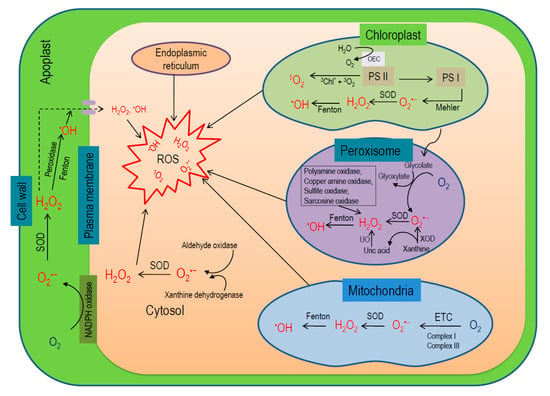
Figure 3.
Localization and processes for the generation of ROS in plant cells (ROS, reactive oxygen species; H2O2, hydrogen peroxide; O2•−, superoxide anion; 1O2, singlet oxygen; •OH, hydroxyl radical; SOD, superoxide dismutase; UO, urate oxidase; XOD, xanthine oxidase; ETC, electron transport chain; PS I, photosystem I; PS II, photosystem II; NADPH, nicotinamide adenine dinucleotide phosphate).
Chloroplasts are the prime sites for ROS production (30–100 times higher than mitochondria), depending on the interaction of chl and light, where triplet chl and ETC of PS I and II are the main sources of ROS production [6,40,41]. In PS II under illumination, chl in light harvesting complex (PSII-LHC) becomes excited to high-energy singlet state (1Chl*; short-lived, ~10−8 s). A portion of this energy is transferred to P680 by photochemical quenching (pQ) for driving the photosynthetic ETC. However, if the absorbed energy exceeds the pQ capacity, the excess energy is dissipated as heat or fluorescence or via intersystem crossing forming 3Chl* (lower energy; longer half-life, ~10−3 s) [42]. The carotenoids present in the LHC (lutein and zeaxanthin) quench 3Chl* preventing transfer of energy to other molecules. If this 3Chl* is not efficiently quenched, it reacts with 3O2 released from splitting of H2O in oxygen-evolving complex (OEC) leading to the formation of 1O2 [43]. Moreover, in the PSII reaction center (RC), P680 absorbs light energy and becomes excited to singlet state (1P680*) pairing with pheophytin (Pheo), 1(P680 + Pheo−) and later, transfers an electron to the quinone (QA), forming P680+ QA−. In an adverse situation, if QA is previously reduced thus being unable to accept any further electrons, 3(P680 + Pheo−) recombines with P680 forming an excited state 3P680* [44]. In the PSII RC, two molecules of β-carotene are present, which are capable of quenching this high-energy 3P680*, but the distance between them is too large (more than the Van der Waal’s distance of 3.6 Å) and quenching remains unsuccessful leading to generation of 1O2 [45]. In addition, some abiotic stresses causing stomatal closure drop chloroplastic carbon dioxide (CO2) levels leading to overreduction of the ETC and enhance the probability of charge recombination between 1P680* and QA− in PS II, increasing 1O2 production [46]. On the other hand, 1O2 is not produced at PS I [47], instead, O2•− can be produced by Mehler reaction and later converted into H2O2 by SOD [48]. Later on, metal ions such as Fe2+ converts both O2•− and H2O2 to highly stable •OH [6]. In the nongreen plant parts, especially in roots, mitochondria are the main source of ROS production, where electron leakage from both complex I and III of ETC produces O2•−, which later catalyzed into H2O2 by Mn-SOD and CuZn-SOD [6,48]. In peroxisomes, glycolate oxidase (GOX) is the main source of ROS production [49]. Moreover, xanthine oxidase (XOD) activity can produce O2•− and uric acid in peroxisomal matrix, which further dismutates to H2O2 by SOD and urate oxidase (UO), respectively [50,51,52]. Besides β-oxidation of fatty acids, O2•− disproportionation and flavin oxidase activity could also produce H2O2 in peroxisomes [49,53]. In addition, polyamine oxidase, copper amine oxidase, sulfite oxidase, and sarcosine oxidase enzyme activity also can generate H2O2 in peroxisome [54]. However, the enzyme MDHAR has been demonstrated in peroxisomes, which helps to scavenge H2O2 by AsA-GSH cycle and regenerate AsA [55,56]. In the apoplast, NADPH oxidase, class III POX, amine and germin-like oxalate oxidases, quinine reductase, and lipoxygenases (LOX) contribute in ROS generation [11,57]. In the cell wall, POX, amine oxidases, and LOX activity are the potential source for ROS in the cell wall [7,58]. Furthermore, NADPH oxidase and quinone reductase guided the ROS production in the plasma membrane [58,59]. Apart from these prime sites, cytochrome (Cyt) P450 produces O2•− in the endoplasmic reticulum. In this process, a radical intermediate (Cyt P450R−) is formed first by the reaction between Cyt P450 and organic substrate (RH) reduced by a flavoprotein. The resulted intermediate further reacts with 3O2 forming a radical oxygenated complex Cyt P450-ROO−, which is finally reduced by Cyt b or spontaneously decomposed to release O2•− [58]. Fatty acid oxidation, as well as GOX and UO activities, produce O2•− and H2O2 in glyoxysomes [60]. Moreover, both XOD and aldehyde oxidase (AO) are potentially involved in cytosolic ROS production [61].
4. Oxidative Stress in Plants and Downstream Implications
Redox reactions (transfer of electrons between a donor and an acceptor) are very common in living organisms, which is responsible for the production of ROS [62]. In plant cells, redox homeostasis is developed in consequence of the equilibrium between the generation of ROS and the functioning of the antioxidant enzymes where efficient defense system in plants keeps the proper balance between ROS generation and elimination [63]. A basal level of ROS, which is maintained above cytostatic or below cytotoxic concentration is, therefore, indispensable for proper ROS or redox signaling in cells, and this level is maintained by the balance between ROS production and ROS scavenging [4,11]. Therefore, scientists used the term “redox biology” to refer to ROS as signaling molecules to control and uphold the usual physiological activities of plants [11,64,65]. Redox signaling has been discerned as the equilibrium between low levels of ROS functioning as signals to activate signaling cascades that adjust usual plant functions and high levels of ROS causing oxidative cellular damage [62]. Therefore, a steady balance between ROS generation and ROS scavenging systems is strongly synchronized over time and space, working together with the cellular redox-sensitive components to shape and finely adapt downstream signaling procedures in a cell-specific and context-specific approach [66,67]. However, any disturbance in the equilibrium of ROS generation and ROS scavenging by antioxidants leads to ROS overaccumulation resulting in oxidative stress under various abiotic stress conditions [9]. Oxidative stress causes lipid peroxidation, damages nucleic acids and proteins, and alters carbohydrate metabolism, resulting in cell dysfunction and death (Figure 4) [4,68].
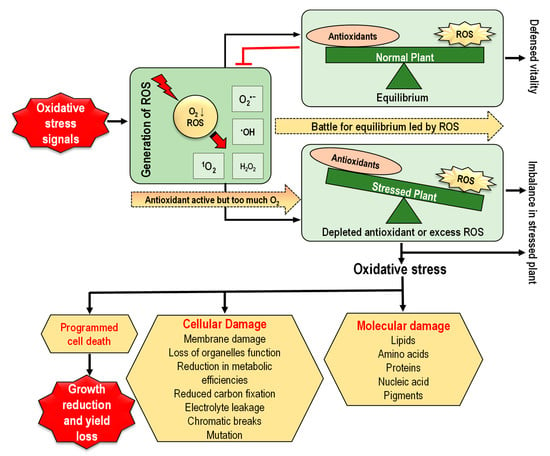
Figure 4.
Oxidative stress in plants and its consequences (ROS, reactive oxygen species; 1O2, singlet oxygen; O2•−, superoxide anion; H2O2, hydrogen peroxide; •OH, hydroxyl radical).
5. Oxidative Stress under Abiotic Stress
Plants are sessile organisms that normally grow under field conditions. Therefore, in most regions of the world, they face excess light (sunny hours) during the hot season. Besides, different environmental/abiotic stresses generated due to anthropogenic activities and harsh climate changes are contributing in inducing oxidative stress through overgeneration of ROS. It is well established that chloroplasts, mitochondria, peroxisomes, apoplast, and plasma membranes are the primary sites of cellular ROS generation but chloroplasts are the leading sites for ROS production [6,40]. Most of the abiotic stresses reduce the availability of CO2 and hinder carbon fixation and contribute to successive reduction of molecular oxygen, which yields excess ROS and impairs the performance of chloroplasts, thus disturbing photosynthetic processes [8]. However, ROS generation greatly varies with plant species, genotypes, stress tolerance level, and duration of stress exposure (Table 1).

Table 1.
Oxidative stress in plants under different abiotic stress factors.
5.1. Oxidative Stress under Salinity
Salinity affects plants by imposing various complications such as ion toxicity, osmotic stress, nutritional deficiency, and genotoxicity, resulting in ROS overproduction and oxidative stress (Table 1) [69]. For instance, Rehman et al. [70] found a 2.5- and a 3-fold, increase in the production of H2O2 together with a 2- and a 3-fold increase in thiobarbituric acid reactive substances (TBARS) content under 100 and 200 mM sodium chloride (NaCl) stress, respectively, compared with control depicting salt-induced oxidative stress condition. It is also reported that the oxidative stress varies among the plant tissues under salt stress. For instance, it was reported that root tissues suffered most from salinity-induced oxidative stress, followed by mature and young leaves. Recently, Cheng et al. [71] reported that the total ROS content, lipid peroxidation, and electrolyte leakage (EL) in rice root tissues were two times higher under salt stress compared with the control. In another study, Ahanger et al. [72] reported an overaccumulation of O2•− and H2O2 (by 157% and 176%, respectively) together with increased malondialdehyde (MDA, by 94%) content and EL (by 158%) confirming salinity (100 mM NaCl)-induced oxidative stress in tomato. Similarly, both MDA and EL were increased by 2-fold due to salt stress (0.4%) in sweet peppers [73], while a 2-fold increase in H2O2, MDA, EL, and O2•− content was found to be caused in mung bean following 100 mM NaCl exposure [74]. Moreover, increased H2O2 (by 50%) and MDA (by 25%) content were noted in maize plants under salt stress (120 mM NaCl) compared with controls [75]. The extent of oxidative stress varied among genotypes within a species. Lalarukh and Shahbaz [76] exposed two sunflower genotypes (FH-572 and FH-621) to salt stress (120 mM NaCl) and observed that H2O2 content increased (by 78%) in FH-572, while decreasing (by 20%) in FH-621, indicating FH-621 as being more salt stress tolerant. In a similar study, Tariq and Shahbaz [77] evaluated two sesame genotypes (TS-5 and TH-6) against salt stress (70 mM NaCl) and concluded that TS-5 showed comparatively better salt tolerance than TS-6. Similarly, Mhadhbi et al. [78] showed a genotype-dependent correlation between salinity tolerance and cellular damage indicators such as MDA and H2O2 content in Medicago truncatula genotypes under salt stress conditions. Interestingly, Ailanthus altissima plants growing under 150 mM NaCl had upregulated antioxidant enzymatic activities and no significant difference in H2O2 content compared with control plants, suggesting a link between the antioxidant defense apparatus and their increased invasiveness in adverse environments [79]. From these examples, it is evident that plants have differential responses towards salt-induced oxidative stress conditions.
5.2. Oxidative Stress under Water Deficit and Simulated Drought
Drought provokes stomatal closure, reduced CO2 entry, and impaired photosynthetic rate, as well as imbalance in the light harvest and utilization and altered photochemistry in chloroplasts, causing ROS overproduction [4,80]. Moreover, protein and membrane denaturation from photorespiration, inactivation of TCA cycle enzymes, and reduced carboxylation efficiency during drought are also linked with ROS overproduction [81]. Additionally, a lower NADP+ regeneration causes greater reduction of ETC, higher EL under drought stress, ultimately resulting in excess ROS metabolism and oxidative stress [82,83]. Many studies have reported drought-induced ROS overproduction and oxidative stress in numerous plant species (Table 1). Abideen et al. [84] grew Phragmites karka under drought conditions by maintaining 40% water holding capacity for 35 d in a plastic tube and found 22% increase of MDA content. Under similar conditions, Campos et al. [85] recorded higher MDA content in Coffea arabica L. after 20 d. Saha et al. [86] created drought conditions for rice plants by withdrawing irrigation for 8 d and found that in contrast to control, drought stress increased O2•−, H2O2, and MDA content by 1.8-, 2.1-, and 1.66-fold, respectively. Severe drought stress (75% water deficit condition) in finger millet plants considerably increased EL and H2O2 content [87]. In another study, Malhotra et al. [88] withheld irrigation in tomato plant for 6 d, which resulted in an increase of MDA content as well as 39% augmentation of EL. Hasanuzzaman et al. [89] and [90] investigated the effect of hyperosmotic stress (10% and 20% polyethylene glycol; PEG) on Brassica napus L. cv. Bina Sharisha-3 and found that both MDA and H2O2 increased under stress conditions. Similarly, hyperosmotic stress (5% PEG, 48 h) induced higher accumulation of H2O2 and O2•− with enhanced membrane peroxidation and LOX activity in Vigna radiata L. cv. BARI Mung-2 [91]. Abbas et al. [92] observed higher TBARS, EL, and H2O2 contents in wheat grown under water deficit condition (70% field capacity; FC). A similar increase in O2•−, H2O2, and MDA content was observed in Oryza sativa L. var. japonica cv. Nipponbare grown under 20% PEG-induced hyperosmotic stress [93]. Rezayian et al. [94] observed significantly increased MDA, H2O2 content, and LOX activity in 15% PEG-stressed Glycine max plants compared with control samples. Rady et al. [95] exposed Solanum lycopersicum L. cv. Login 935 plants to drought stress (60% FC, 20 d) and observed increased O2•−, H2O2, and MDA contents by 75%, 37%, and 83%, respectively. In agreement with this, Filippou et al. [96] recorded significantly increased MDA and H2O2 content in M. truncatula plants under severe drought stress conditions, while parameters were reversed to prestress conditions following rewatering. The extent of drought severity also depends on the genotypic capability to cope with oxidative stress. Kusvuran and Dasgan [97] compared two Phaseolus vulgaris genotypes (Bn-150 (drought-tolerant) and Bn-16 (drought-sensitive)) under drought (50% FC, 14 d), where Bn-16 exhibited 2-fold greater MDA content than Bn-150. Moreover, O2•−, H2O2, and •OH were also found to be higher in Bn-16.
