
Low NETosis Induced in Anaplasma phagocytophilum-Infected Cells


 ,
,  , ,
, ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.2. HL60 Cell Culture
2.3. Anaplasma phagocytophilum-Infected HL60 Cells and Bacteria Isolation
2.4. HL60 Differentiation and NETs Stimulation
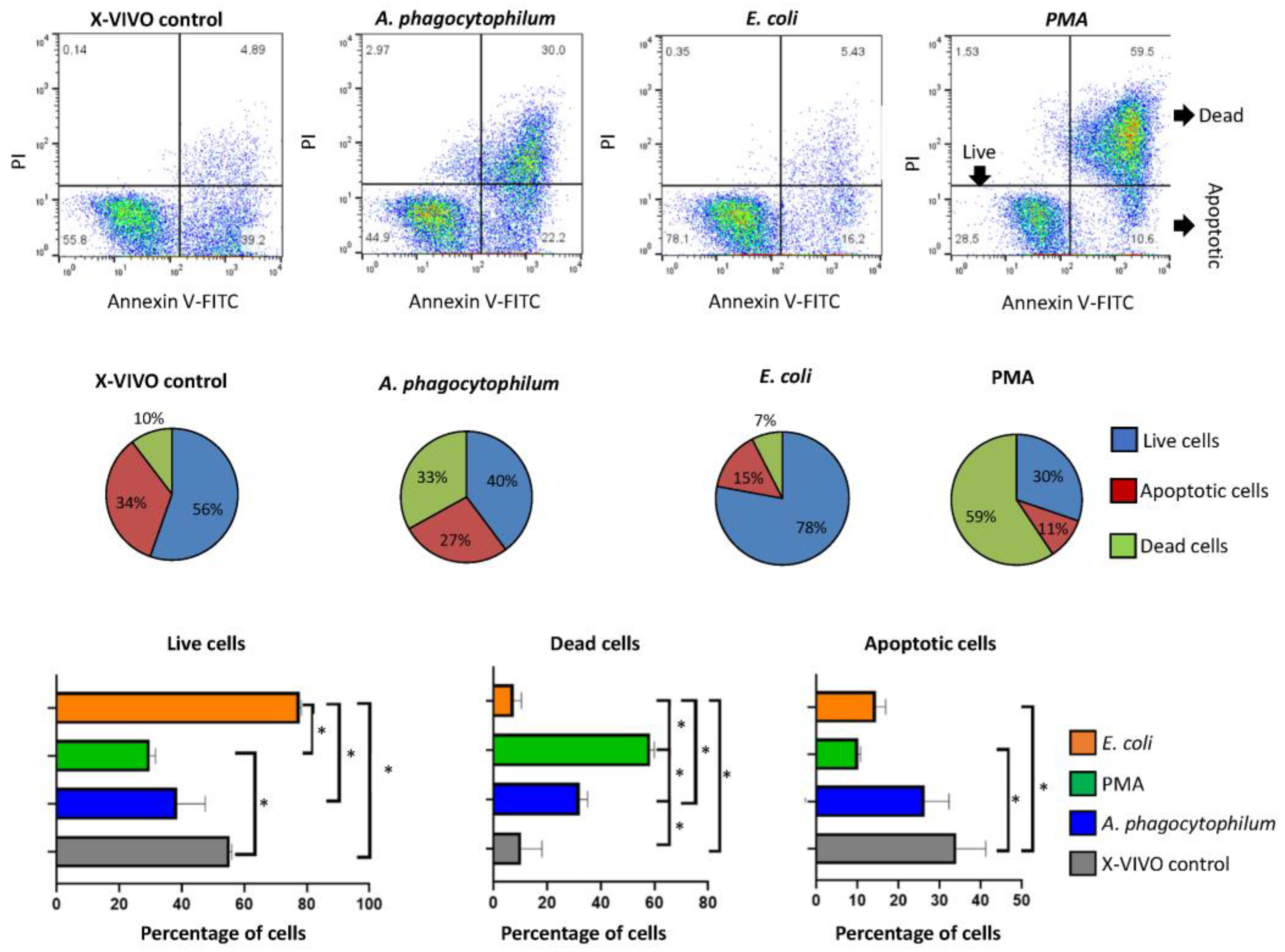
2.5. Annexin V-FITC Staining to Detect Cell Apoptosis
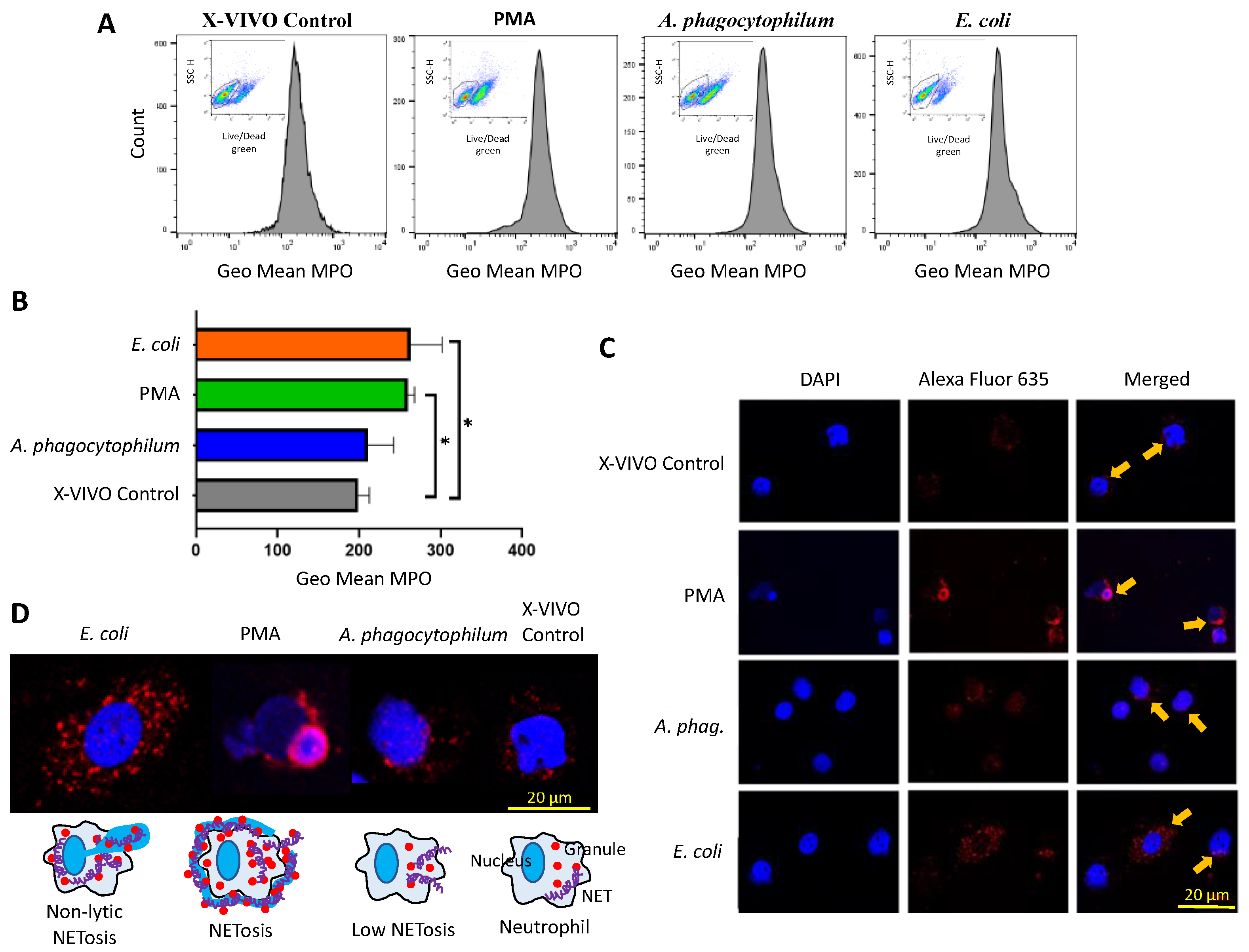
2.6. Live/Dead Cells and MPO Quantification by Flow Cytometry
2.7. Assessment of HL60 Differentiation by Flow Cytometry
2.8. Immunofluorescence Assay (IFA) of HL60 Cells
2.9. Determination of A. phagocytophilum DNA Levels by qRT-PCR
3. Results and Discussion
3.1. Bioinformatics Analysis Showed Overactivation of NETosis-Related Proteins in Infected Human HL60 Cells
3.2. Anaplasma phagocytophilum and E. coli Differently Affect Differentiated HL60 Cells
3.3. A. phagocytophilum Does Not Increase MPO Expression, and thus, NETs Formation in Differentiated HL60 Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Villar, M.; López, V.; Ayllón, N.; Cabezas-Cruz, A.; López, J.A.; Vázquez, J.; Alberdi, P.; de la Fuente, J. The intracellular bacterium Anaplasma phagocytophilum selectively manipulates the levels of vertebrate host proteins in the tick vector Ixodes scapularis. Parasites Vectors 2016, 9, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas-Jerónimo, S.; Villar, M.; Cabezas-Cruz, A.; Valdés, J.J.; Estrada-Peña, A.; Alberdi, P.; De la Fuente, J. Functional evolution of subolesin/akirin. Front. Physiol. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T. Biosynthesis and biology of mammalian GPI-anchored proteins. Open Biol. 2020, 10, 190290. [Google Scholar] [CrossRef] [Green Version]
