

An IgY Effectively Prevents Goslings from Virulent GAstV Infection
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus
2.2. Establishment of Challenge Model
2.3. Preparation of Inactivated Virus
2.4. Immunization of Laying Hens
2.5. Assessment of Specific IgY Titers
2.6. Preparation of IgY Antibody
2.7. Virus Neutralization Assay (VN)
2.8. Protection of Goslings Inoculated with GAstV
2.8.1. Determination of Minimum Effective Dose
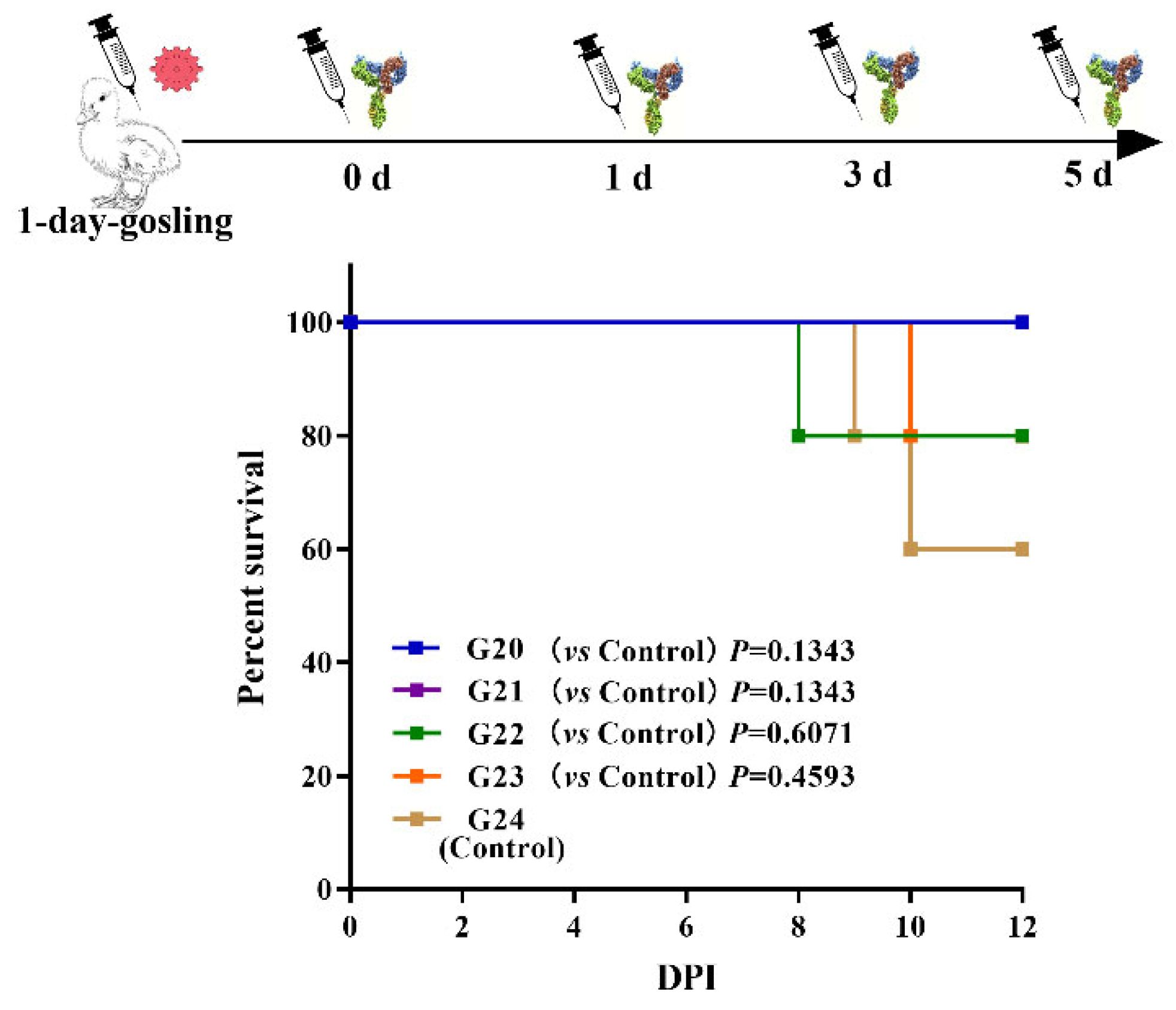
2.8.2. Estimation of Prophylactic Antiviral Treatment and Post-Infection Treatment
2.9. qRT-PCR Analysis to Determine Viral Loads
2.10. Statistical Analysis
3. Results
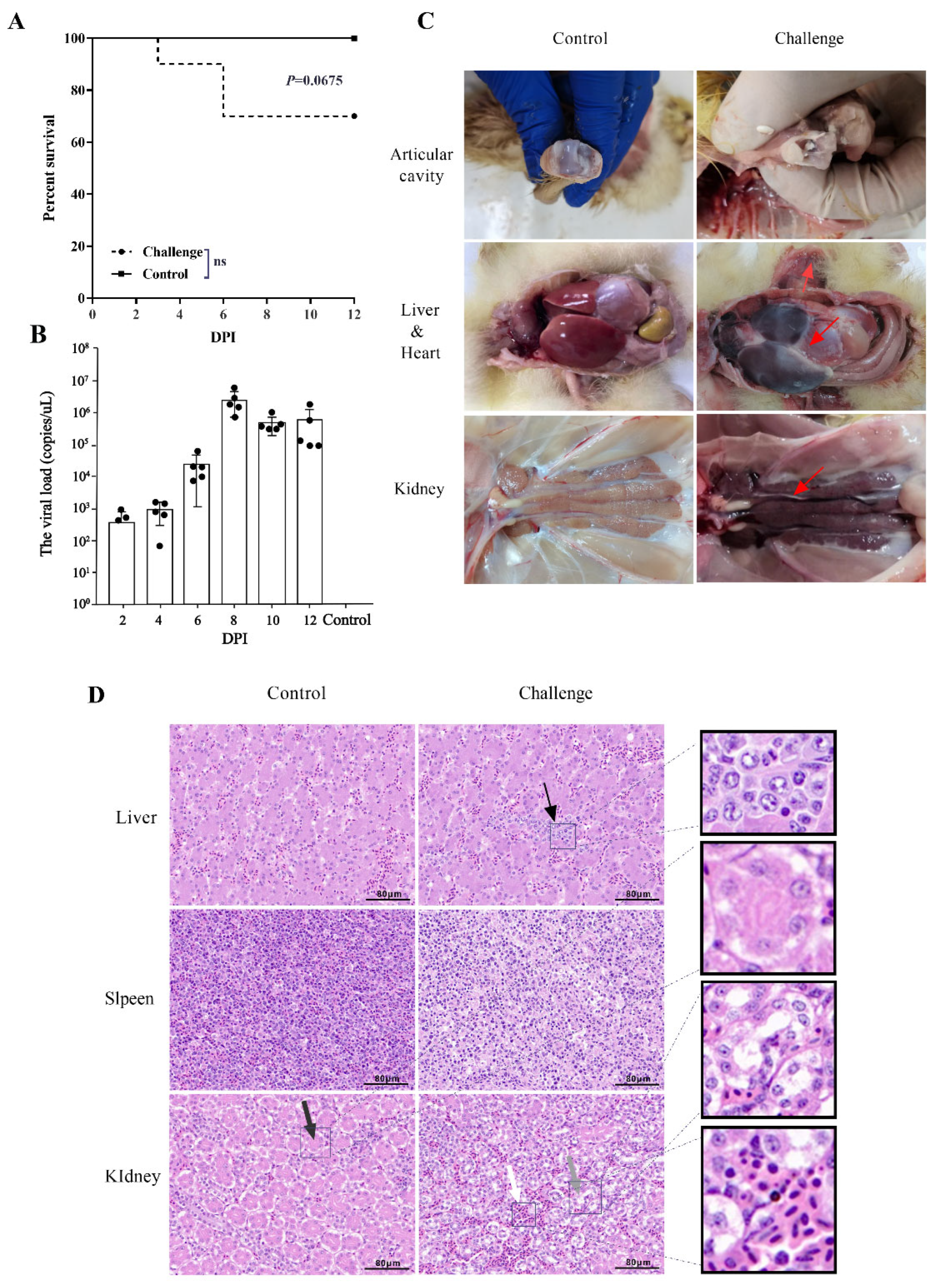
3.1. Phylogenetic and Pathogenicity Analysis of GAstV-AHQJ18
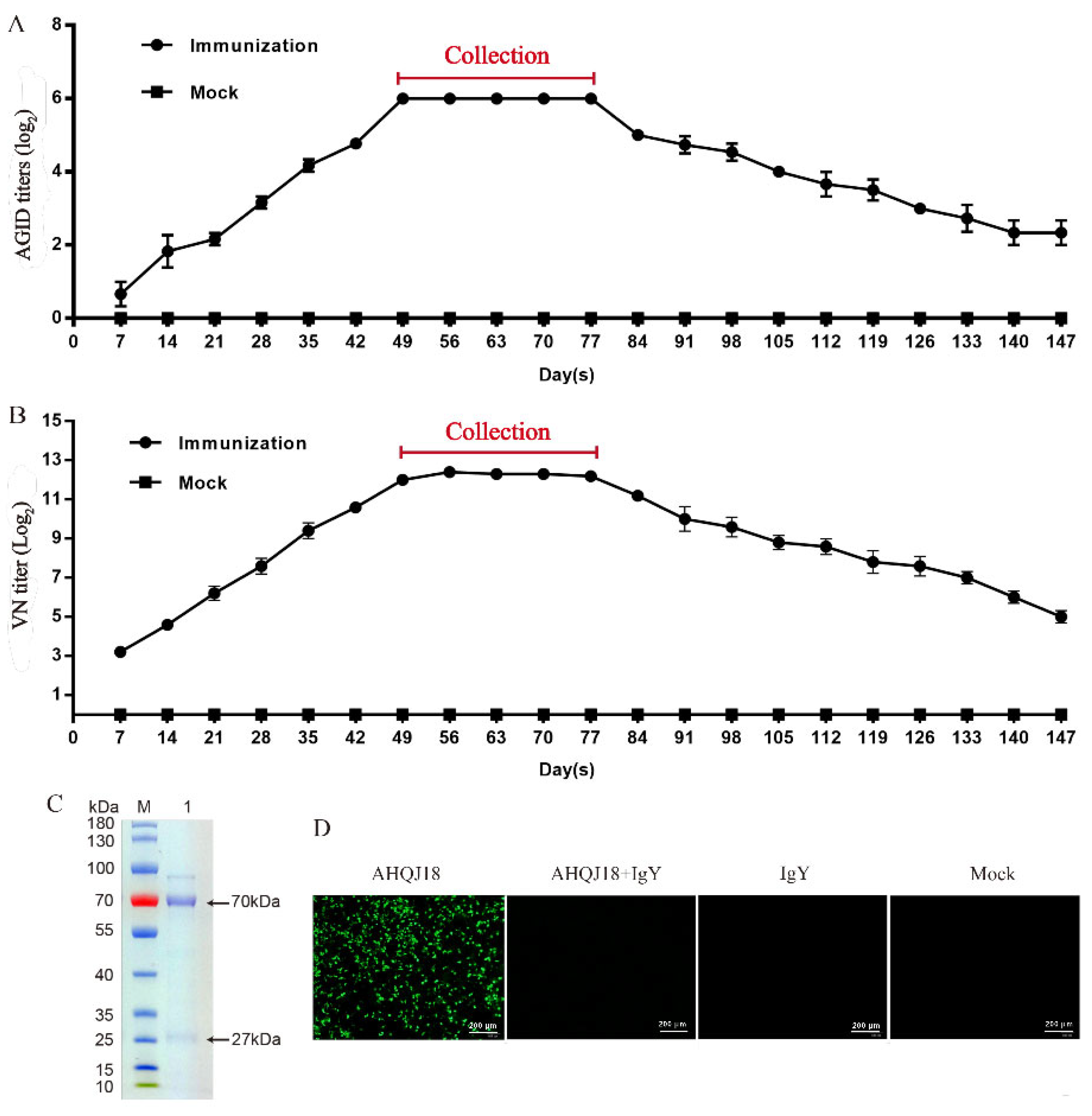
3.2. Determination of IgY Titer against GAstV
3.3. Protection of Goslings Inoculated with Anti-GAstV IgY
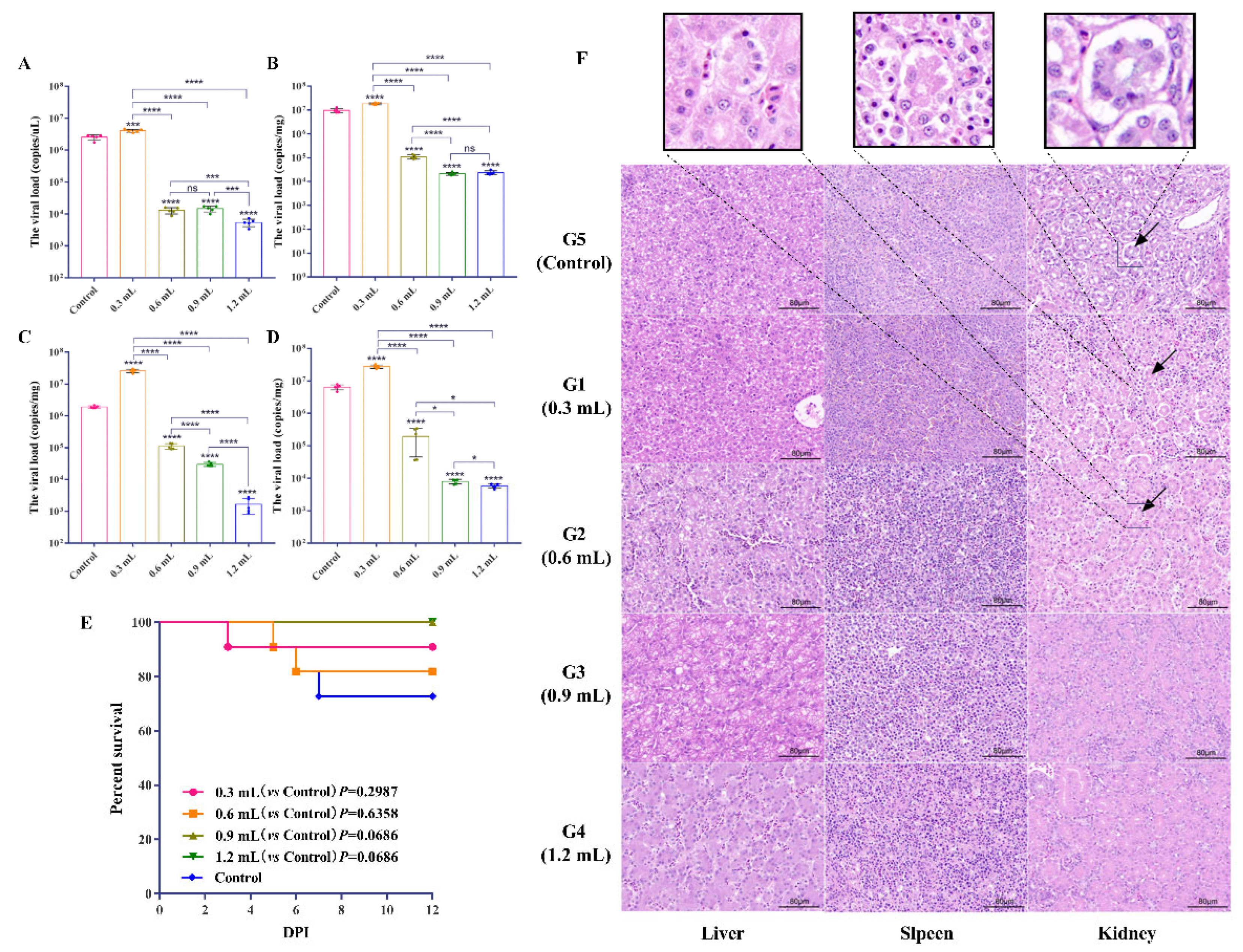
3.3.1. Determination of Minimum Protective Dose of IgY
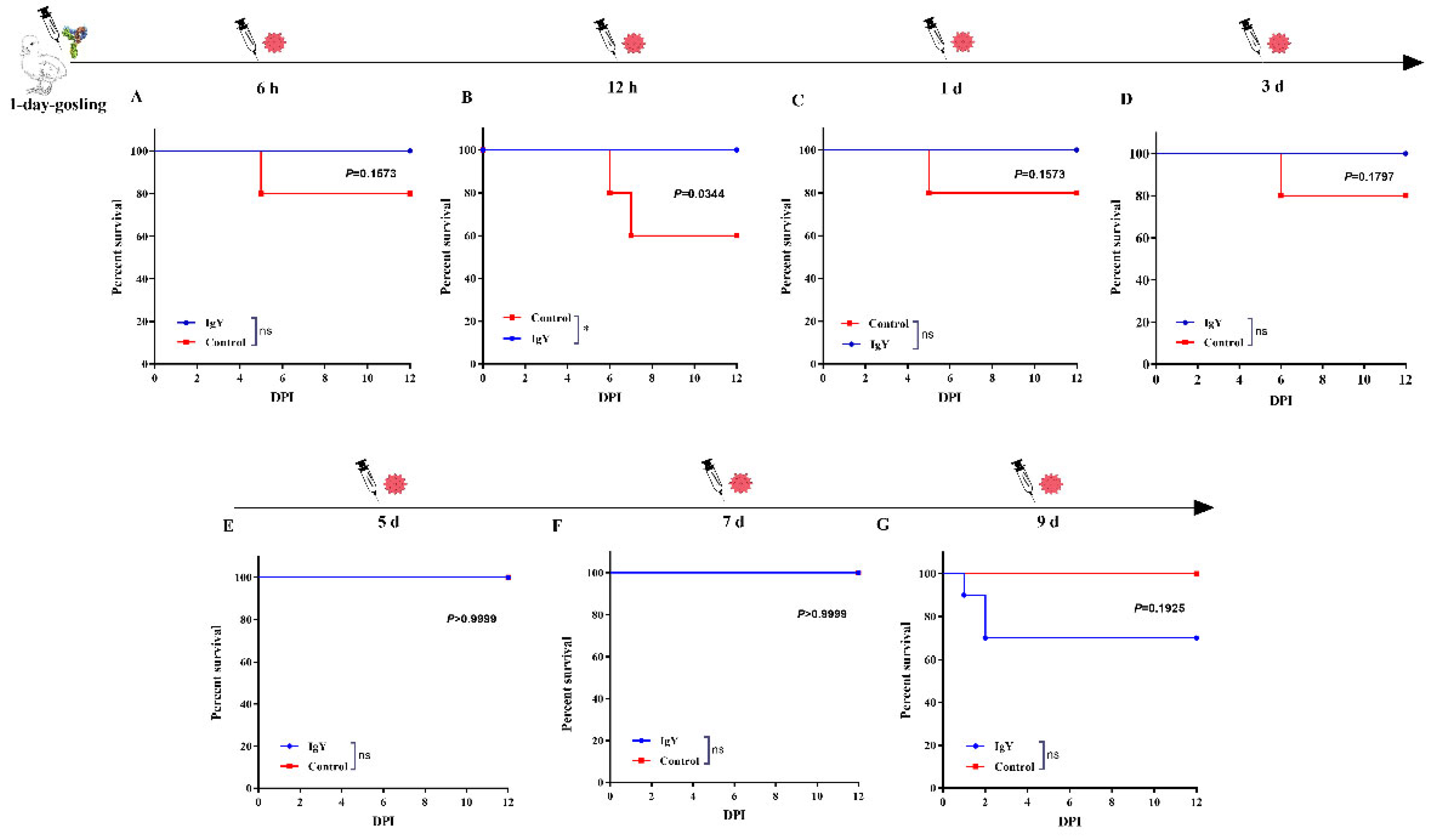
3.3.2. Prophylactic and Curative Effect of IgY against GAstV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Qin, Y.; Fang, Q.; Liu, H.; Ji, C.; Chen, Y.; Ouyang, K.; Wei, Z.; Huang, W. Construction of a reverse genetic system for porcine astrovirus. Arch. Virol. 2018, 163, 1511–1518. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, D.; Li, T.; Zhou, H.; Liu, X.; Wang, X.; Lu, H.; Gao, W.; Wang, Y.; Zou, X.; et al. An emerging novel goose astrovirus associated with gosling gout disease, China. Emerg. Microbes Infect. 2018, 7, 152. [Google Scholar] [CrossRef]