5.3. Oxidative Stress under Metals/Metalloids Toxicity
Metals/metalloids generate ROS in plant cells by disrupting the chloroplastic and mitochondrial electron transfer activities as well as peroxisomal oxidative metabolism. A number of studies demonstrated toxic metals-/metalloids-induced overproduction of ROS and subsequent oxidative damage in different plants (Table 1). A remarkable increase in lipid peroxidation along with the higher accumulation of H2O2 was observed in O. sativa seedlings grown under nickel (Ni; 0.25 and 0.5 mM NiSO4, 72 h) toxicity [98]. El-Amier et al. [99] also reported similar results with lower levels of Ni (100 µM Ni as NiCl2) in Pisum sativum. On the other hand, cadmium (Cd) stress has been shown to increase MDA, H2O2, and O2•– levels in different crops [100,101,102]. For example, Cd stress (100 µM CdCl2) resulted in increased MDA and H2O2 in Arabidopsis thaliana [103] and Cucumis sativus seedlings [104]. In a recent study, Ahanger et al. [105] reported that lipid peroxidation, EL, H2O2, and O2•− contents as well as LOX activity were markedly increased in V. angularis seedlings under Cd stress (100 µM CdCl2, 20 d), while a similar increase in EL, H2O2 and TBARS contents was recorded in Mentha arvensis under Cd stress (50 µM CdCl2, 100 d) [106]. Hasanuzzaman et al. [107] demonstrated higher MDA, H2O2, and O2•− content in wheat plants subjected to lead (Pb) stress (0.5 and 1.0 mM Pb(NO3)2) compared with control. The nonredox metalloid arsenic (As) has also been reported to induce oxidative stress. Cajanus cajan seedlings exposed to As (10 µM As as Na3AsO4, 5 d) had significantly higher H2O2, O2•−, and MDA content, as well as 4-fold increased LOX activity compared with unstressed plants [108]. Such an increase in H2O2 and TBARS contents was also evident in two different cultivars of Chenopodium quinoa under higher levels of As (150 and 300 µM Na3AsO4, 35 d) stress [109]. Interestingly, significant cellular damage in the form of increased MDA and H2O2 content was also observed in basil plants growing under copper (Cu) stress (1000 ppm), although no significant increase in MDA content was observed in plants growing under milder Cu stress conditions (500 ppm) [110].
5.4. Oxidative Stress under High Temperature
High temperature (HT) leads to the overproduction of ROS, resulting in altered cellular metabolism, inactivated oxygen-evolving complex and increased lipid peroxidation, membrane damage, and nicking of DNA, and further, it drives to cell death (Table 1) [111]. High temperature (35 °C/32 °C day/night) blocked PSII reaction center and electron flow, reduced quantum efficiency (Fv/Fm), and downregulated PSII photochemistry in two rice cultivars (IR64 and Huanghuazhan) [112]. Ding et al. [113] reported a 79.9% increase in O2•− content cucumber (C. sativus L.) seedling due to HT stress (35 °C). In tobacco (Nicotiana tabacum cv. Bright-Yellow 2), HT (50 °C) increased 50% of O2•− content over a period of 5 min creating oxidative stress. Likewise, Djanaguiraman et al. [114] observed O2•− content increased by 3.5-fold and 2.3-fold in pollen and pistils, respectively, in field-grown sorghum plants under HT stress, but Liu et al. [93] did not observe any significant change in O2•− and MDA content under HT stress (38 °C, 5 d), in spite of H2O2 increased by 1.27-fold in rice seeds compared with control samples.
5.5. Oxidative Stress under Low Temperature
Low temperature (LT) causes overproduction of ROS in plants through degradation of membrane fluidity, inhibiting photosynthetic apparatus activity, and imbalanced ROS detoxification, which lead to lipid peroxidation and EL (Table 1) [115]. Han et al. [116] showed increased MDA (by 180%) and EL (by 49%) contents in cold-stressed (12 °C, 6 d) 14-d-old rice seedlings. Similarly, Liu et al. [117] treated a cold-sensitive S. lycopersicum (Jinpeng No. 1) genotype with LT stress (15 °C/8 °C day/night; 24 and 48 h), leading to significantly higher MDA and H2O2 content compared with controls. Similarly, Xue et al. [118] evaluated wild-type (WT) and transgenic (G-1 and G-2) Ammopiptanthus mongolicus for LT stress tolerance in a controlled system (4 °C for first 24 h, 0 °C for next 12 h, and −6 °C for last 12 h) and found that WT plants accumulate higher levels of H2O2 compared with transgenic plants (detected through 3,3′-diaminobenzidine staining), indicating oxidative stress under LT stress.
5.6. Oxidative Stress under Flooding
Flooding or waterlogging (WL)-induced hypoxic or anoxic conditions generate toxic compounds that impair plant metabolism resulting in ROS overgeneration and oxidative damages (Table 1) [119]. Zhang et al. [120] also experimented with two Sorghum bicolor genotypes JN01 (WL-tolerant) and JN31 (WL-sensitive) and reported a remarkably higher accumulation of MDA in JN31 compared with JN01, at different duration (6, 9, and 12 d) of WL treatment. Anee et al. [121] studied the WL-sensitive Sesamum indicum L. cv. BARI Til-4 under different durations (2, 4, 6, and 8 d) of WL stress and reported that MDA and H2O2 increased in a duration-dependent manner. Similar enhancement of these oxidative stress markers was also observed in S. lycopersicum [122]. However, an Antarctic plant, named Deschampsia antarctica, also produced higher MDA and H2O2 when exposed to WL condition for 7 d [123].
6. Overview of Plant Antioxidant Defense System
Antioxidants directly or indirectly scavenge ROS and/or control ROS production [138]. The antioxidant defense system consists of low-molecular-weight nonenzymatic antioxidants and some antioxidant enzymes [4]. The nonenzymatic antioxidants such as AsA, GSH, α-tocopherol, phenolic compounds (PhOH), flavonoids, alkaloids, and nonprotein amino acids work in a coordinated fashion with antioxidant enzymes such as SOD, CAT, POX, polyphenol oxidase (PPO), APX, MDHAR, DHAR, GR, GPX, GST, TRX, and PRX in order to inhibit overproduction of ROS (Figure 5) [139,140]. The catalytic reaction of enzymatic and nonenzymatic antioxidants and the reaction sites in cellular organ is represented in Table 2. In plants, the enzyme SOD is directly related to stress, which initiates the first line of defense, converting O2•− into H2O2 (Table 2) [141,142]. This generated H2O2 can be further converted into H2O by the enzymes CAT, APX, GPX, or catalyzed in the AsA-GSH cycle. In plant cell, the AsA-GSH cycle or Asada—Halliwell cycle is the major antioxidant defense pathway to detoxify H2O2, which consist nonenzymatic antioxidants AsA and GSH as well as four important enzymes APX, MDHAR, DHAR, and GR. In the antioxidant defense system, a key role is performed by the AsA-GSH cycle to minimize H2O2 and redox homeostasis [4,143]. In addition, GPX and GST are also vital enzymes for the detoxification of H2O2 and xenobiotics (Figure 5) [144]. Among nonenzymatic antioxidants, AsA and GSH are the most abundant soluble antioxidants in higher plants [145], those play a vital role as electron donors and scavenge ROS directly through AsA-GSH cycle [4]. Moreover, beta-carotene reacts with •OH, O2•−, and ROO• radicals resulting in reduced cellular ROS concentrations [146].
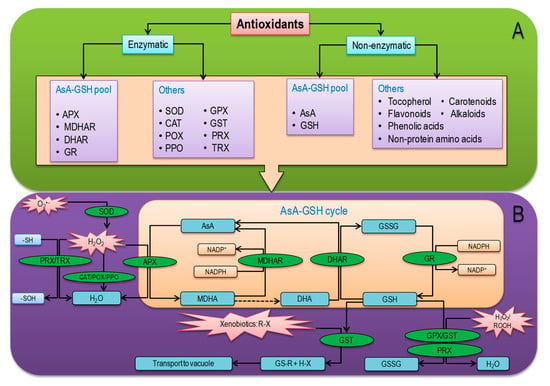
Figure 5.
Overview of plant antioxidant defense system: (A) types of antioxidants and (B) combined mechanisms of enzymatic and nonenzymatic antioxidants. See the text for a more detailed description. APX, ascorbate peroxidase; AsA, ascorbate; CAT, catalase; DHA, dehydroascorbate; DHAR, dehydroascorbate reductase; GPX, glutathione peroxidase; GR, glutathione reductase; GSH, reduced glutathione; GSSG, oxidized glutathione; GST, glutathione S-transferase; H2O2, hydrogen peroxide; MDHA, monodehydroascorbate; MDHAR, monodehydroascorbate reductase; NADPH, nicotinamide adenine dinucleotide phosphate; O2•−, superoxide anion; POX, peroxidases; PRX, peroxiredoxins; R, aliphatic, aromatic, or heterocyclic group; ROOH, hydroperoxides; –SH, thiolate; SOD, superoxide dismutase; –SOH, sulfenic acid; TRX, thioredoxin; X, sulfate, nitrite, or halide group.
Table 2.
Reaction mechanisms of major reactive oxygen species (ROS) scavenging enzymatic antioxidants.
6.1. Nonenzymatic Antioxidants
Ascorbate plays a significant role in AsA-GSH cycle to scavenge ROS through its capacity to donate electrons and remain stable due to electron delocalization that results from the resonance between two forms [4]. Many phytohormone biosynthesis pathways are regulated by AsA. Moreover, AsA regenerates α-tocopherol (vitamin E) from tocopheroxyl radical or by scavenging of •OH and O2•− [147,148]. Contrarily, another vital component of the antioxidant defense system, GSH, plays a significant role in the regulation of AsA-GSH cycle for scavenging cellular ROS and redox homeostasis [4]. Tocopherol protects the chloroplast and maintains photosynthesis by scavenging ROS, mainly 1O2 and •OH [149]. Carotenoids constitute another important class of antioxidant molecules, which are known to scavenge harmful free radicals, as well as to protect light-harvesting complex proteins and thylakoid membrane stability [3,150]. Low-molecular-weight compound flavonoids and, especially, dihydroxy B-ring-substituted flavones and flavonols have great potential to scavenge free radicals and reduce cell damage from lipid peroxidation [151,152,153]. Moreover, abiotic stresses upregulated the expression of genes related to flavonoids biosynthesis and activate antioxidant defense mechanisms [3]. The antioxidant phenolic acids are mainly composed of hydroxybenzoic and hydroxycinnamic acids, those show antioxidant activity as chelators and scavengers of free radicals, especially O2•−, •OH, ROO•, and ONOO– [138]. Alkaloids also have antioxidant ability as free radical scavengers and inhibit H2O2-induced oxidation [154]. Furthermore, nonprotein amino acids (gamma-aminobutyric acid, ornithine, and citrulline) are also considered as effective nonenzymatic antioxidant [155].
6.2. Antioxidant Enzymes
Superoxide dismutase (SOD; EC 1.15.1.1), categorized into three main types—Cu/Zn-SOD, Fe-SOD, and Mn-SOD, leads the frontline defense in the antioxidant defense system by dismutating O2•− into H2O2 and reducing the possibility of •OH formation [156]. In the antioxidant defense system, catalase (CAT; EC 1.11.1.6) is a tetrameric heme-containing enzyme for ROS detoxification, which converts 26 million H2O2 molecules into H2O in 1 minute [3]. Peroxidase (EC. 1.11.1.7) mainly oxidizes PhOH for producing phenoxyl radical (PhO•) more commonly referred to QA, where H2O2 accepts electron and is converted to H2O. In the absence of AsA, PhO• cross-reacts producing suberin, lignin, and quinines, but in the presence of AsA, PhO• reacts with AsA generating monodehydroascorbate (MDHA) and, subsequently, DHA (Figure 5 and Table 2) [157].
Polyphenol oxidase (EC 1.14.18.1) mostly found in thylakoid membrane of chloroplast can influence photosynthesis directly. The enzyme polyphenol oxidase could also interact with peroxidase, or water–water cycle to facilitate ROS scavenging. PPO oxidizes PhOH to QA and H2O by using available O2 [158]. In plant cells, AsA-dependent APX (EC 1.11.1.1) occurs in different isoforms (cytosolic APX (cAPX), mitochondrial APX (mtAPX), chloroplastic APX (chlAPX; APX is the only enzyme capable of scavenging H2O2 in chloroplast since CAT is not present), and peroxisomal/glyoxysomal APX (mAPX; including)) and are H2O2 scavengers, which participates in AsA-GSH cycle producing monodehydroascorbate (MDHA) [159]. The produced MDHA is converted to AsA by a NADPH-dependent flavin adenine dinucleotide enzyme—MDHAR (EC 1.6.5.4)—found as two isoforms in various cellular locations [4]. Monodehydroascorbate reductase containing a thiol group regenerates AsA by phenoxyl radical reduction [160]. Monodehydroascorbate is further reduced into DHA nonenzymatically, which is then recycled to AsA by the activity of GSH-dependent DHAR (EC1.8.5.1) activity [160]. In this reaction, GSH is oxidized to GSSG that is further reduced to GSH by NADPH-dependent GR (EC 1.6.4.2) enzyme, which is also vital for the regulation of redox homeostasis [161]. Beyond, GPX (EC 1.11.1.9) is a member of nonheme containing POX family antioxidant enzyme having a highly reactive thiol group, which utilizes GSH and TRXs to scavenge H2O2, reducing lipids, and organic hydroperoxides [162]. In line, GST (EC 2.5.1.18) conjugates GSH and electrophilic substrates, in its active sites, thus metabolizing xenobiotics (especially, herbicides and pharmaceutically active compounds) and transport them into vacuoles [163,164]. It is also involved in peroxide breakdown, hormone biosynthesis, and stress signaling as well as accelerating GPX activity [165,166]. Moreover, TRX (EC 1.8.1.9), having different isoforms (f, m, h, s, o, x, y, and z), contain an active redox site (WCG/PPC), which reduces disulfide bonds into dithiol by H2O2 and regulate the target proteins faster than GSH or dithiothreitol [167]. Among the isoforms, TRXx and TRXy can regulate redox homeostasis in chloroplast by reducing 2-Cysteine (Cys) PRX, whereas in mitochondria, TRXo1 participates with PRX and sulfiredoxin to activate antioxidant defense [168]. Another antioxidant enzyme, the thiol-based PRX (EC 1.11. 1.15), exhibits POX-like activity neutralizing peroxides (H2O2 and ROOH) in the cytosol, chloroplasts, mitochondria, and nucleus of plant cells [169,170]. The PRXs are thiol-dependent (GSH or any other thiol), playing a vital role in ROS regulation due to the capability of reducing various organic and inorganic peroxides (Figure 5 and Table 2) [171].
7. Antioxidant Defense in Plants under Abiotic Stress: Recent Approaches
Plants activate their antioxidant defense system to mitigate the adverse effects of oxidative stress. However, antioxidant defense capacity varies among plant species and genotypes, as well as stress types and duration (Table 3). Moreover, different approaches to enhance plant antioxidant defense have also been revealed (Table 3).
Table 3.
Antioxidant defense in plants under different abiotic stress factors.