- Dumler, J.S.; Lichay, M.; Chen, W.-H.; Rennoll-Bankert, K.E.; Park, J.-h. Anaplasma phagocytophilum Activates NF-κB Signaling via Redundant Pathways. Front. Public Health 2020, 8, 558283. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Villar, M.; Cabezas-Cruz, A.; Estrada-Pena, A.; Ayllon, N.; Alberdi, P. Tick–host–pathogen interactions: Conflict and cooperation. PLoS Pathog. 2016, 12, e1005488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Amulic, B.; Hayes, G. Neutrophil extracellular traps. Curr. Biol. 2011, 21, R297–R298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gao, F.; Wang, Q.; Wang, K.; Pan, S.; Pan, Z.; Xu, S.; Li, L.; Zhao, D. Differentiation of HL-60 cells in serum-free hematopoietic cell media enhances the production of neutrophil extracellular traps. Exp. Ther. Med. 2021, 21, 353. [Google Scholar] [CrossRef]
- Metzler, K.D.; Fuchs, T.A.; Nauseef, W.M.; Reumaux, D.; Roesler, J.; Schulze, I.; Wahn, V.; Papayannopoulos, V.; Zychlinsky, A. Myeloperoxidase is required for neutrophil extracellular trap formation: Implications for innate immunity. Blood J. Am. Soc. Hematol. 2011, 117, 953–959. [Google Scholar] [CrossRef]
- Peña, A.E.; Villar, M.; Jerónimo, S.A.; López, V.; Alberdi, P.; Cabezas-Cruz, A.; De La Fuente, J. Use of Graph Theory to Characterize Human and Arthropod Vector Cell Protein Response to Infection with Anaplasma phagocytophilum. Front. Cell. Infect. Microbiol. 2018, 8, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g: Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, M.; von Mering, C.; Campillos, M.; Jensen, L.J.; Bork, P. STITCH: Interaction networks of chemicals and proteins. Nucleic Acids Res. 2007, 36, D684–D688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karki, P.; Ijdo, J.W. Anaplasma phagocytophilum infection induces apoptosis in HL-60 cells. World J. Microbiol. Biotechnol. 2011, 27, 2741–2746. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, J.; Ayoubi, P.; Blouin, E.F.; Almazán, C.; Naranjo, V.; Kocan, K.M. Gene expression profiling of human promyelocytic cells in response to infection with Anaplasma phagocytophilum. Cell Microbiol. 2004, 7, 549–559. [Google Scholar] [CrossRef]