- Wang, A.P.; Zhang, S.; Xie, J.; Gu, L.L.; Wu, S.; Wu, Z.; Liu, L.; Feng, Q.; Dong, H.Y.; Zhu, S.Y. Isolation and characterization of a goose astrovirus 1 strain causing fatal gout in goslings, China. Poult. Sci. 2021, 100, 101432. [Google Scholar] [CrossRef]
- Zhu, Q.; Sun, D. Goose Astrovirus in China: A Comprehensive Review. Viruses 2022, 14, 1759. [Google Scholar] [CrossRef]
- Roach, S.N.; Langlois, R.A. Intra- and Cross-Species Transmission of Astroviruses. Viruses 2021, 13, 1127. [Google Scholar] [CrossRef]
- Li, J.Y.; Hu, W.Q.; Liu, T.N.; Zhang, H.H.; Opriessnig, T.; Xiao, C.T. Isolation and evolutionary analyses of gout-associated goose astrovirus causing disease in experimentally infected chickens. Poult. Sci. 2021, 100, 543–552. [Google Scholar] [CrossRef]
- Wei, F.; Yang, J.; Wang, Y.; Chen, H.; Diao, Y.; Tang, Y. Isolation and characterization of a duck-origin goose astrovirus in China. Emerg. Microbes Infect. 2020, 9, 1046–1054. [Google Scholar] [CrossRef]
- Michael, A.; Meenatchisundaram, S.; Parameswari, G.; Subbraj, T.; Selvakumaran, R.; Ramalingam, S. Chicken egg yolk antibodies (IgY) as an alternative to mammalian antibodies. Indian J. Sci. Technol. 2010, 3, 468–474. [Google Scholar] [CrossRef]
- Krief, A.; Letesson, J.-J.; Billen, D. Comparison between ‘IgY technology’from chickens and ‘IgG technology’from mice for production of tailor-made antibodies. Tetrahedron Lett. 2002, 43, 1843–1846. [Google Scholar] [CrossRef]