7.1. Antioxidant Defense in Plants under Salinity
Regulation of antioxidant machinery ameliorates the effects of salt stress in plants, as reported in many plant studies (Table 3). Researchers have reported that differential activities of antioxidant enzymes vary according to salinity extent, exposure time, and the developmental stages of plants [172,173]. For instance, Vighi et al. [174] observed differential response in salt-tolerant (BRS Bojuru) rice cultivar compared with salt-sensitive (BRS Pampa) one and concluded that OsAPX3, OsGR2, OsGR3, and OsSOD3-Cu/Zn genes were the basic differential markers between tolerant and sensitive rice genotypes. In another study, Zeeshan et al. [175] compared wheat (salt-tolerant cv. Suntop and -sensitive Sunmate) and barley (salt-tolerant cv. CM72) cultivars and concluded that higher activities of antioxidants (SOD, peroxidase; POD, APX, GR, and CAT) are strongly correlated with the higher salt tolerance depicting a clear role of antioxidant activities in mitigation of salt-induced oxidative stress. Similarly, Alzahrani et al. [124] found increased SOD, CAT, GR, and AsA levels in faba bean genotypes, when H2O2 increased above 90% under salinity stress, confirming the regulation of antioxidant response under salt stress and its mitigation. The regulation of antioxidant activities through the use of either chemical or natural protectants under salt stress has also confirmed the role of plant antioxidant machinery in ameliorating stresses such as salinity [176,177,178]. For instance, Alsahli et al. [179] found that a 2-fold increase in SOD, CAT, and APX activity decreased 3-fold H2O2 in salt-stressed wheat by salicylic acid (SA) application compared with untreated control plants. Similarly, the combined application of jasmonic acid (JA) and humic acid also increased APX activity, resulting in salinity tolerance in sorghum [180], while exogenous application of polyamines regulated sour orange antioxidant responses under salinity stress conditions [181]. Nitrogen supplementation is also reported to increase the antioxidant (SOD, CAT, APX, GR, MDHAR, DHAR activities and the biosynthesis of AsA and GSH) levels with declining 2.5-fold H2O2 and 1.7-fold O2•− generation in wheat under 100 mM NaCl stress [182]. Moreover, silicon (Si) supplementation also increased antioxidant activities and decreased ROS, MDA, and EL levels in mung bean under salinity [74]. Chung et al. [183] reported Si-induced upregulation of antioxidant enzyme genes GmCAT1 (by 3-fold), GmCAT2 (by 4-fold), and GmAPX1 (by 8-fold), leading to salt stress tolerance in soybean after 6 h of stress exposure. Similar transcriptional regulation of antioxidant enzyme transcript levels (cAPX, CAT, GR, and MnSOD) decreased 0.4-fold H2O2 and 3.9-fold NO in hydrogen sulfide (H2S)-primed strawberry plants under NaCl stress in a hydroponic setup [184]. Santander et al. [185] reported that arbuscular mycorrhizae-induced increased SOD, CAT, and APX activities in 40 and 80 mM NaCl-stressed cucumber. Moreover, Moringa oleifera leaf extract (6%) or Glycyrrhiza glabra root extract mitigates salt stress by upregulating antioxidants in wheat and bean (P. vulgaris) [186,187]. Finally, supplementation of fungicidal compound penconazole (15 mg L−1) regulated SOD, CAT, POX, and PPO activity to mitigate the negative impact of salinity in sesame (S. indicum) [188]. From the above-discussed examples, the overall influence of plant antioxidant system in the mitigation of salt stress and associated oxidative stress conditions is clear and an immensely important factor to be aware of.
7.2. Antioxidant Defense in Plants under Water Deficit and Simulated Drought
Activating antioxidant defense as an adaptive mechanism against drought stress was reported in different plants (Table 3) [83,89,90]. Nahar et al. [91] demonstrated decreased AsA/DHA and GSH/GSSG ratio with increased APX, GR, GPX, and GST activities in drought-exposed V. radiate seedlings compared with control, which contributed in drought-induced oxidative damage tolerance. Akram et al. [189] compared the performances of two canola cultivars B. napus (cv. Dunkeld and Cyclone) exposed to water deficit condition (60% FC, 21 d) and found increased total phenolic contents as well as upregulated CAT and POD activities in both cultivars. When studying two Sorghum bicolor L. cultivars (M-81E (tolerant) and Roma (sensitive)), Guo et al. [190] found that drought stress increased 28.9% and 54.9% H2O2 in M-81E and Roma, respectively, relative to control, when SOD activity increased 1.6 and 1.1 times and APX activity increased 1.7 and 0.9 times, respectively, to improve drought tolerance. Yet, inhibited CAT activity but enhanced GPX activity was found under drought stress (irrigation stopped at 10 days after sowing, 11 d) in Triticum aestivum cv. Sakha-94 [191]. Meanwhile, a comparative study with Zea mays cv. Xida 889 and Xida 319 subjected to drought (50% FC, 15 d) reported that GSH content increased by 17% and 28% in Xida 319 and Xida 889, respectively, compared with the well-watered condition [150]. In addition, Rady et al. [95] observed higher H2O2 (26.2%) and O2•− (51%) generation with enhanced SOD, CAT, and APX activities by 110%, 66%, and 77%, respectively, as well as significantly increased AsA, GSH, and α-tocopherol content in S. lycopersicum cv. Login 935 exposed to drought stress (60% FC, 20 d), which indicates increased antioxidant capacity to tolerate drought-induced oxidative stress. Improved tolerance against drought stress through the regulation of the antioxidant apparatus has also been shown in a number of chemical priming approaches, such as that of Antoniou et al. [130] where pretreatment of M. sativa plants with melatonin resulted in increased CAT activity and lowered H2O2 content compared with unprimed, drought-stressed plants. Similarly, the employment of nitric oxide and hydrogen sulfide aspirin (NOSH-aspirin) leads to improved performance in M. sativa plants under severe drought stress through the regulation of CAT and SOD activity, as well as cAPX, Cu/ZnSOD, and FeSOD transcripts [192].
7.3. Antioxidant Defense in Plants under Toxic Metals/Metalloids
Metals/metalloids toxicity tolerance is positively correlated with improved antioxidant activities for ROS detoxification and metal chelation (Table 3) [68,193]. Among major antioxidants, GST assists GSH to reduce metals/metalloids toxicity by conjugating with them [166]. Additionally, GSH works as a cytosolic precursor of phytochelatins (PC), which bind the metals and facilitates the compound transport into cell vacuole by catalyzing the shuttle of metal ions and other xenobiotics [9,194]. Movement of cytosolic metals/metalloids ions into the vacuole in inert form reduces cellular toxicity [68]. Moreover, both GST and GSH contribute in the accumulation of some flavonoids (anthocyanin), which also act as metal binder and may use the same pathway to be accumulated into the vacuole [195,196]. Hasanuzzaman et al. [98] observed an enhancement in both the GSH and GSSG contents in O. sativa seedlings under Ni stress (0.25 and 0.5 mM NiSO4.7H2O), among which GSH was further increased, but GSSG declined by exogenous Si (0.05 mM Na2SiO3) application signifying the role of Si in upregulating GSH. Moreover, Ni stress-induced enhancement of SOD, GPX, APX, MDHAR, DHAR, and GR activities, which was further upregulated by Si supplementation that helped to minimize Ni toxicity. Ahanger et al. [105] reported an increment in GSH and tocopherol content along with SOD, GST, and DHAR activities with higher H2O2 (61%) and O2•− (47%) content in Cd-stressed (100 µM CdCl2, 20 d) V. angularis seedlings, whereas AsA levels and CAT activity declined. Contrarily, SOD, CAT, POX, and GR activities were upregulated with higher content of H2O2 (53.45% and 69.83%, respectively) under Cd stress (50 µM CdCl2, 100 d) in two Mentha arvensis (cv. Kosi and Kusha) genotypes pointing out the activation of an antioxidant defense system for conferring Cd toxicity tolerance [106]. The authors also reported a further upregulated antioxidant defense following application of gibberellic acid, triacontanol, or SA. Mahmud et al. [102] measured the AsA, DHA, GSH, and GSSG contents of B. juncea seedlings grown under Cd toxicity (0.5 and 1.0 mM CdCl2, 3 d) and found that AsA content along with CAT, MDHAR, DHAR, and GR activities declined in a dose-dependent manner, which was reversed by citric acid (CA, 0.5 and 1.0 mM) cotreatment. Moreover, CA cotreatment increased GSH content, SOD, APX, and GPX activities, and further assisted in reducing oxidative stress [102]. Lead (Pb) stress (1.0 mM Pb(NO3)2) resulted in lower AsA content as well as declined CAT, MDHAR, GR, and GPX activities with increasing H2O2 content by 41% and 95% at mild and severe stress, respectively, in wheat seedlings, which was inversely altered by 1.0 mM of GSH supplementation, thus demonstrating the effect of GSH in activating antioxidant defense system [107]. However, exogenous spermidine assisted in the restoration of AsA and GSH contents, as well as AsA/DHA and GSH/GSSG ratio, together with APX, DHAR, GR, and CAT activity, resulting in lower aluminum (Al; AlCl3 0.5 mM, 48 and 72 h)-induced oxidative stress in V. radiate seedling [133].
7.4. Antioxidant Defense in Plants under High Temperature
Like other abiotic stress factors, the antioxidant defense mechanism is activated to cope with high temperature (HT) stress in plants (Table 3) [113,197], but overall antioxidant capacity differs between species as well as tolerant and sensitive genotypes [9]. According to Kumar et al. [149], the activity of APX and GR was significantly suppressed in sensitive chickpea genotypes (ICC14183 and ICC5912) with increasing almost 2-fold H2O2 under HT conditions in comparison with tolerant genotypes (ICCV07110 and ICCV92944). Liu et al. [93] reported decreased SOD and CAT activities with corresponding suppressed OsSOD, OsCAT, and OsAPX2 expression, resulting in higher accumulation of H2O2 (1.27-fold) in germinating rice seeds under HT stress. Sarkar et al. [198] found elevated activity of CAT and POX in wheat genotypes under HT stress (30 °C). In another example, Zandalinas et al. [199] observed increased GSH and AsA content in Carrizo citrange along with enhanced SOD and CAT activity compared with Cleopatra mandarin under HT stress (40 °C). Furthermore, Sarwar et al. [134] pretreated cotton plants with H2O2 under HT stress and found increased SOD and CAT activity in comparison with unprimed, HT-stressed plants. Similar findings were reported by Christou et al. [20] who showed that strawberry plants pretreated with sodium hydrosulfide (NaHS) under HT stress (42 °C, 8 h) became more resilient than unprimed, stressed plants, and this was linked with the enhanced transcription of AsA (GDH) and GSH biosynthetic enzymes (GS, GCS), as well as enzymatic antioxidants (cAPX, CAT, MnSOD, and GR).
7.5. Antioxidant Defense in Plants under Low Temperature
Plants activate the antioxidant defense system to cope with low temperature (LT) stress as well (Table 3). A 3- and 2-fold increased Cu-ZnSOD and Fe-SOD activities, respectively, to a response of higher H2O2 and O2•− production were reported in cucumber (C. sativus cv. Xinyan 4) seedling exposed to LT [15/8 °C day/night, 8 d] stress [200]. Moreover, significantly increased CAT activity was observed in Cynodon dactylon, Capsella bursa pastoris, and Citrus reticulata, during LT stress [201,202,203]. Contrarily, higher APX activity was observed in Jatropha macrocarpa as a response to high H2O2, which improved LT stress tolerance, whereas reduced APX activity (>6-fold) in J. curcas was linked with increased sensitivity under LT conditions [204]. Cheng et al. [205] experimented with Citrullus lanatus under LT stress (10/5 °C, 7 d) and observed the activation of the antioxidant defense system, where GSH/GSSG and AsA/DHA ratios increased significantly only a day after treatment compared with control samples. Similarly, Wang et al. [206] observed increased AsA and GSH levels as a response to higher H2O2 content in transgenic apple seedlings under LT stress (8 °C, 12 h). More recently, Han et al. [116] exposed 14-d-old rice seedlings to LT (12 °C, 6 d) stress and reported higher content of H2O2 and O2•− accumulation increased SOD and CAT activity along with enhanced GSH/GSSG ratio.
7.6. Antioxidant Defense in Plants under Flooding
Several crop species exhibit their ability to survive under the flooded or WL condition for short or even longer durations by activating antioxidant defense systems (Table 3). Li et al. [207] exposed 18 maize genotypes to WL conditions. After 2 d of WL stress, 19–57% higher SOD activity was observed in 12 genotypes, 19.16–106.96% higher POD activity was found in 13 genotypes, while 26–57% higher CAT activity was found in only 9 genotypes. Lower AsA but increased GSH and GSSG content along with higher H2O2 content were observed in sesame seedling under WL stress in a time-dependent manner [121]. However, AsA-GSH cycle enzymes were not regulated in the same manner, showing significantly higher APX and MDHAR activity and lower DHAR and GR activity, during prolonged (8 d) WL stress [121]. Moreover, Park and Lee [123] recorded higher H2O2 (52%) accumulation increased 91% higher CAT activity compared with controls in the Antarctic plant D. antarctica exposed to WL (7 d) conditions.
8. Revisiting ROS Signaling in Plant Defense
Excess ROS are generated under abiotic stress owing to the disturbance of different metabolic functions and physiological disorders [5]. The antioxidant defense pathways such as the AsA-GSH pathway require energy in the form of NADPH, and once this energy is depleted, these pathways would be incapable of avoiding ROS toxicity [5,218]. However, the functions of ROS (especially H2O2) in plant responses to stresses came into the spotlight at the end of the 20th and the beginning of the 21st century. Few groups of scientists recognized H2O2 as a signaling molecule, which leads to acclimation processes and confers tolerance under different biotic and abiotic stresses [219,220]. Reactive oxygen species generated in the chloroplast during stress might divert electrons from the photosynthetic machinery preventing overload of the antenna and subsequent damage. Reactive oxygen species also protect mitochondria in a similar way [5,221]. Cell wall peroxidase might contribute to ROS generation towards signaling where H2O2 utilizes Ca2+ and MAPK pathway as a downstream signaling cascade. Moreover, plant hormones, especially ethylene (ET) and abscisic acid (ABA), are involved with stress responses via cross-talk with ROS and enhance stress tolerance, which confirms the dual role of ROS under stress condition [222]. Besides signal transduction and interaction with hormones, ROS can also regulate metabolic fluxes under abiotic stress conditions, which jointly control plant acclimation processes where redox reactions control transcription and translation of stress acclimation proteins and enzymes, ultimately protecting plant cells from damage [5,11]. Moreover, H2O2 modulates NO and Ca2+ signaling pathways, which control plant growth and development, as well as other cellular and physiological responses under diverse abiotic stresses [223,224]. As endogenous H2O2 is involved in increasing tolerance against abiotic stress, exogenous application of H2O2 is gaining increasing attention and has largely proved its efficacy [13,18,22,225]. In Table 4, we have listed some key reports dealing with the effect of H2O2 treatment under different abiotic stress conditions.
Table 4.
Effect of exogenous H2O2 in plants under different abiotic stress factors.
Moreover, ROS collaborate with RNS, RSS, and RCS under stress conditions and work jointly in signal transduction pathways [23,226]. Cellular antioxidant levels might be influenced to alter ROS generation and contribute to signaling [227]. On the other hand, RSS influence the manufacture, perception, and further signaling of ROS and RNS [226], while RCS act downstream of ROS as signal mediators under a variety of stress situations [228]. Therefore, the interacting role among ROS, RNS, RSS, and RCS is discussed in the following section.
9. Cross-Talk of Reactive Nitrogen, Sulfur, and Carbonyl Species with ROS
Apart from ROS, other reactive species are produced in plant cells during adverse environmental conditions, including RNS, RSS, and RCS (Figure 6) [146,226,232]. All these reactive species are involved in a molecular cross-talk and have a particular role in cellular signaling cascades [23]. Therefore, the following subsections discuss the intimate relationship among ROS, RNS, RSS, and RCS.
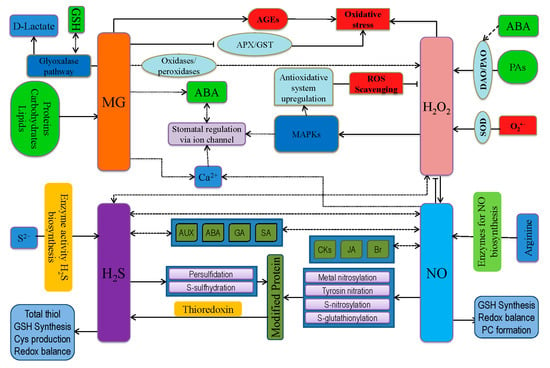
Figure 6.