- Asanovich, K.M.; Bakken, J.S.; Madigan, J.E.; Aguero-Rosenfeld, M.; Wormser, G.P.; Dumler, J.S. Antigenic diversity of granulocytic Ehrlichia isolates from humans in Wisconsin and New York and a horse in California. J. Infect. Dis. 1997, 176, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Reppert, E.; Galindo, R.C.; Ayllón, N.; Breshears, M.A.; Kocan, K.M.; Blouin, E.F.; de la Fuente, J. Studies of Anaplasma phagocytophilum in sheep experimentally infected with the human NY-18 isolate: Characterization of tick feeding sites. Ticks Tick Borne Dis. 2014, 5, 744–752. [Google Scholar] [CrossRef]
- Ayllón, N.; Villar, M.; Busby, A.T.; Kocan, K.M.; Blouin, E.F.; Bonzón-Kulichenko, E.; Galindo, R.C.; Mangold, A.J.; Alberdi, P.; de la Lastra, J.M.P.; et al. Anaplasma phagocytophilum Inhibits Apoptosis and Promotes Cytoskeleton Rearrangement for Infection of Tick Cells. Infect. Immun. 2013, 81, 2415–2425. [Google Scholar] [CrossRef] [Green Version]
- Lehman, H.K.; Segal, B.H. The role of neutrophils in host defense and disease. J. Allergy Clin. Immunol. 2020, 145, 1535–1544. [Google Scholar] [CrossRef]
- Notarangelo, L.D.; Badolato, R. Leukocyte trafficking in primary immunodeficiencies. J. Leukoc. Biol. 2009, 85, 335–343. [Google Scholar] [CrossRef]
- Paolillo., R.; Boulanger, M.; Gâtel, P.; Gabellier, L.; De Toledo, M.; Tempé, D.; Hallal, R.; Akl, D.; Moreaux, J.; Baik, H.; et al. The NADPH oxidase NOX2 is a marker of adverse prognosis involved in chemoresistance of acute myeloid leukemias. Haematologica 2022. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Droste, A.; Chaves, G.; Stein, S.; Trzmiel, A.; Schweizer, M.; Karl, H.; Musset, B. Zinc accelerates respiratory burst termination in human PMN. Redox Biol. 2021, 47, 102133. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, H.; Genagon, S.A.; Russo, T.A. Extraintestinal Pathogenic Escherichia coli Survives within Neutrophils. Infect. Immun. 2007, 75, 2776–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente, J.; Peña, A.E.; Cabezas-Cruz, A.; Kocan, K.M. Anaplasma phagocytophilum Uses Common Strategies for Infection of Ticks and Vertebrate Hosts. Trends Microbiol. 2016, 24, 173–180. [Google Scholar] [CrossRef]