- Pérez de la Lastra, J.M.; Baca-González, V.; Asensio-Calavia, P.; González-Acosta, S.; Morales-delaNuez, A. Can immunization of hens provide oral-based therapeutics against COVID-19? Vaccines 2020, 8, 486. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Jin, L.; Zhen, Y.; Lu, Y.; Li, S.; You, J.; Wang, L. Application of chicken egg yolk immunoglobulins in the control of terrestrial and aquatic animal diseases: A review. Biotechnol. Adv. 2011, 29, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Handharyani, E.; Sari, D.K.; Takama, K.; Fukuda, K.; Endo, I.; Yamamoto, R.; Sawa, M.; Tanaka, M.; Konishi, I. Development of neutralization antibodies against highly pathogenic H5N1 avian influenza virus using ostrich (Struthio camelus) yolk. Mol. Med. Rep. 2008, 1, 203–209. [Google Scholar] [PubMed]
- Nguyen, H.H.; Tumpey, T.M.; Park, H.J.; Byun, Y.H.; Tran, L.D.; Nguyen, V.D.; Kilgore, P.E.; Czerkinsky, C.; Katz, J.M.; Seong, B.L.; et al. Prophylactic and therapeutic efficacy of avian antibodies against influenza virus H5N1 and H1N1 in mice. PLoS ONE 2010, 5, e10152. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Aldous, E.W.; Fuller, C.M. The long view: A selective review of 40 years of Newcastle disease research. Avian Pathol. 2012, 41, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Box, P.; Stedman, R.; Singleton, L. Newcastle Disease: I. The use of egg yolk derived antibody for passive immunisation of chickens. J. Comp. Pathol. 1969, 79, 495–506. [Google Scholar] [CrossRef]
- Cova, L. DNA-designed avian IgY antibodies: Novel tools for research, diagnostics and therapy. J. Clin. Virol. 2005, 34 (Suppl. 1), S70–S74. [Google Scholar] [CrossRef]
- Salem, R.; Assem, S.K.; Omar, O.A.; Khalil, A.A.; Basry, M.A.; Waly, F.R.; Samir, N.; El-Kholy, A.A. Expressing the immunodominant projection domain of infectious bursal disease virus fused to the fragment crystallizable of chicken IgY in yellow maize for a prospective edible vaccine. Mol. Immunol. 2020, 118, 132–141. [Google Scholar] [CrossRef]