Cross-talk among vital ROS (H2O2), RNS (NO), RSS (H2S), and RCS (MG) in plant cells for oxidative stress and defense response in plants. APX, ascorbate peroxidase; AUX, auxin; ET, ethylene; ABA, abscisic acid; ROS, reactive oxygen species; GSH, reduced glutathione; JA, jasmonates, MAPKs, mitogen-activated protein kinases; SA, salicylic acid; AEGs, advanced glycation end products; PAs, polyamines; MG, methylglyoxal; NO, nitric oxide; H2S, hydrogen sulfide. Dotted lines represent activation/enhancement.
9.1. Interaction between RNS and ROS
Nitric oxide (NO) is considered to be the most important RNS in plants and is considered as one of three gasotransmitter molecules (Figure 6) [233,234]. During abiotic stress, ROS is overproduced, resulting in enhanced NO generation primarily by nitrate reductase (NR), indicating an interconnection between ROS and RNS [83,235]. As previously reported, NO generation increased by 8-fold in Arabidopsis following exogenous H2O2 application and subsequent NO accumulation activated antioxidant defense system, reduced ROS overgeneration, and reestablished redox balance [83,235]. Contrarily, H2O2 removal by antioxidants or NADPH oxidase inhibitor prevents NO production [236]. Likely, Ca2+-channel inhibitors (ned-19; 3-(2,3-dichlorophenyl)-1,1-dimethylurea and antimycin A) also inhibit H2O2-induced NO production [237]. Moreover, H2O2 is vital for ABA-mediated NO production [238]. Differently, ABA-induced H2O2 production is not NO dependent, which was confirmed by treating with NO donor (sodium nitroprusside, SNP), NO scavenger, and NO synthesis inhibitor. Therefore, H2O2 actively can modulate NO synthesis, via the NR activity [239]. Importantly, both H2O2 and NO play a vital role in signal transduction as well as phytotoxicity [240]. They also cross-react, generating •OH, which is highly reactive.
H2O2 + NO → HNO2 + •OH
Although H2O2 is detoxified by CAT, APX, and GPX, a small amount might escape, which reacts with NO and generates damaging •OH [240,241]. In the absence of metal ions, this reaction is one of the most important mechanisms for generating •OH, providing new insights for ROS-induced tissue-specific oxidative damage as well as signal transduction guided by NO and/or H2O2 [240]. Under abiotic stress conditions, a number of ROS and RNS, e.g., 1O2 and ONOO− might form, leading to oxidative damage [242]. On the other hand, O2•− reacts with NO producing a powerful oxidant, peroxynitrite (ONOO–), which participates in the post-translational modification (PTM) of tyrosine nitration of proteins [243,244]. In peroxisomes, S-nitrosylation inhibits the functioning of glycolate oxidase, CAT, and it can regulate H2O2 levels at the cellular level [245], whereas, the generated ONOO− molecule cause tyrosine nitration and nitrosative alteration in plants. In addition, the proteomic evaluation showed peroxisomal NADH-dependent hydroxypyruvate reductase, which is dysfunctioned by peroxynitrite through nitration [50]. Therefore, it could be concluded that the cross-talk among the ROS and RNS leads to several damaging or signaling episodes where many factors participate for building a complex network of ROS/RNS.
9.2. Interaction between RSS and ROS
Sulfur (S) is the fourth major essential plant nutrient and a structural component for secondary sulfur compounds, such as polysulfides, glucosinolates, PC, thiols, GSH, S-nitrosoglutathione (GSNO), and sulfolipids (Figure 6) [144,234]. Generally, RSS is sometimes called second-generation reactive species as they are formed from the reaction between S and ROS [246]. Interaction of ROS with thiols produces sulfenic acid (R-SOH), further to disulfides, which can donate electrons [246]. Again, R-SOH can undergo PTM (S-glutathionylation and S-cysteinylation), leading to protein stability and proper functioning [247]. Moreover, sulfide-S-oxides can be formed by the decomposition of S-nitrosothiols in the presence of high GSH concentration. This mechanism plays vital role in maintaining the redox balance of thiols as well as modulating S-proteins [248]. Meanwhile, the transformation of R-SOH to sulfinic acid to sulfonic acid is also possible by the oxidation via ROS [249]. H2S actively interacts with ROS to regulate the plasma membrane antiporter (Na+/H+) system [250]. Contrarily, H2S activates enzymatic antioxidants (SOD, CAT, and APX) and enhances GSH content, thus reducing oxidative damages [144,251]. Moreover, H2S is involved in production, perception, and further signal transduction of ROS as well as RNS [234]. As an integral part of the AsA-GSH cycle, redox ratio of GSH:GSSG is important for H2O2 scavenging, which is influenced by H2S [234,251]. Moreover, AsA content is also manipulated by H2S, thus maintaining proper ROS scavenging and acting as a protective molecule at lower cellular concentration [251]. Cysteine plays vital role at the chemical signaling junction of ROS and RSS, where both the molecule regulates signal by Cys-oxidation on proteins. Peroxidation of Cys-S by H2O2 produces the Cys-peroxide, Cys-SeOH [252], whereas persulfidation with H2S2 produces a Cys-persulfide [253]. In addition, Cys-persulfidation prior to protein synthesis has been described [254], but where these Cys-persulfides are targeted yet unidentified as well as the specific regulatory proteins remains to be determined.
9.3. Interaction between RCS and ROS
There is an intimate relationship between RCS and ROS. Reactive carbonyl species include mainly unsaturated aldehydes and ketones produced during lipid peroxidation, which mediate ROS signals. A dozen different RCS are reported to be produced from various membranes [255]. In addition, if RCS is exogenously added to the plants, a similar response as that induced by ROS could be obtained [255]. Considering these criteria, the damaging/signaling role of RCS has been reported to induce root injury, PCD, senescence of leaves and fruits, ABA-mediated stomatal closure, and root response to auxin. Thus, RCS act downstream of ROS as signal transducers during a variety of physiological situations [228]. On the other hand, RCS can modulate antioxidants (CAT, APX, and POD) and thus induce ROS overaccumulation and oxidative stress [256]. It has been demonstrated that RCS-induced ROS generation leads to ABA signaling suggesting ABA-induced ROS (H2O2) production in the guard cell, which increases RCS levels and modulates the signal for stomatal closure [257]. Among RCS, acrolein and 4-Hydroxy-2(E)-nonenal (HNE) formation is stimulated by ROS very early, but other RCS like crotonaldehyde, (E)-2-pentenal, and (E)-2-hexenal are also induced by ROS signals [258]. Furthermore, auxin signaling can induce ROS and RCS formation leading to lateral root initiation [228].
Kaur et al. [259] reported that the generation of methylglyoxal (MG) under stress could overaccumulate ROS directly or induce advanced glycation end products (AGEs) formation. Reports also suggested that increased O2•− production is accelerated by MG [260]. Methylglyoxal also induced ABA or methyl jasmonates or NAD(P)H deficit dependent on stress signaling. Like ROS, RCS might modify Cys residues of proteins in a reversible way to regulate their activity, which would be effective at low levels. However, at higher levels, RCS can have deleterious effects on proteins since histidine and lysine residues can also react with RCS to form stable adducts and might mimic ROS signals potentially associated with regulating activities of proteins such as TRx, which can further regulate activities of other target proteins via redox regulation [256]. Reactive carbonyl species can also interact with zinc ion (Zn2+) and thus release Zn2+ from proteins affecting transcription factors (Zn finger proteins) as well as metabolic and defense enzymes [261]. Therefore, whether RCS could limit the activity of ZnSOD and other antioxidants could be a new area of research. In conclusion, ROS can increase RCS production, with RCS potentially interfering with the antioxidant defense system and exerting oxidative stress. Moreover, RCS can also modulate phytohormone biosynthesis and other signals, thus playing a vital role in constructing a complex ROS/RCS network. Therefore, deeper research should be done in this regard to open a new door of climate-smart crop production.
10. The Transgenic Approach in Enhancing Antioxidant Defense in Plants
In the past few decades, transgenic approaches have been widely used to improve plant health under oxidative stress. Thus, transgenic plants can be engineered to enhance stress tolerance and the activities of antioxidant enzymes. An overview of transgenic plants with enhanced activities of antioxidant defense systems under several stresses is presented in Table 5. Kiranmai et al. [262] isolated a MuWRKY3 gene from drought-adapted horse gram and overexpressed it in groundnut. Transgenic plants showed lower MDA, H2O2, and O2•− contents and enhanced the activities of SOD by 3 to 5-folds and APX by 3 to 7-folds, resulting in increased drought tolerance. Overexpression of MdATG18a in apple enhanced tolerance to drought stress and increased the activities of CAT and POD by 2-fold in transgenic lines. Results also indicated that stress tolerance was improved due to a high frequency of autophagy and restriction of oxidative damage [263]. Overexpression of Chrysanthemum TF gene, DgNAC1 increased salt tolerance in transgenic plants showing lower accumulation of MDA, H2O2, and O2•−, and significantly enhanced the activities of SOD, CAT, and POD [264]. Likewise, PaSOD from Potentilla atrosanguinea and RaAPX from Rheum australe were overexpressed in potato. Transgenic plants demonstrated enhanced activities of SOD and APX in dual transgenic plants (DTP). Superoxide dismutase and APX genes may serve as a positive regulator to increased salt tolerance by the regulation of ROS and lignin biosynthesis signaling pathways in transgenic plants [265]. Transgenic tobacco exhibited tolerance to heavy metals and overexpression of SbMYB15 enhanced the activities of CAT and SOD, also increasing the expression of MnSOD at 100 μM (2-fold) and 300 μM (3-fold) of CdCl2, as well as CAT1 by 62- and 9-fold at 100 and 300 μM of CdCl2 [266]. In regard to heavy metal tolerance, CaGrx from chickpea was overexpressed in A. thaliana. Transgenic plants showed maximal activities of GRX, GR, GPX, GST, and APX under AsIII and Cr stress compared with controls, whereas CAT, SOD, and MDHAR activities were also significantly increased. Authors suggested that CaGrx can be a suitable candidate gene to overcome metal stresses in other crops [267]. Overexpression of the A. thaliana AtDREB1A gene in tomato increased chilling tolerance. Transgenic plants enhanced the activities of SOD by 29% and CAT by 21%, indicating superior chilling stress tolerance [268]. As another example, overexpression of the potato StSOD1 gene enhanced the activities of SOD, POD, and CAT under cold stress and improved cold tolerance in transgenic plants [269]. Similarly, the overexpression of Chrysanthemum CmSOS1 gene enhances SOD and CAT by 171% in transgenic plants under WL conditions [270].
Table 5.
Recent advancements in transgenic approaches to enhance the activities of antioxidant defense systems under abiotic stress conditions. Described studies increased stress tolerance by reducing the damage of oxidative stress and by increasing scavenging of ROS under stressful conditions.
Interestingly, overexpression of DaAPX combined with supplementation of transgenic plants with H2O2 significantly enhanced the activity of APX, improving flooding and cold tolerance [217]. Therefore, a number of genes have been documented to provide significant protection against abiotic stress through genetic modification in several plant species; however, many of the genes are yet to be reported in major crops.
11. Conclusions and Perspectives
It is understood that abiotic stresses are major limiting factors affecting plant growth and development, globally. Thus, there is a growing interest in deciphering the physiological, biochemical, molecular, and cellular mechanisms of abiotic stress responses and tolerance and to introduce potential mitigation techniques that would enhance sustainable agricultural production. Abiotic stresses lead to the accumulation of ROS, which can be a source of oxidative injury in plants. Initially, ROS were considered as toxic molecules and products of aerobic metabolism, found in several subcellular compartments. The metabolism of ROS is crucial in crop growth, development, adaptation, and existence under stressful environments. The production and scavenging of ROS are essential factors of plant defense processes, and modulation and overexpression of candidate genes encoding ROS detoxifying enzymes are widely used to enhance tolerance against several abiotic stresses. However, the balance among the detoxification and generation of ROS is maintained by both enzymatic and nonenzymatic antioxidant systems under stressful conditions. Notably, ROS are known to perform a dual role in plant biology due to molecular cross-talk with other signaling molecules such as RNS, RSS, and RCS. Based on the literature, ROS is very important for various biological mechanisms, such as cellular proliferation and differentiation, and are known to exert a signaling role at low concentrations. However, ROS toxicity openly kills cells through oxidative stress, which is the outcome of ROS-activated pathways responsible for cell death.
In addition, interrelationship exists between ROS, RNS, RSS, and RCS metabolisms under normal and stressed situations; however, a few studies have been carried out to address these interactions. Both ROS and RNS can create oxidative and nitrosative stress solely or together nitro-oxidative stress; however, they are also involved in signaling process of higher plants especially under adverse environmental situations. On the other hand, both ROS and RSS signals are identical and signal through their reaction with Cys, however, RSS signaling appears to be more extensive than ROS signaling. Contrary, RCS can regulate ROS metabolism since these molecules are direct products of oxidative stress and have the potential to act as its sensors. Therefore, these four reactive molecules could be the new gateway of interests for the plant biologists. Although accumulation of knowledge related to signaling roles of these reactive molecules have been accelerated over the last decade, more detailed work is needed to elucidate their roles in plant stress responses.
With the recent progress in molecular and genetic tools, significant progress has been made in enhancing stress tolerance in plants through the development of transgenic plants with increased activities of antioxidant enzymes. Nevertheless, overexpression of genes encoding antioxidant enzymes in transgenic plants has a positive effect on abiotic stress tolerance and the increasing potential of antioxidant enzymes. Based on the available literature, there is a need to identify and report candidate genes that can considerably enhance the tolerance and yield of transgenic plants under stressful environments. In addition, chemical priming offers an attractive alternative to genetic engineering in order to achieve similar goals, often through the regulation of the antioxidant defense apparatus. In the future, systems biology approaches such as genomics, transcriptomics, proteomics, and metabolomics could aid us in introducing new ways for the development of stress tolerance. The integration of these approaches should be implemented to identify key and stress-related regulators, genes, proteins, and metabolites. Furthermore, identification and manipulation of pathways associated with ROS-detoxifying regulators can be improved to generate stress tolerance genotypes.
In the field environment, the plant has to face a variety of stresses at once; thus, identification of core genes, which can confer multiple abiotic stress tolerance, is of paramount importance. In addition, state-of-the-art genome-editing tools like CRISPR/Cas could help to modify the genome through the development of mutant plants with single or multiple genes (ROS-detoxifying regulators) for proper plant growth and development and to enhance the activities of antioxidant defense systems. Recently, speed breeding has also emerged as a powerful tool to boost the growth and development of plants under desired conditions. Therefore, to save time, genome editing could be coupled with speed breeding to develop transgenic plants with induced antioxidant apparatus that are stress tolerant and will thus contribute to feed millions and to ensure world food security.