- Artigas-Jerónimo, S.; Alberdi, P.; Rayo, M.V.; Cabezas-Cruz, A.; Prados, P.J.E.; Mateos-Hernández, L.; De La Fuente, J. Anaplasma phagocytophilum modifies tick cell microRNA expression and upregulates isc-mir-79 to facilitate infection by targeting the Roundabout protein 2 pathway. Sci. Rep. 2019, 9, 9073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas-Jerónimo, S.; Estrada-Peña, A.; Cabezas-Cruz, A.; Alberdi, P.; Villar, M.; De La Fuente, J. Modeling Modulation of the Tick Regulome in Response to Anaplasma phagocytophilum for the Identification of New Control Targets. Front. Physiol. 2019, 10, 462. [Google Scholar] [CrossRef]
- Feng, L.; Han, W.; Wang, Q.; Bastin, D.A.; Wang, L. Characterization of Escherichia coli O86 O-antigen gene cluster and identification of O86-specific genes. Vet. Microbiol. 2005, 106, 241–248. [Google Scholar] [CrossRef]
- Li, M.; Shen, J.; Liu, X.; Shao, J.; Yi, W.; Chow, C.S.; Wang, P.G. Identification of a new α1, 2-fucosyltransferase involved in O-antigen biosynthesis of Escherichia coli O86: B7 and formation of H-type 3 blood group antigen. Biochemistry 2008, 47, 11590–11597. [Google Scholar] [CrossRef]
- Soares, M.P.; Yilmaz, B. Microbiota Control of Malaria Transmission. Trends Parasitol. 2016, 32, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Antal-Szalmas, P.; Van Strijp, J.A.G.; Weersink, A.J.L.; Verhoef, J.; Van Kessel, K.P.M. Quantitation of surface CD14 on human monocytes and neutrophils. J. Leukoc. Biol. 1997, 61, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Farahvash, A.; Douda, D.N.; Licht, J.