- El-Ghany, W. Comparison between immunoglobulins IgY and the vaccine for prevention of infectious bursal disease in chickens. Glob. Vet. 2011, 6, 16–24. [Google Scholar]
- Jung, K.M.; Bae, E.H.; Jung, Y.T.; Kim, J.W. Use of IgY antibody to recombinant avian reovirus σC protein in the virus diagnostics. Acta Virol. 2014, 58, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C.; Zhang, X.F.; Tan, M.; Huang, P.; Lei, W.; Fang, H.; Zhong, W.; Jiang, X. A dual chicken IgY against rotavirus and norovirus. Antivir. Res. 2013, 97, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Chen, L.; Luo, Y.; Lin, L.; Li, Q.; Peng, P.; Du, Y.; Xu, Z.; Xue, C.; Cao, Y.; et al. Recombinant fiber-2 protein protects Muscovy ducks against duck adenovirus 3 (DAdV-3). Virology 2019, 526, 99–104. [Google Scholar] [CrossRef]
- Luo, Y. Isolation and Identification of Goose Astrovirus and Establishment of Real-Time Quantitative PCR Detection Method. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. [Google Scholar]
- Martín, J.; Crossland, G.; Wood, D.J.; Minor, P.D. Characterization of formaldehyde-inactivated poliovirus preparations m ade from live-attenuated strains. J. Gen. Virol. 2003, 84Pt 7, 1781–1788. [Google Scholar] [CrossRef]
- Sun, L.; Li, M.; Fei, D.; Diao, Q.; Wang, J.; Li, L.; Ma, M. Preparation and Application of Egg Yolk Antibodies Against Chinese Sacbrood Virus Infection. Front. Microbiol. 2018, 9, 1814. [Google Scholar] [CrossRef]