Author Contributions
Conceptualization, M.H., M.F., and V.F.; writing—original draft preparation, M.H., M.B.B., F.Z., A.R., S.M.M., and J.A.M.; and writing—review and editing, M.H., M.B.B., M.F., and V.F.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We acknowledge Khursheda Parvin, Tasnim Farha Bhuiyan, Taufika Islam Anee, Shahadat Hossen, and Kamrun Nahar for their help in collecting literature and some artworks. V.F. would like to acknowledge support by the Cyprus University of Technology Open Access Author Fund. M.H. acknowledges Sher-e-Bangla Agricultural University Research System (SAURES) for its funding in conducting recent research on plant stress physiology.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
ABA—Abscisic acid; AEGs—Advanced glycation end products; AO—Aldehyde oxidase; RO•—Alkoxy radical; Al—Aluminum; As—Arsenic; APX—Ascorbate peroxidase; AsA—Ascorbic acid; AUX—Auxin; Cd—Cadmium; CAT—Catalase; Chl—Chlorophyll; Cr—Chromium; CA—Citric acid; Cu—Copper; Cys—Cysteine; DHA—Dehydroascorbate; DHAR—Dehydroascorbate reductase; DTP—Dual transgenic plants; EL—Electrolyte leakage; ETC—Electron transport chains; ET—Ethylene; RO*—Excited carbonyls; GRX—Glutaredoxin; GPX—Glutathione peroxidase; GR—Glutathione reductase; GST—Glutathione S-transferase; GOX—Glycolate oxidase; GOPX—Guaiacol peroxidase; HNE—4-Hydroxy-2(E)-nonenal; HT—High temperature; RH—Hydrocarbon; H2O2—Hydrogen peroxide; H2S—Hydrogen sulfide; ROOH—Hydroperoxides; HO2•—Hydroperoxyl radical; •OH—Hydroxyl radical; HOBr—Hypobromous acid; HOCl—Hypochlorous acid; HOI—Hypoiodous acid; JA—Jasmonic acid; Pb—Lead; LOX—Lipoxygenases; LT—Low temperature; MDA—Malondialdehyde; MG—Methylglyoxal; MAPK—Mitogen-activated protein kinase; MDHA—Monodehydroascorbate; MDHAR—Monodehydroascorbate reductase; AOX—NADPH oxidase-like alternative oxidase; Ni—Nickel; NADPH—Nicotinamide adenine dinucleotide phosphate; NR—Nitrate reductase; NO—Nitric oxide; GSSG—Oxidized glutathione; OEC—Oxygen-evolving complex; O3—Ozone; POD/POX—Peroxidase; PRXs—Peroxiredoxins; ROO•—Peroxyl radical; ONOO–—Peroxynitrite; PhOH—Phenolic compounds; PhO•—Phenoxyl radical; pQ—Photochemical quenching; PS I—Photosystem I; PS II—Photosystem II; PSII-LHC—Photosystem II—light harvesting complex; PSII RC—Photosystem II—reaction center; PCs—Phytochelatins; PA—Polyamines; PEG—Polyethylene glycol; PPO—Polyphenol oxidase; PTM—Post-translational modification; PCD—Programmed cell death; QA—Quinone; RCS—Reactive carbonyl species; RNS—Reactive nitrogen species; ROI—Reactive oxygen intermediates; ROS—Reactive oxygen species; RSS—Reactive sulfur species; GSH—Reduced glutathione; RWC—Relative water content; SA—Salicylic acid; SQ•−—Semiquinone; Si—Silicon; 1O2—Singlet oxygen; GSNO—S-nitrosoglutathione; NaCl—Sodium chloride; NaHS—Sodium hydrosulfide; –SOH—Sulfenic acid; O2•−—Superoxide; SOD—Superoxide dismutase; TBARS—Thiobarbituric acid reactive substances; –SH—Thiolate; TRXs—Thioredoxins; 3O2—Triplet oxygen; UO—Urate oxidase; WL—Waterlogging; WT—Wild-type; XOD—Xanthine oxidase.
References
- Pereira, A. Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Mehla, N.; Sindhi, V.; Josula, D.; Bisht, P.; Wani, S.H. An introduction to antioxidants and their roles in plant stress tolerance. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 1–23. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kumar, A.; Yadav, S.; Singh, I.K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene 2019, 18, 100173. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Teixeira da Silva, J.A.; Fujita, M. Plant responses and tolerance to abiotic oxidative stress: Antioxidant defense is a key factor. In Crop Stress and its Management: Perspectives and Strategies; Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, M., Eds.; Springer: Berlin, Germany, 2012; pp. 261–316. [Google Scholar]
- Kaur, N.; Kaur, J.; Grewal, S.K.; Singh, I. Effect of Heat Stress on Antioxidative defense system and its amelioration by heat acclimation and salicylic acid pre-treatments in three pigeonpea genotypes. Indian J. Agric. Biochem. 2019, 32, 106–110. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Antoniou, C.; Savvides, A.; Christou, A.; Fotopoulos, V. Unravelling chemical priming machinery in plants: The role of reactive oxygen–nitrogen–sulfur species in abiotic stress tolerance enhancement. Curr. Opin. Plant Biol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Brestic, M.; Afrin, S.; Sakil, M.A.; Hossain, M.T.; Hossain, M.A.; Hossain, M.A. Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A. Use of Nitric oxide and hydrogen peroxide for better yield of wheat (Triticum aestivum L.) under water deficit conditions: Growth, osmoregulation, and antioxidative defense mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Terzi, R.; Kadioglu, A.; Kalaycioglu, E.; Saglam, A. Hydrogen peroxide pretreatment induces osmotic stress tolerance by influencing osmolyte and abscisic acid levels in maize leaves. J. Plant Interact. 2014, 9, 559–565. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Khan, T.A.; Yusuf, M. Hydrogen peroxide mediated tolerance to copper stress in the presence of 28-homobrassinolide in Vigna radiata. Acta Physiol. Plant. 2014, 36, 2767–2778. [Google Scholar] [CrossRef]
- Guler, N.S.; Pehlivan, N. Exogenous low-dose hydrogen peroxide enhances drought tolerance of soybean (Glycine max L.) through inducing antioxidant system. Acta Biol. Hung. 2016, 67, 169–183. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Liu, S.; Peng, X. Exogenous application of hydrogen peroxide alleviates drought stress in cucumber seedlings. S. Afr. J. Bot. 2016, 106, 23–28. [Google Scholar] [CrossRef]
- Tanou, G.; Filippou, P.; Belghazi, M.; Job, D.; Diamantidis, G.; Fotopoulos, V.; Molassiotis, A. Oxidative and nitrosative-based signaling and associated post-translational modifications orchestrate the acclimation of citrus plants to salinity stress. Plant J. 2012, 72, 585–599. [Google Scholar] [CrossRef]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Gohari, G.; Alavi, Z.; Esfandiari, E.; Panahirad, S.; Hajihoseinlou, S.; Fotopoulos, V. Interaction between hydrogen peroxide and sodium nitroprusside following chemical priming of Ocimum basilicum L. against salt stress. Physiol. Plant. 2020, 168, 361–373. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Yamasaki, H.; Ogura, M.P.; Kingjoe, K.A.; Cohen, M.F. D-Cysteine-induced rapid root abscission in the water fern Azolla pinnata: Implications for the linkage between d-amino acid and reactive sulfur species (RSS) in plant environmental responses. Antioxidants 2019, 8, 411. [Google Scholar] [CrossRef] [PubMed]
- Mailloux, J.R. Application of mitochondria-targeted pharmaceuticals for the treatment of heart disease. Curr. Pharm. Des. 2016, 22, 4763–4779. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, V.A.; Tyutereva, E.V.; Voitsekhovskaja, O.V. Singlet oxygen in plants: Generation, detection, and signaling roles. Int. J. Mol. Sci. 2020, 21, 3237. [Google Scholar] [CrossRef] [PubMed]
- Fischer, B.B.; Hideg, E.; Krieger-Liszkay, A. Production, detection, and signaling of singlet oxygen in photosynthetic organisms. Antioxid. Redox Signal. 2013, 18, 2145–2162. [Google Scholar] [CrossRef]
- Rejeb, K.B.; Benzarti, M.; Debez, A.; Bailly, C.; Savouré, A.; Abdelly, C. NADPH oxidase-dependent H2O2 production is required for salt-induced antioxidant defense in Arabidopsis thaliana. J. Plant Physiol. 2015, 174, 5–15. [Google Scholar] [CrossRef]
- Berwal, M.K.; Ram, C. Superoxide dismutase: A stable biochemical marker for abiotic stress tolerance in higher plants. In Abiotic and Biotic Stress in Plants; De Oliveira, A., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant catalases: Peroxisomal redox guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Maurya, A.K. Oxidative stress in crop plants. In Agronomic Crops; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 349–380. [Google Scholar]
- Kapoor, D.; Sharma, R.; Handa, N.; Kaur, H.; Rattan, A.; Yadav, P.; Gautam, V.; Kaur, R.; Bhardwaj, R. Redox homeostasis in plants under abiotic stress: Role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 2015, 3, 13. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. SnapShot: Reactive oxygen intermediates (ROI). Cell 2010, 140, 951. [Google Scholar] [CrossRef]
- Farnese, F.S.; Menezes-Silva, P.E.; Gusman, G.S.; Oliveira, J.A. When bad guys become good ones: The key role of reactive oxygen species and nitric oxide in the plant responses to abiotic stress. Front. Plant Sci. 2016, 7, 471. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Ann. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Molassiotis, A.; Fotopoulos, V. Oxidative and nitrosative signaling in plants: Two branches in the same tree? Plant Signal. Behav. 2011, 6, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd_Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of subcellular ROS and NO metabolism in higher plants: Multifunctional signaling molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef]
- Dietz, K.-J. Thiol-based peroxidases and ascorbate peroxidases: Why plants rely on multiple peroxidase systems in the photosynthesizing chloroplast? Mol. Cells 2016, 39, 20. [Google Scholar]
- Kim, C.; Dogra, V. Singlet oxygen metabolism: From genesis to signaling. Front. Plant Sci. 2019, 10, 1640. [Google Scholar]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Ann. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2005, 56, 337–346. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Fufezan, C.; Trebst, A. Singlet oxygen production in photosystem II and related protection mechanism. Photosyn. Res. 2008, 98, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Flors, C.; Fryer, M.J.; Waring, J.; Reeder, B.; Bechtold, U.; Mullineaux, P.M.; Nonell, S.; Wilson, M.T.; Baker, N.R. Imaging the production of singlet oxygen in vivo using a new fluorescent sensor, Singlet Oxygen Sensor Green. J. Exp. Bot. 2006, 57, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.; Waszczak, C.; Lewandowska, A.; Willems, P.; Shapiguzov, A.; Li, Z.; Alseekh, S.; Mühlenbock, P.; Hoeberichts, F.A.; Huang, J. Lack of glycolate oxidase1, but not glycolate oxidase2, attenuates the photorespiratory phenotype of catalase2-deficient arabidopsis. Plant Physiol. 2016, 171, 1704–1719. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef]
- Reumann, S.; Chowdhary, G.; Lingner, T. Characterization, prediction and evolution of plant peroxisomal targeting signals type 1 (PTS1s). Biochim. Biophys. Acta 2016, 1863, 790–803. [Google Scholar] [CrossRef]
- Corpas, F.J.; Del Río, L.A.; Palma, J.M. Plant peroxisomes at the crossroad of NO and H2O2 metabolism. J. Integr. Plant Biol. 2019, 61, 803–816. [Google Scholar]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signaling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Plant peroxisomes: A factory of reactive species. Front. Plant Sci. 2020, 11, 853. [Google Scholar] [CrossRef]
- Lisenbee, C.S.; Lingard, M.J.; Trelease, R.N. Arabidopsis peroxisomes possess functionally redundant membrane and matrix isoforms of monodehydroascorbate reductase. Plant J. 2005, 43, 900–914. [Google Scholar] [CrossRef] [PubMed]
- Leterrier, M.; Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; Luis, A. Peroxisomal monodehydroascorbate reductase. Genomic clone characterization and functional analysis under environmental stress conditions. Plant Physiol. 2005, 138, 2111–2123. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Kumar, A.; Kaur, N. ROS and oxidative burst: Roots in plant development. Plant Divers. 2020, 42, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012. [Google Scholar] [CrossRef]
- Heyno, E.; Mary, V.; Schopfer, P.; Krieger-Liszkay, A. Oxygen activation at the plasma membrane: Relation between superoxide and hydroxyl radical production by isolated membranes. Planta 2011, 234, 35–45. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef]
- Janků, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef]
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; Gibon, Y.; Pétriacq, P. Get the balance right: ROS homeostasis and redox signalling in fruit. Front. Plant Sci. 2019, 10, 1091. [Google Scholar] [CrossRef]
- Paciolla, C.; Paradiso, A.; de Pinto, M.C. Cellular redox homeostasis as central modulator in plant stress response. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–23. [Google Scholar]
- Reczek, C.R.; Chandel, N.S. ROS-dependent signal transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Santoro, M.M. ROS signaling and redox biology in endothelial cells. Cell. Mol. Life Sci. 2015, 72, 3281–3303. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, J.A.; Bhuyan, M.H.M.B.; Anee, T.I.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Reactive oxygen species metabolism and antioxidant defense in plants under metal/metalloid stress. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 221–257. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Abbas, G.; Shahid, M.; Saqib, M.; Farooq, A.B.U.; Hussain, M.; Murtaza, B.; Amjad, M.; Naeem, M.A.; Farooq, A. Effect of salinity on cadmium tolerance, ionic homeostasis and oxidative stress responses in conocarpus exposed to cadmium stress: Implications for phytoremediation. Ecotoxicol. Environ. Saf. 2019, 171, 146–153. [Google Scholar] [CrossRef]
- Cheng, Y.-W.; Kong, X.-W.; Wang, N.; Wang, T.-T.; Chen, J.; Shi, Z.Q. Thymol confers tolerance to salt stress by activating anti-oxidative defense and modulating Na+ homeostasis in rice root. Ecotoxicol. Environ. Saf. 2020, 188, 109894. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Mir, R.A.; Alyemeni, M.N.; Ahmad, P. Combined effects of brassinosteroid and kinetin mitigates salinity stress in tomato through the modulation of antioxidant and osmolyte metabolism. Plant Physiol. Biochem. 2020, 147, 31–42. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of sweet pepper with stress tolerance-inducing compounds alleviates salinity stress oxidative damage by mediating the physio-biochemical activities and antioxidant systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) supplementation alleviates NaCl toxicity in mung bean [Vigna radiata (L.) Wilczek] through the modifications of physio-biochemical attributes and key antioxidant enzymes. J. Plant Growth Regul. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Arora, M.; Saxena, P.; Abdin, M.; Varma, A. Interaction between Piriformospora indica and Azotobacter chroococcum diminish the effect of salt stress in Artemisia annua L. by enhancing enzymatic and non-enzymatic antioxidants. Symbiosis 2020, 80, 61–73. [Google Scholar] [CrossRef]