-C.; Grasemann, H.; Sweezey, N.; Palaniyar, N. JNK Activation Turns on LPS- and Gram-Negative Bacteria-Induced NADPH Oxidase-Dependent Suicidal NETosis. Sci. Rep. 2017, 7, 3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Philip, L.M.; Cheung, G.; Vadakepeedika, S.; Grasemann, H.; Sweezey, N.; Palaniyar, N. Regulating NETosis: Increasing pH Promotes NADPH Oxidase-Dependent NETosis. Front. Med. 2018, 5, 19. [Google Scholar] [CrossRef]
- Lam, G.Y.; Huang, J.; Brumell, J.H. The Many Roles of NOX2 NADPH Oxidase-Derived ROS in Immunity; Springer: New York, NY, USA, 2010; pp. 415–430. [Google Scholar]
- Alberdi, P.; Espinosa, P.J.; Cabezas-Cruz, A.; De La Fuente, J. Anaplasma phagocytophilum Manipulates Host Cell Apoptosis by Different Mechanisms to Establish Infection. Vet. Sci. 2016, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.M.; Herron, M.J.; Wang, X.-R.; Baldridge, G.D.; Oliver, J.D.; Munderloh, U.G. Global Transcription Profiles of Anaplasma phagocytophilum at Key Stages of Infection in Tick and Human Cell Lines and Granulocytes. Front. Vet. Sci. 2020, 7, 111. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artigas-Jerónimo, S.; González-García, A.; de la Fuente, J.; Blanda, V.; Shekarkar Azgomi, M.; Villar, M.; Mohammadnezhad, L.; Grippi, F.; Torina, A.; Sireci, G. Low NETosis Induced in Anaplasma phagocytophilum-Infected Cells. Vaccines 2022, 10, 1756. https://doi.org/10.3390/vaccines10101756
Artigas-Jerónimo S, González-García A, de la Fuente J, Blanda V, Shekarkar Azgomi M, Villar M, Mohammadnezhad L, Grippi F, Torina A, Sireci G. Low NETosis Induced in Anaplasma phagocytophilum-Infected Cells. Vaccines. 2022; 10(10):1756. https://doi.org/10.3390/vaccines10101756
Chicago/Turabian StyleArtigas-Jerónimo, Sara, Almudena González-García, José de la Fuente, Valeria Blanda, Mojtaba Shekarkar Azgomi, Margarita Villar, Leila Mohammadnezhad, Francesca Grippi, Alessandra Torina, and Guido Sireci. 2022. "Low NETosis Induced in Anaplasma phagocytophilum-Infected Cells" Vaccines 10, no. 10: 1756. https://doi.org/10.3390/vaccines10101756