- Lee, C.H.; Liu, C.I.; Leu, S.J.; Lee, Y.C.; Chiang, J.R.; Chiang, L.C.; Mao, Y.C.; Tsai, B.Y.; Hung, C.S.; Chen, C.C.; et al. Chicken antibodies against venom proteins of Trimeresurus stejnegeri in Taiwan. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200056. [Google Scholar] [CrossRef]
- Yang, D.; Mai, K.; Zhou, Q.; Zhu, Y.; Xing, J.; Luo, C.; Liu, S.; Zhou, Q.; Huang, W.; Luo, J.; et al. The protective efficacy of specific egg yolk immunoglobulin Y(IgY) against Riemerella Anatipestifer infections. Vet. Microbiol. 2020, 243, 108642. [Google Scholar] [CrossRef]
- Serena, M.S.; Geisler, C.; Metz, G.E.; Mórtola, E.C.; Echeverría, M.G. Production of pseudorabies virus recombinant glycoprotein B and its use in an agar gel immunodiffusion (AGID) test for detection of antibodies with sensitivity and specificity equal to the virus neutralization assay. J. Virol. Methods 2016, 230, 9–12. [Google Scholar] [CrossRef]
- Smith, P.C.; Stewart, W.C. Agar-gel immunodiffusion assay for pseudorabies virus antibody. J. Clin. Microbiol. 1978, 7, 423–425. [Google Scholar] [CrossRef]
- Akita, E.M.; Nakai, S. Comparison of four purification methods for the production of immunoglobulins from eggs laid by hens immunized with an enterotoxigenic E. coli strain. J. Immunol. Methods 1993, 160, 207–214. [Google Scholar] [CrossRef]
- Polson, A.; von Wechmar, M.B.; van Regenmortel, M.H. Isolation of viral IgY antibodies from yolks of immunized hens. Immunol. Commun. 1980, 9, 475–493. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Y.; Li, Y.; Wang, X.; Liu, Y.; Tian, D.; Jia, X.; Gong, R.; Liu, W.; Yang, L. IgY antibodies against Ebola virus possess post-exposure protection in a murine pseudovirus challenge model and excellent thermostability. PLoS Negl. Trop. Dis. 2021, 15, e0008403. [Google Scholar] [CrossRef]