- Lalarukh, I.; Shahbaz, M. Response of antioxidants and lipid peroxidation to exogenous application of alpha-tocopherol in sunflower (Helianthus annuus L.) under salt stress. Pak. J. Bot. 2020, 52, 75–83. [Google Scholar] [CrossRef]
- Tariq, A.; Shahbaz, M. Glycinebetaine induced modulation in oxidative defense system and mineral nutrients sesame (Sesamum indicum L.) under saline regimes. Pak. J. Bot. 2020, 52, 775–782. [Google Scholar] [CrossRef]
- Mhadhbi, H.; Fotopoulos, V.; Mylona, P.V.; Jebara, M.; Aouani, M.E.; Polidoros, A.N. Alternative oxidase 1 (AOX1) gene expression in roots of Medicago truncatulais a genotype-specific component of salt stress tolerance. J. Plant Physiol. 2013, 170, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Filippou, P.; Bouchagier, P.; Skotti, E.; Fotopoulos, V. Proline and reactive oxygen/nitrogen species metabolism is involved in the tolerant response of the invasive plant species Ailanthus altissima to drought and salinity. Environ. Exp. Bot. 2014, 97, 1–10. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Khan, M.I.R.; Al Mahmud, J.; Alam, M.M.; Fujita, M. Regulation of reactive oxygen species metabolism and glyoxalase systems by exogenous osmolytes confers thermotolerance in Brassica napus. Gesunde Pflanz. 2020, 72, 3–16. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought stress responses in plants, oxidative stress, and antioxidant defense. In Climate Change and Plant Abiotic Stress Tolerance; Tuteja, N., Gill, S.S., Eds.; Wiley: Weinheim, Germany, 2014; pp. 209–250. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous nitric oxide donor and arginine provide protection against short-term drought stress in wheat seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ansari, R.; Zulfiqar, F.; Gul, B. Ameliorating effects of biochar on photosynthetic efficiency and antioxidant defence of Phragmites karka under drought stress. Plant Biol. 2020, 22, 259–266. [Google Scholar] [CrossRef]
- Campos, C.N.; Ávila, R.G.; de Souza, K.R.D.; Azevedo, L.M.; Alves, J.D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agric. Water Manag. 2019, 211, 37–47. [Google Scholar] [CrossRef]
- Saha, I.; De, A.K.; Sarkar, B.; Ghosh, A.; Dey, N.; Adak, M.K. Cellular response of oxidative stress when sub1A QTL of rice receives water deficit stress. Plant Sci. Today 2018, 5, 84–94. [Google Scholar] [CrossRef]
- Satish, L.; Rency, A.S.; Ramesh, M. Spermidine sprays alleviate the water deficit-induced oxidative stress in finger millet (Eleusine coracana L. Gaertn.) plants. 3 Biotech 2018, 8, 63. [Google Scholar] [CrossRef]
- Malhotra, C.; Kapoor, R.T.; Ganjewala, D.; Singh, N. Sodium silicate mediated response of antioxidative defense system in Lycopersicon esculentum mill. under water stress. Int. J. Phytomed. 2017, 9, 364–378. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Anee, T.I.; Parvin, K.; Fujita, M. Nitric oxide pretreatment enhances antioxidant defense and glyoxalase systems to confer PEG-induced oxidative stress in rapeseed. J. Plant Interact. 2017, 12, 323–331. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.; Khan, M.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Mahmud, J.A.; Suzuki, T.; Fujita, M. Insights into spermine-induced combined high temperature and drought tolerance in mung bean: Osmoregulation and roles of antioxidant and glyoxalase system. Protoplasma 2017, 254, 445–460. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Mahmood, A.; Zia-ur-Rehman, M.; Ibrahim, M.; Arshad, M.; Qayyum, M.F. Biochar application increased the growth and yield and reduced cadmium in drought stressed wheat grown in an aged contaminated soil. Ecotoxicol. Environ. Saf. 2018, 148, 825–833. [Google Scholar] [CrossRef]
- Liu, J.; Hasanuzzaman, M.; Wen, H.; Zhang, J.; Peng, T.; Sun, H.; Zhao, Q. High temperature and drought stress cause abscisic acid and reactive oxygen species accumulation and suppress seed germination growth in rice. Protoplasma 2019, 256, 1217–1227. [Google Scholar] [CrossRef]
- Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Nitric oxide stimulates antioxidant system and osmotic adjustment in soybean under drought stress. J. Soil Sci. Plant Nutr. 2020, 1–11. [Google Scholar] [CrossRef]
- Rady, M.M.; Belal, H.E.; Gadallah, F.M.; Semida, W.M. Selenium application in two methods promotes drought tolerance in Solanum lycopersicum plant by inducing the antioxidant defense system. Sci. Hortic. 2020, 266, 109290. [Google Scholar] [CrossRef]
- Filippou, P.; Antoniou, C.; Fotopoulos, V. Effect of drought and rewatering on the cellular status and antioxidant response of Medicago truncatula plants. Plant Signal. Behav. 2011, 6, 270–277. [Google Scholar] [CrossRef]
- Kusvuran, S.; Dasgan, H.Y. Effects of drought stress on physiological and biochemical changes in Phaseolus vulgaris L. Legume Res. 2017, 40, 55–62. [Google Scholar]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Mohsin, S.M.; Bhuyan, M.B.; Parvin, K.; Hawrylak-Nowak, B.; Fujita, M. Silicon-induced antioxidant defense and methylglyoxal detoxification works coordinately in alleviating nickel toxicity in Oryza sativa L. Ecotoxicology 2019, 28, 261–276. [Google Scholar] [CrossRef]
- El-Amier, Y.; Elhindi, K.; El-Hendawy, S.; Al-Rashed, S.; Abd-ElGawad, A. Antioxidant system and biomolecules alteration in Pisum sativum under heavy metal stress and possible alleviation by 5-aminolevulinic acid. Molecules 2019, 24, 4194. [Google Scholar] [CrossRef]
- Nahar, K.; Rahman, M.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Physiological and biochemical mechanisms of spermine-induced cadmium stress tolerance in mung bean (Vigna radiata L.) seedlings. Environ. Sci. Pollut. Res. 2016, 23, 21206–21218. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Al Mahmud, J.; Hasanuzzaman, M.; Nahar, K.; Bhuyan, M.B.; Fujita, M. Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: Coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol. Environ. Saf. 2018, 147, 990–1001. [Google Scholar] [CrossRef]
- Gupta, D.; Pena, L.B.; Romero-Puertas, M.C.; Hernández, A.; Inouhe, M.; Sandalio, L.M. NADPH oxidases differentially regulate ROS metabolism and nutrient uptake under cadmium toxicity. Plant Cell Environ. 2017, 40, 509–526. [Google Scholar] [CrossRef]
- Kabała, K.; Zboińska, M.; Głowiak, D.; Reda, M.; Jakubowska, D.; Janicka, M. Interaction between the signaling molecules hydrogen sulfide and hydrogen peroxide and their role in vacuolar H+-ATPase regulation in cadmium-stressed cucumber roots. Physiol. Plant. 2019, 166, 688–704. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Aziz, U.; Sahli, A.A.; Alyemeni, M.N.; Ahmad, P. Combined kinetin and spermidine treatments ameliorate growth and photosynthetic inhibition in Vigna angularis by up-regulating antioxidant and nitrogen metabolism under cadmium stress. Biomolecules 2020, 10, 147. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F.; Fariduddin, Q. Plant growth regulators improve growth, photosynthesis, mineral nutrient and antioxidant system under cadmium stress in menthol mint (Mentha arvensis L.). Physiol. Mol. Biol. Plants 2020, 26, 25–39. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Alharby, H.F.; Fujita, M. Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J. Plant Interact. 2018, 13, 203–212. [Google Scholar] [CrossRef]
- Yadu, B.; Chandrakar, V.; Tamboli, R.; Keshavkant, S. Dimethylthiourea antagonizes oxidative responses by up-regulating expressions of pyrroline-5-carboxylate synthetase and antioxidant genes under arsenic stress. Int. J. Environ. Sci. Technol. 2019, 16, 8401–8410. [Google Scholar] [CrossRef]
- Parvez, S.; Abbas, G.; Shahid, M.; Amjad, M.; Hussain, M.; Asad, S.A.; Imran, M.; Naeem, M.A. Effect of salinity on physiological, biochemical and photostabilizing attributes of two genotypes of quinoa (Chenopodium quinoa Willd.) exposed to arsenic stress. Ecotoxicol. Environ. Saf. 2020, 187, 109814. [Google Scholar] [CrossRef] [PubMed]
- Georgiadou, E.C.; Kowalska, E.; Patla, K.; Kulbat, K.; Smolińska, B.; Leszczyńska, J.; Fotopoulos, V. Influence of heavy metals (Ni, Cu, and Zn) on nitro-oxidative stress responses, proteome regulation and allergen production in basil (Ocimum basilicum L.) plants. Front. Plant Sci. 2018, 9, 862. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Khan, F.; Hassan, S.; Nasim, W.; Arif, M.; Wang, F.; Huang, J. Exogenously applied plant growth regulators affect heat-stressed rice pollens. J. Agron. Crop Sci. 2016, 202, 139–150. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, Y.; He, L.; Zhou, Q.; Yu, J.; Hui, D.; Huang, D. Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci. Rep. 2016, 6, 35424. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Perumal, R.; Jagadish, S.; Ciampitti, I.; Welti, R.; Prasad, P. Sensitivity of sorghum pollen and pistil to high-temperature stress. Plant Cell Environ. 2018, 41, 1065–1082. [Google Scholar] [CrossRef]
- Repetto, M.; Semprine, J.; Boveris, A. Lipid peroxidation: Chemical mechanism, biological implications and analytical determination. In Lipid Peroxidation; Catala, A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 3–30. [Google Scholar]
- Han, Q.-H.; Huang, B.; Ding, C.-B.; Zhang, Z.-W.; Chen, Y.-E.; Hu, C.; Zhou, L.-J.; Huang, Y.; Liao, J.-Q.; Yuan, S. Effects of melatonin on anti-oxidative systems and photosystem II in cold-stressed rice seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]
- Liu, T.; Ye, X.; Li, M.; Li, J.; Qi, H.; Hu, X. H2O2 and NO are involved in trehalose-regulated oxidative stress tolerance in cold-stressed tomato plants. Environ. Exp. Bot. 2020, 171, 103961. [Google Scholar] [CrossRef]
- Xue, M.; Guo, T.; Ren, M.; Wang, Z.; Tang, K.; Zhang, W.; Wang, M. Constitutive expression of chloroplast glycerol-3-phosphate acyltransferase from Ammopiptanthus mongolicus enhances unsaturation of chloroplast lipids and tolerance to chilling, freezing and oxidative stress in transgenic Arabidopsis. Plant Physiol. Biochem. 2019, 143, 375–387. [Google Scholar] [CrossRef]
- Loreti, E.; van Veen, H.; Perata, P. Plant responses to flooding stress. Curr. Opin. Plant Biol. 2016, 33, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhou, Y.; Yue, Z.; Chen, X.; Cao, X.; Xu, X.; Xing, Y.; Jiang, B.; Ai, X.; Huang, R. Changes in photosynthesis, chloroplast ultrastructure, and antioxidant metabolism in leaves of sorghum under waterlogging stress. Photosynthetica 2019, 57, 1076–1083. [Google Scholar] [CrossRef]
- Anee, T.I.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Bhuiyan, T.F.; Alam, M.U.; Fujita, M.; Hasanuzzaman, M. Oxidative damage and antioxidant defense in Sesamum indicum after different waterlogging durations. Plants 2019, 8, 196. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotechnol. 2018, 93, 385–391. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, E.J. Waterlogging induced oxidative stress and the mortality of the Antarctic plant, Deschampsia antarctica. J. Ecol. Environ. 2019, 43, 29. [Google Scholar] [CrossRef]
- Alzahrani, S.M.; Alaraidh, I.A.; Migdadi, H.; Alghamdi, S.; Khan, M.A.; Ahmad, P. Physiological, biochemical, and antioxidant properties of two genotypes of Vicia faba grown under salinity stress. Pak. J. Bot. 2019, 51, 786–798. [Google Scholar] [CrossRef]
- Hossain, M.; Alam, M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Al Mahmud, J.; Fujita, M. Use of iso-osmotic solution to understand salt stress responses in lentil (Lens culinaris Medik.). S. Afr. J. Bot. 2017, 113, 346–354. [Google Scholar] [CrossRef]
- Nxele, X.; Klein, A.; Ndimba, B. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 2017, 108, 261–266. [Google Scholar] [CrossRef]
- Derbali, W.; Goussi, R.; Koyro, H.-W.; Abdelly, C.; Manaa, A. Physiological and biochemical markers for screening salt tolerant quinoa genotypes at early seedling stage. J. Plant Interact. 2020, 15, 27–38. [Google Scholar] [CrossRef]
- Han, F.; Sun, M.; He, W.; Cui, X.; Pan, H.; Wang, H.; Song, F.; Lou, Y.; Zhuge, Y. Ameliorating effects of exogenous Ca2+ on foxtail millet seedlings under salt stress. Funct. Plant Biol. 2019, 46, 407–416. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Anwar, Y.; Hasan, M.; Iqbal, A.; Ali, M.; Alharby, H.F.; Hakeem, K.R.; Hasanuzzaman, M. Attenuation of drought stress in Brassica seedlings with exogenous application of Ca2+ and H2O2. Plants 2017, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, T.F.; Ahamed, K.U.; Nahar, K.; Al Mahmud, J.; Bhuyan, M.B.; Anee, T.I.; Fujita, M.; Hasanuzzaman, M. Mitigation of PEG-induced drought stress in rapeseed (Brassica rapa L.) by exogenous application of osmolytes. Biocatal. Agric. Biotechnol. 2019, 20, 101197. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Suzuki, T.; Fujita, M. Polyamines-induced aluminum tolerance in mung bean: A study on antioxidant defense and methylglyoxal detoxification systems. Ecotoxicology 2017, 26, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Saleem, M.F.; Ullah, N.; Rizwan, M.; Ali, S.; Shahid, M.R.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Exogenously applied growth regulators protect the cotton crop from heat-induced injury by modulating plant defense mechanism. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Diao, Q.; Song, Y.; Shi, D.; Qi, H. Interaction of polyamines, abscisic acid, nitric oxide, and hydrogen peroxide under chilling stress in tomato (Lycopersicon esculentum Mill.) seedlings. Front. Plant Sci. 2017, 8, 203. [Google Scholar] [CrossRef]
- Ghanbari, F.; Sayyari, M. Controlled drought stress affects the chilling-hardening capacity of tomato seedlings as indicated by changes in phenol metabolisms, antioxidant enzymes activity, osmolytes concentration and abscisic acid accumulation. Sci. Hortic. 2018, 229, 167–174. [Google Scholar] [CrossRef]
- Luan, H.; Shen, H.; Pan, Y.; Guo, B.; Lv, C.; Xu, R. Elucidating the hypoxic stress response in barley (Hordeum vulgare L.) during waterlogging: A proteomics approach. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Carocho, M.; Ferreira, I.C. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Nath, M.; Bhatt, D.; Bhatt, M.D.; Prasad, R.; Tuteja, N. Microbe-mediated enhancement of nitrogen and phosphorus content for crop improvement. In Crop Improvement through Microbial Biotechnology; Prasad, R., Gill, S.S., Tuteja, N., Eds.; Academic Press: Cambridge, UK, 2018; pp. 291–301. [Google Scholar]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Biczak, R. Quaternary ammonium salts with tetrafluoroborate anion: Phytotoxicity and oxidative stress in terrestrial plants. J. Hazard. Mater. 2016, 304, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A.; Corpas, F.J.; López-Huertas, E.; Palma, J.M. Plant superoxide dismutases: Function under abiotic stress conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar]
- Fotopoulos, V.; Tanou, G.; Ziogas, V.; Molassiotis, A. Involvement of AsA/DHA and GSH/GSSG ratios in gene and protein expression and in the activation of defense mechanisms under abiotic stress conditions. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Anjum, N.A., Chan, M.T., Umar, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 265–302. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Mahmud, J.; Nahar, K.; Mohsin, S.; Parvin, K.; Fujita, M. Interaction of sulfur with phytohormones and signaling molecules in conferring abiotic stress tolerance to plants. Plant Signal. Behav. 2018, 13, e1477905. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Naz, H.; Akram, N.A.; Ashraf, M. Impact of ascorbic acid on growth and some physiological attributes of cucumber (Cucumis sativus) plants under water-deficit conditions. Pak. J. Bot. 2016, 48, 877–883. [Google Scholar]
- Seminario, A.; Song, L.; Zulet, A.; Nguyen, H.T.; González, E.M.; Larrainzar, E. Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front. Plant Sci. 2017, 8, 1042. [Google Scholar] [CrossRef]
- Kumar, S.; Thakur, P.; Kaushal, N.; Malik, J.A.; Gaur, P.; Nayyar, H. Effect of varying high temperatures during reproductive growth on reproductive function, oxidative stress and seed yield in chickpea genotypes differing in heat sensitivity. Arch. Agron. Soil Sci. 2013, 59, 823–843. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Liu, S.; Ju, J.; Xia, G. Identification of the flavonoid 3′-hydroxylase and flavonoid 3′, 5′-hydroxylase genes from Antarctic moss and their regulation during abiotic stress. Gene 2014, 543, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as antioxidants in plants under abiotic stresses. In Abiotic Stress Responses in Plants; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012; pp. 159–179. [Google Scholar]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.-C.; Mustafa, M.R.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef] [PubMed]
- Vranova, V.; Rejsek, K.; Skene, K.R.; Formanek, P. Non-protein amino acids: Plant, soil and ecosystem interactions. Plant Soil 2011, 342, 31–48. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Jovanović, S.V.; Kukavica, B.; Vidović, M.; Morina, F.; Menckhoff, L. Class III peroxidases: Functions, localization and redox regulation of isoenzymes. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 269–300. [Google Scholar]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Hasanuzzaman, M.; Lao, M.T. Oxidative stress and antioxidant defense in plants under salinity. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 291–309. [Google Scholar]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Xu, J.; Xing, X.-J.; Tian, Y.-S.; Peng, R.-H.; Xue, Y.; Zhao, W.; Yao, Q.-H. Transgenic Arabidopsis plants expressing tomato glutathione S-transferase showed enhanced resistance to salt and drought stress. PLoS ONE 2015, 10, e0136960. [Google Scholar] [CrossRef]
- Christou, A.; Antoniou, C.; Christodoulou, C.; Hapeshi, E.; Stavrou, I.; Michael, C.; Fatta-Kassinos, D.; Fotopoulos, V. Stress-related phenomena and detoxification mechanisms induced by common pharmaceuticals in alfalfa (Medicago sativa L.) plants. Sci. Total Environ. 2016, 557, 652–664. [Google Scholar] [CrossRef]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef]
- Calderón, A.; Sevilla, F.; Jiménez, A. Redox protein thioredoxins: Function under salinity, drought and extreme temperature conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 123–162. [Google Scholar]
- Sevilla, F.; Jiménez, A.; Lázaro, J.J. What do the plant mitochondrial antioxidant and redox systems have to say under salinity, drought, and extreme temperature? In Reactive Oxygen Species and Oxidative Damage in Plants under Stress; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer: Cham, Switzerland, 2015; pp. 23–55. [Google Scholar]
- Pedrajas, J.R.; Bárcena, J.A. Peroxiredoxins: Types, characteristics and functions in higher plants. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 95–121. [Google Scholar]
- Liebthal, M.; Maynard, D.; Dietz, K.-J. Peroxiredoxins and redox signaling in plants. Antioxid. Redox Signal. 2018, 28, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.R.; Neto, M.C.L.; Carvalho, F.E.; Martins, M.O.; Jardim-Messeder, D.; Margis-Pinheiro, M.; Silveira, J.A. Salinity and osmotic stress trigger different antioxidant responses related to cytosolic ascorbate peroxidase knockdown in rice roots. Environ. Exp. Bot. 2016, 131, 58–67. [Google Scholar] [CrossRef]
- Li, Y.; Cao, X.L.; Zhu, Y.; Yang, X.M.; Zhang, K.N.; Xiao, Z.Y.; Wang, H.; Zhao, J.H.; Zhang, L.L.; Li, G.B. Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol. 2019, 222, 1507–1522. [Google Scholar] [CrossRef] [PubMed]
- Vighi, I.; Benitez, L.; Amaral, M.; Moraes, G.; Auler, P.; Rodrigues, G.; Deuner, S.; Maia, L.; Braga, E. Functional characterization of the antioxidant enzymes in rice plants exposed to salinity stress. Biol. Plant. 2017, 61, 540–550. [Google Scholar] [CrossRef]
- Zeeshan, M.; Lu, M.; Sehar, S.; Holford, P.; Wu, F. Comparison of biochemical, anatomical, morphological, and physiological responses to salinity stress in wheat and barley genotypes deferring in salinity tolerance. Agronomy 2020, 10, 127. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 3. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Ahmed, A.-H. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An overview of plant-based natural biostimulants for sustainable horticulture with a particular focus on moringa leaf extracts. Plant Sci. 2019, 110194. [Google Scholar] [CrossRef] [PubMed]
- Alsahli, A.; Mohamed, A.-K.; Alaraidh, I.; Al-Ghamdi, A.; Al-Watban, A.; El-Zaidy, M.; Alzahrani, S.M. Salicylic acid alleviates salinity stress through the modulation of biochemical attributes and some key antioxidants in wheat seedlings. Pak. J. Bot. 2019, 51, 1551–1559. [Google Scholar] [CrossRef]
- Ali, A.Y.A.; Ibrahim, M.E.H.; Zhou, G.; Nimir, N.E.A.; Jiao, X.; Zhu, G.; Elsiddig, A.M.I.; Suliman, M.S.E.; Elradi, S.B.M.; Yue, W. Exogenous jasmonic acid and humic acid increased salinity tolerance of sorghum. Agron. J. 2020, 112, 871–884. [Google Scholar] [CrossRef]
- Tanou, G.; Ziogas, V.; Belghazi, M.; Christou, A.; Filippou, P.; Job, D.; Fotopoulos, V.; Molassiotis, A. Polyamines reprogram oxidative and nitrosative status and the proteome of citrus plants exposed to salinity stress. Plant Cell Environ. 2014, 37, 864–885. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Chung, Y.S.; Kim, K.-S.; Hamayun, M.; Kim, Y. Silicon confers soybean resistance to salinity stress through regulation of reactive oxygen and reactive nitrogen species. Front. Plant Sci. 2019, 10, 1725. [Google Scholar] [CrossRef]
- Christou, A.; Manganaris, G.A.; Papadopoulos, I.; Fotopoulos, V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways. J. Exp. Bot. 2013, 64, 1953–1966. [Google Scholar] [CrossRef]
- Santander, C.; Ruiz, A.; García, S.; Aroca, R.; Cumming, J.; Cornejo, P. Efficiency of two arbuscular mycorrhizal fungal inocula to improve saline stress tolerance in lettuce plants by changes of antioxidant defense mechanisms. J. Sci. Food Agric. 2020, 100, 1577–1587. [Google Scholar] [CrossRef]
- Rady, M.M.; Kuşvuran, A.; Alharby, H.F.; Alzahrani, Y.; Kuşvuran, S. Pretreatment with proline or an organic bio-stimulant induces salt tolerance in wheat plants by improving antioxidant redox state and enzymatic activities and reducing the oxidative stress. J. Plant Growth Regul. 2019, 38, 449–462. [Google Scholar] [CrossRef]
- Rady, M.M.; Elrys, A.S.; El-Maati, M.F.A.; Desoky, E.-S.M. Interplaying roles of silicon and proline effectively improve salt and cadmium stress tolerance in Phaseolus vulgaris plant. Plant Physiol. Biochem. 2019, 139, 558–568. [Google Scholar] [CrossRef]
- Heydari, H.; Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Role of Penconazole in salt stress amelioration in Sesamum indicum L. Soil Sci. Plant Nutr. 2019, 65, 243–250. [Google Scholar] [CrossRef]
- Akram, N.A.; Iqbal, M.; Muhammad, A.; Ashraf, M.; Al-Qurainy, F.; Shafiq, S. Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. Protoplasma 2018, 255, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Tian, S.; Liu, S.; Wang, W.; Sui, N. Energy dissipation and antioxidant enzyme system protect photosystem II of sweet sorghum under drought stress. Photosynthetica 2018, 56, 861–872. [Google Scholar] [CrossRef]
- Hassan, N.; Ebeed, H.; Aljaarany, A. Exogenous application of spermine and putrescine mitigate adversities of drought stress in wheat by protecting membranes and chloroplast ultra-structure. Physiol. Mol. Biol. Plants 2020, 26, 233–245. [Google Scholar] [CrossRef]
- Antoniou, C.; Xenofontos, R.; Chatzimichail, G.; Christou, A.; Kashfi, K.; Fotopoulos, V. Exploring the potential of nitric oxide and hydrogen sulfide (NOSH)-releasing synthetic compounds as novel priming agents against drought stress in Medicago sativa plants. Biomolecules 2020, 10, 120. [Google Scholar] [CrossRef]
- Gratão, P.L.; Alves, L.R.; Lima, L.W. Heavy metal toxicity and plant productivity: Role of metal scavengers. In Plant-Metal Interactions; Srivastava, S., Srivastava, A.K., Suprasanna, P., Eds.; Springer: Cham, Switzerland, 2019; pp. 49–60. [Google Scholar]
- Chakravarthi, S.; Jessop, C.E.; Bulleid, N.J. The role of glutathione in disulphide bond formation and endoplasmic-reticulum-generated oxidative stress. EMBO Rep. 2006, 7, 271–275. [Google Scholar] [CrossRef]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant flavonoids—biosynthesis, transport and involvement in stress responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Awasthi, R.; Gaur, P.; Turner, N.C.; Vadez, V.; Siddique, K.H.; Nayyar, H. Effects of individual and combined heat and drought stress during seed filling on the oxidative metabolism and yield of chickpea (Cicer arietinum) genotypes differing in heat and drought tolerance. Crop Past. Sci. 2017, 68, 823–841. [Google Scholar] [CrossRef]
- Sarkar, J.; Chakraborty, B.; Chakraborty, U. Temperature stress induced antioxidative and biochemical changes in wheat (Triticum aestivum L.) cultivars. J. Plant Stress Physiol. 2016, 22–30. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Modulation of antioxidant defense system is associated with combined drought and heat stress tolerance in citrus. Front. Plant Sci. 2017, 8, 953. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ye, L.; Wang, Y.; Zhou, X.; Yang, J.; Wang, J.; Cao, K.; Zou, Z. Melatonin increases the chilling tolerance of chloroplast in cucumber seedlings by regulating photosynthetic electron flux and the ascorbate-glutathione cycle. Front. Plant Sci. 2016, 7, 1814. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ye, T.; Zhong, B.; Liu, X.; Chan, Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (Cynodon dactylon (L.) Pers.) by exogenous calcium. J. Integr. Plant Biol. 2014, 56, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.A.; Jan, N.; Qazi, H.A.; Andrabi, K.I.; John, R. Cold stress induces biochemical changes, fatty acid profile, antioxidant system and gene expression in Capsella bursa pastoris L. Acta Physiol. Plant. 2018, 40, 167. [Google Scholar] [CrossRef]
- Mohammadrezakhani, S.; Hajilou, J.; Rezanejad, F.; Zaare-Nahandi, F. Assessment of exogenous application of proline on antioxidant compounds in three citrus species under low temperature stress. J. Plant Interact. 2019, 14, 347–358. [Google Scholar] [CrossRef]
- Spanò, C.; Bottega, S.; Castiglione, M.R.; Pedranzani, H.E. Antioxidant response to cold stress in two oil plants of the genus Jatropha. Plant Soil Environ. 2017, 63, 271–276. [Google Scholar]
- Cheng, F.; Lu, J.; Gao, M.; Shi, K.; Kong, Q.; Huang, Y.; Bie, Z. Redox signaling and CBF-responsive pathway are involved in salicylic acid-improved photosynthesis and growth under chilling stress in watermelon. Front. Plant Sci. 2016, 7, 1519. [Google Scholar] [CrossRef]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef]
- Li, W.; Mo, W.; Ashraf, U.; Li, G.; Wen, T.; Abrar, M.; Gao, L.; Liu, J.; Hu, J. Evaluation of physiological indices of waterlogging tolerance of different maize varieties in South China. Appl. Ecol. Environ. Res. 2018, 16, 2059–2072. [Google Scholar] [CrossRef]
- Qin, C.; Ahanger, M.A.; Zhou, J.; Ahmed, N.; Wei, C.; Yuan, S.; Ashraf, M.; Zhang, L. Beneficial role of acetylcholine in chlorophyll metabolism and photosynthetic gas exchange in Nicotiana benthamiana seedlings under salinity stress. Plant Biol. 2020, 22, 357–365. [Google Scholar] [CrossRef]
- Parvin, K.; Nahar, K.; Hasanuzzaman, M.; Bhuyan, M.B.; Mohsin, S.M.; Fujita, M. Exogenous vanillic acid enhances salt tolerance of tomato: Insight into plant antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2020, 150, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Cen, H.; Wang, T.; Liu, H.; Tian, D.; Zhang, Y. Melatonin application improves salt tolerance of alfalfa (Medicago sativa L.) by enhancing antioxidant capacity. Plants 2020, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Shi, Z.; Zhang, X.; Zheng, S.; Wang, J.; Mo, J. Alleviating effects of exogenous melatonin on salt stress in cucumber. Sci. Hortic. 2020, 262, 109070. [Google Scholar] [CrossRef]
- Sehar, Z.; Masood, A.; Khan, N.A. Nitric oxide reverses glucose-mediated photosynthetic repression in wheat (Triticum aestivum L.) under salt stress. Environ. Exp. Bot. 2019, 161, 277–289. [Google Scholar] [CrossRef]
- Jiang, J.-L.; Tian, Y.; Li, L.; Yu, M.; Hou, R.-P.; Ren, X.-M. H2S alleviates salinity stress in cucumber by maintaining the Na+/K+ balance and regulating H2S metabolism and oxidative stress response. Front. Plant Sci. 2019, 10, 678. [Google Scholar] [CrossRef] [PubMed]
- Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Khan, N. Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol. Plant. 2020, 168, 490–510. [Google Scholar]
- Hamim, H.; Violita, V.; Triadiati, T.; Miftahudin, M. Oxidative stress and photosynthesis reduction of cultivated (Glycine max L.) and wild soybean (G. tomentella L.) exposed to drought and paraquat. Asian J. Plant Sci. 2017, 16, 65–77. [Google Scholar] [CrossRef][Green Version]