- Bao, L.; Zhang, C.; Lyu, J.; Yi, P.; Shen, X.; Tang, B.; Zhao, H.; Ren, B.; Kuang, Y.; Zhou, L.; et al. Egg yolk immunoglobulin (IgY) targeting SARS-CoV-2 S1 as potential virus entry blocker. J. Appl. Microbiol. 2022, 132, 2421–2430. [Google Scholar] [CrossRef]
- Nie, J.; Li, Q.; Wu, J.; Zhao, C.; Hao, H.; Liu, H.; Zhang, L.; Nie, L.; Qin, H.; Wang, M.; et al. Quantification of SARS-CoV-2 neutralizing antibody by a pseudotyped virus-based assay. Nat. Protoc. 2020, 15, 3699–3715. [Google Scholar] [CrossRef]
- An, D.; Zhang, J.; Yang, J.; Tang, Y.; Diao, Y. Novel goose-origin astrovirus infection in geese: The effect of age at infection. Poult. Sci. 2020, 99, 4323–4333. [Google Scholar] [CrossRef]
- Wei, F.; Yang, J.; He, D.; Diao, Y.; Tang, Y. Evidence of vertical transmission of novel astrovirus virus in goose. Vet. Microbiol. 2020, 244, 108657. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Yang, J.; Tian, J.; He, D.; Tang, Y.; Diao, Y. Establishment and application of a TaqMan-based one-step real-time RT-PCR for the detection of novel goose-origin astrovirus. J. Virol. Methods 2020, 275, 113757. [Google Scholar] [CrossRef]
- Ji, J.; Chen, Q.; Sui, C.; Hu, W.; Yu, Z.; Zhang, Z.; Mu, X.; Xu, X.; Yao, L.; Kan, Y. Rapid and visual detection of novel astroviruses causing fatal gout in goslings using one-step reverse transcription loop-mediated isothermal amplification. Poult. Sci. 2020, 99, 4259–4264. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, H.; Gao, S.; Song, M.; Shi, Z.; Peng, Z.; Jin, Q.; Zhao, L.; Qiao, H.; Bian, C. Core antigenic advantage domain–based ELISA to detect antibody against novel goose astrovirus in breeding geese. Appl. Microbiol. Biotechnol. 2022, 106, 2053–2062. [Google Scholar] [CrossRef]
- Huang, H.; Ding, R.; Chen, Z.; Yi, Z.; Wang, H.; Lv, Y.; Bao, E. Goose nephritic astrovirus infection increases autophagy, destroys intercellular junctions in renal tubular epithelial cells, and damages podocytes in the kidneys of infected goslings. Vet. Microbiol. 2021, 263, 109244. [Google Scholar] [CrossRef] [PubMed]
- Leiva, C.L.; Gallardo, M.J.; Casanova, N.; Terzolo, H.; Chacana, P. IgY-technology (egg yolk antibodies) in human medicine: A review of patents and clinical trials. Int. Immunopharmacol. 2020, 81, 106269. [Google Scholar] [CrossRef]
- Goudswaard, J. Neonatology: Immunological features. Tijdschr. Voor Diergeneeskd. 1983, 108, 12–21. [Google Scholar]
- Zang, Y.T.; Wang, B.; Song, Y.; Ding, G.J. Research of Passive Immunity Periods of Goose Parvovirus Yolk Antibodies. Anim. Husb. Feed Sci. 2017, 19, 65–66. [Google Scholar]
- Yang, Y.; Ren, P.S.; Cui, H.J.; Zhang, Y.X. Study on Therapeutic Effect of Yolk Antibody Against Duck Viral Hepatitis. Feed Rev. 2021, 8, 16–18. [Google Scholar]
- Schade, R.; Hlinak, A. Egg Yolk Antibodies, State of the Art and Future Prospects. Altex 1996, 13, 5–9. [Google Scholar]
- Kacskovics, I. Fc receptors in livestock species. Vet. Immunol. Immunopathol. 2004, 102, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Lee, Y.C. Site-specific N-glycosylation of chicken serum IgG. Glycobiology 2004, 14, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Calvert, R.A.; Sutton, B.J.; Doré, K.A. IgY: A key isotype in antibody evolution. Biol. Rev. Camb. Philos. Soc. 2017, 92, 2144–2156. [Google Scholar] [CrossRef] [PubMed]