- Jan, N.; Majeed, U.; Andrabi, K.I.; John, R. Cold stress modulates osmolytes and antioxidant system in Calendula officinalis. Acta Physiol. Plant. 2018, 40, 73. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Wang, Y.-T.; Pan, X.-B.; Xi, Z.-M. Amelioration of cold-induced oxidative stress by exogenous 24-epibrassinolide treatment in grapevine seedlings: Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2019, 244, 379–387. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant. Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kar, R.K. Plant responses to water stress: Role of reactive oxygen species. Plant Signal. Behav. 2011, 6, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef]
- Janicka, M.; Reda, M.; Napieraj, N.; Kabała, K. Plant abiotic stress: Function of nitric oxide and hydrogen peroxide. In Nitric Oxide and Hydrogen Peroxide Signaling in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Swizerland, 2019; pp. 201–219. [Google Scholar]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef]
- Kaur, P.; Handa, N.; Verma, V.; Bakshi, P.; Kalia, R.; Sareen, S.; Nagpal, A.; Vig, A.; Mir, B.A.; Bhardwaj, R. Cross talk among reactive oxygen, nitrogen and sulfur during abiotic stress in plants. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 857–871. [Google Scholar]
- Hancock, J.T.; Whiteman, M. Hydrogen sulfide signaling: Interactions with nitric oxide and reactive oxygen species. Ann. N. Y. Acad. Sci. 2016, 1365, 5–14. [Google Scholar] [CrossRef]
- Biswas, M.S.; Fukaki, H.; Mori, I.C.; Nakahara, K.; Mano, J.I. Reactive oxygen species and reactive carbonyl species constitute a feed-forward loop in auxin signaling for lateral root formation. Plant J. 2019, 100, 536–548. [Google Scholar] [CrossRef]
- Li, J.-T.; Qiu, Z.-B.; Zhang, X.-W.; Wang, L.-S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta Physiol. Plant. 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Guo, Y.-K.; Bai, J.-G. Exogenous hydrogen peroxide changes antioxidant enzyme activity and protects ultrastructure in leaves of two cucumber ecotypes under osmotic stress. J. Plant Growth Regul. 2010, 29, 171–183. [Google Scholar] [CrossRef]
- Zhang, X.-L.; Jia, X.-F.; Yu, B.; Gao, Y.; Bai, J.-G. Exogenous hydrogen peroxide influences antioxidant enzyme activity and lipid peroxidation in cucumber leaves at low light. Sci. Hortic. 2011, 129, 656–662. [Google Scholar] [CrossRef]
- Nawaz, F.; Majeed, S.; Ahmad, K.S.; Aqib, M.; Shehzad, M.A.; Aurangzaib, M.; Shahbaz, M. Reactive sulfur species-key regulators of abiotic stress tolerance in plants. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 685–713. [Google Scholar]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.B.; Al Mahmud, J.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Bhuyan, M.B.; Hasanuzzaman, M.; Parvin, K.; Mohsin, S.M.; Al Mahmud, J.; Nahar, K.; Fujita, M. Nitric oxide and hydrogen sulfide: Two intimate collaborators regulating plant defense against abiotic stress. Plant Growth Regul. 2020, 1–16. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhuyan, M.B.; Oku, H.; Fujita, M. Exogenous nitric oxide pretreatment protects Brassica napus L. seedlings from paraquat toxicity through the modulation of antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2018, 126, 173–186. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- González, A.; de Los Ángeles Cabrera, M.; Henríquez, M.J.; Contreras, R.A.; Morales, B.; Moenne, A. Cross talk among calcium, hydrogen peroxide, and nitric oxide and activation of gene expression involving calmodulins and calcium-dependent protein kinases in Ulva compressa exposed to copper excess. Plant Physiol. 2012, 158, 1451–1462. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, M.; Zhang, J.; Ding, H.; Xu, S.; Hu, X.; Tan, M. Nitric oxide induced by hydrogen peroxide mediates abscisic acid-induced activation of the mitogen-activated protein kinase cascade involved in antioxidant defense in maize leaves. New Phytol. 2007, 175, 36–50. [Google Scholar] [CrossRef]
- Qiao, W.; Li, C.; Fan, L.-M. Cross-talk between nitric oxide and hydrogen peroxide in plant responses to abiotic stresses. Environ. Exp. Bot. 2014, 100, 84–93. [Google Scholar] [CrossRef]
- Palma, J.M.; Gupta, D.K.; Corpas, F.J. Hydrogen peroxide and nitric oxide generation in plant cells: Overview and queries. In Nitric Oxide and Hydrogen Peroxide Signaling in Higher Plants; Springer: Cham, Swizerland, 2019; pp. 1–16. [Google Scholar]
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Production sites of reactive oxygen species (ROS) in organelles from plant cells. In Reactive Oxygen Species and Oxidative Damage in Plants under Stress; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer: Cham, Swizerland, 2015; pp. 1–22. [Google Scholar]
- Corpas, F.J.; Barroso, J.B.; Palma, J.M.; Rodriguez-Ruiz, M. Plant peroxisomes: A nitro-oxidative cocktail. Redox Biol. 2017, 11, 535–542. [Google Scholar] [CrossRef]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. In Proceedings of the National Academy of Sciences of the United States of America, Los Angeles, CA, USA, 12 January 2004; pp. 4003–4008. [Google Scholar]
- Liu, D.; Liu, M.; Liu, X.-L.; Cheng, X.-G.; Liang, Z.-W. Silicon priming created an enhanced tolerance in alfalfa (Medicago sativa L.) seedlings in response to high alkaline stress. Front. Plant Sci. 2018, 9, 716. [Google Scholar] [CrossRef]
- Ortega-Galisteo, A.P.; Rodríguez-Serrano, M.; Pazmiño, D.M.; Gupta, D.K.; Sandalio, L.M.; Romero-Puertas, M. S-Nitrosylated proteins in pea (Pisum sativum L.) leaf peroxisomes: Changes under abiotic stress. J. Exp. Bot. 2012, 63, 2089–2103. [Google Scholar] [CrossRef] [PubMed]
- Gruhlke, M.C. Reactive sulfur species: A new player in plant physiology? In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 715–728. [Google Scholar]
- Hill, B.G.; Bhatnagar, A. Protein S-glutathiolation: Redox-sensitive regulation of protein function. J. Mol. Cell. Cardiol. 2012, 52, 559–567. [Google Scholar] [CrossRef]
- Tao, L.; English, A.M. Protein S-glutathiolation triggered by decomposed S-nitrosoglutathione. Biochemistry 2004, 43, 4028–4038. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Jiménez-Moreno, N.; Dias, I.H.; Debelec, B.; Vucetic, M.; Fladmark, K.E.; Basaga, H.; Ribaric, S.; Milisav, I.; Cuadrado, A. Redox control of protein degradation. Redox Biol. 2015, 6, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jia, H.; Wang, J.; Cao, Q.; Wen, Z. Hydrogen sulfide is involved in maintaining ion homeostasis via regulating plasma membrane Na+/H+ antiporter system in the hydrogen peroxide-dependent manner in salt-stress Arabidopsis thaliana root. Protoplasma 2014, 251, 899–912. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef]
- Gupta, V.; Carroll, K.S. Sulfenic acid chemistry, detection and cellular lifetime. Biochim. Biophys. Acta 2014, 1840, 847–875. [Google Scholar] [CrossRef]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J. Redox chemistry and chemical biology of H2S, hydropersulfides, and derived species: Implications of their possible biological activity and utility. Free Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef]
- Akaike, T.; Ida, T.; Wei, F.-Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Mano, J.I.; Biswas, M.; Sugimoto, K. Reactive carbonyl species: A missing link in ROS signaling. Plants 2019, 8, 391. [Google Scholar] [CrossRef]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I. The roles of reactive carbonyl species in induction of antioxidant defence and ROS signalling in extreme halophytic model Eutrema parvulum and glycophytic model Arabidopsis thaliana. Environ. Exp. Bot. 2019, 160, 81–91. [Google Scholar] [CrossRef]
- Mano, J.I.; Kanameda, S.; Kuramitsu, R.; Matsuura, N.; Yamauchi, Y. Detoxification of reactive carbonyl species by glutathione transferase Tau isozymes. Front. Plant Sci. 2019, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Ye, W.; Matsushima, D.; Rhaman, M.S.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.S.; Mano, J.I.; Murata, Y. Reactive carbonyl species function as signal mediators downstream of H2O2 production and regulate [Ca2+] cyt elevation in ABA signal pathway in Arabidopsis guard cells. Plant Cell Physiol. 2019, 60, 1146–1159. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Glyoxalase and methylglyoxal as biomarkers for plant stress tolerance. Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Saito, R.; Yamamoto, H.; Makino, A.; Sugimoto, T.; Miyake, C. Methylglyoxal functions as Hill oxidant and stimulates the photoreduction of O2 at photosystem I: A symptom of plant diabetes. Plant Cell Environ. 2011, 34, 1454–1464. [Google Scholar] [CrossRef]
- Hao, Q.; Maret, W. Aldehydes release zinc from proteins. A pathway from oxidative stress/lipid peroxidation to cellular functions of zinc. FEBS J. 2006, 273, 4300–4310. [Google Scholar] [CrossRef]
- Kiranmai, K.; Lokanadha Rao, G.; Pandurangaiah, M.; Nareshkumar, A.; Amaranatha Reddy, V.; Lokesh, U.; Venkatesh, B.; Anthony Johnson, A.; Sudhakar, C. A novel WRKY transcription factor, MuWRKY3 (Macrotyloma uniflorum Lam. Verdc.) enhances drought stress tolerance in transgenic groundnut (Arachis hypogaea L.) plants. Front. Plant Sci. 2018, 9, 346. [Google Scholar] [CrossRef]
- Sun, X.; Wang, P.; Jia, X.; Huo, L.; Che, R.; Ma, F. Improvement of drought tolerance by overexpressing MdATG18a is mediated by modified antioxidant system and activated autophagy in transgenic apple. Plant Biotechnol. J. 2018, 16, 545–557. [Google Scholar] [CrossRef]
- Wang, K.; Zhong, M.; Wu, Y.-h.; Bai, Z.-y.; Liang, Q.-y.; Liu, Q.-l.; Pan, Y.-z.; Zhang, L.; Jiang, B.-b.; Jia, Y. Overexpression of a chrysanthemum transcription factor gene DgNAC1 improves the salinity tolerance in chrysanthemum. Plant Cell Rep. 2017, 36, 571–581. [Google Scholar] [CrossRef]
- Shafi, A.; Pal, A.K.; Sharma, V.; Kalia, S.; Kumar, S.; Ahuja, P.S.; Singh, A.K. Transgenic potato plants overexpressing SOD and APX exhibit enhanced lignification and starch biosynthesis with improved salt stress tolerance. Plant Mol. Biol. Rep. 2017, 35, 504–518. [Google Scholar] [CrossRef]
- Sapara, K.K.; Khedia, J.; Agarwal, P.; Gangapur, D.R.; Agarwal, P.K. SbMYB15 transcription factor mitigates cadmium and nickel stress in transgenic tobacco by limiting uptake and modulating antioxidative defence system. Funct. Plant Biol. 2019, 46, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dubey, A.K.; Kumar, V.; Ansari, M.A.; Narayan, S.; Kumar, S.; Pandey, V.; Shirke, P.A.; Pande, V.; Sanyal, I. Over-expression of chickpea glutaredoxin (CaGrx) provides tolerance to heavy metals by reducing metal accumulation and improved physiological and antioxidant defence system. Ecotoxicol. Environ. Saf. 2020, 192, 110252. [Google Scholar] [CrossRef] [PubMed]
- Karkute, S.; Krishna, R.; Ansari, W.; Singh, B.; Singh, P.; Singh, M.; Singh, A. Heterologous expression of the AtDREB1A gene in tomato confers tolerance to chilling stress. Biol. Plant. 2019, 63, 268–277. [Google Scholar] [CrossRef]
- Che, Y.; Zhang, N.; Zhu, X.; Li, S.; Wang, S.; Si, H. Enhanced tolerance of the transgenic potato plants overexpressing Cu/Zn superoxide dismutase to low temperature. Sci. Hortic. 2020, 261, 108949. [Google Scholar] [CrossRef]
- Wang, L.; Gao, J.; Zhang, Z.; Liu, W.; Cheng, P.; Mu, W.; Su, T.; Chen, S.; Chen, F.; Jiang, J. Overexpression of CmSOS1 confers waterlogging tolerance in Chrysanthemum. J. Integr. Plant Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Hou, L.; Xiao, P.; Guo, Y.; Deyholos, M.K.; Liu, X. VvWRKY30, a grape WRKY transcription factor, plays a positive regulatory role under salinity stress. Plant Sci. 2019, 280, 132–142. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, N.; Yang, J.; Guo, H.; Liu, Z.; Zheng, X.; Li, S.; Xiang, F. The salt-induced transcription factor GmMYB84 confers salinity tolerance in soybean. Plant Sci. 2020, 291, 110326. [Google Scholar] [CrossRef]
- Duan, F.; Ding, J.; Lee, D.; Lu, X.; Feng, Y.; Song, W. Overexpression of SoCYP85A1, a spinach cytochrome p450 gene in transgenic tobacco enhances root development and drought stress tolerance. Front. Plant Sci. 2017, 8, 1909. [Google Scholar] [CrossRef]
- Xia, Z.; Xu, Z.; Wei, Y.; Wang, M. Overexpression of the maize sulfite oxidase increases sulfate and GSH levels and enhances drought tolerance in transgenic tobacco. Front. Plant Sci. 2018, 9, 298. [Google Scholar] [CrossRef]
- Dubey, A.K.; Kumar, N.; Kumar, A.; Ansari, M.A.; Ranjan, R.; Gautam, A.; Sahu, N.; Pandey, V.; Behera, S.K.; Mallick, S. Over-expression of CarMT gene modulates the physiological performance and antioxidant defense system to provide tolerance against drought stress in Arabidopsis thaliana L. Ecotoxicol. Environ. Saf. 2019, 171, 54–65. [Google Scholar] [CrossRef]
- Tan, Y.; Wang, L. MpDGK2, a novel diacylglycerol kinase from Malus prunifolia, confers drought stress tolerance in transgenic Arabidopsis. Plant Mol. Biol. Rep. 2020, 1–9. [Google Scholar] [CrossRef]
- Kumar, S.; Khare, R.; Trivedi, P.K. Arsenic-responsive high-affinity rice sulphate transporter, OsSultr1;1, provides abiotic stress tolerance under limiting sulphur condition. J. Hazard. Mater. 2019, 373, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, W.; Wang, W.; Zhang, G.; Liu, Y.; Wang, Y.; Wang, W. Wheat F-box protein gene TaFBA1 is involved in plant tolerance to heat stress. Front. Plant Sci. 2018, 9, 521. [Google Scholar] [CrossRef]
- Huo, L.; Sun, X.; Guo, Z.; Jia, X.; Che, R.; Sun, Y.; Zhu, Y.; Wang, P.; Gong, X.; Ma, F. MdATG18a overexpression improves basal thermotolerance in transgenic apple by decreasing damage to chloroplasts. Hortic. Res. 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Lv, Y.; Fu, S.; Chen, S.; Zhang, W.; Qi, C. Ethylene response factor BnERF2-like (ERF2. 4) from Brassica napus L. enhances submergence tolerance and alleviates oxidative damage caused by submergence in Arabidopsis thaliana. Crop J. 2016, 4, 199–211. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Tripathi, V.; Shukla, R.K. MaRAP2-4, a waterlogging-responsive ERF from Mentha, regulates bidirectional sugar transporter AtSWEET10 to modulate stress response in Arabidopsis. Plant Biotechnol. J. 2018, 16, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Guo, B.; Shen, H.; Pan, Y.; Hong, Y.; Lv, C.; Xu, R. Overexpression of barley transcription factor HvERF2.11 in Arabidopsisenhances plant waterlogging tolerance. Int. J. Mol. Sci. 2020, 21, 1982. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).