- Hair, P.S.; Gronemus, J.Q.; Crawford, K.B.; Salvi, V.P.; Cunnion, K.M.; Thielens, N.M.; Arlaud, G.J.; Rawal, N.; Krishna, N.K. Human astrovirus coat protein binds C1q and MBL and inhibits the classical and lectin pathways of complement activation. Mol. Immunol. 2010, 47, 792–798. [Google Scholar] [CrossRef]
- Meyerhoff, R.R.; Nighot, P.K.; Ali, R.A.; Blikslager, A.T.; Koci, M.D. Characterization of turkey inducible nitric oxide synthase and identification of its expression in the intestinal epithelium following astrovirus infection. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Qiu, S.; Huang, H.; Xu, R.; Bao, E.; Lv, Y. Immune-related gene expression in the kidneys and spleens of goslings infected with goose nephritic astrovirus. Poult. Sci. 2021, 100, 100990. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mL) | Number of Samples | Number of Infections | Ratio of Positive to Total Samples (a/c) | Ratio of Negative to Total Samples (b/c) |
|---|---|---|---|---|---|
| 1 | 0.3 | 11 | 5 | 5/11 | 6/11 |
| 2 | 0.6 | 11 | 3 | 3/11 | 8/11 |
| 3 | 0.9 | 11 | 1 | 1/11 | 10/11 |
| 4 | 1.2 | 11 | 0 | 0/11 | 11/11 |
| 5 | PBS | 11 | 7 | 7/11 | 4/11 |
| Group | Number of Samples | The Ratio of Positive to Negative Samples at Different Time Points (a/b) | Ratio of Negative to Total Samples (b/c) | ||
|---|---|---|---|---|---|
| 4 d | 8 d | 12 d | |||
| 6 | 10 | 0/10 | 1/9 | 1/9 | 9/10 |
| 13 | 5 | 2/3 | 1/3 | 1/3 | 3/5 |
| 7 | 10 | 0/10 | 1/9 | 1/9 | 9/10 |
| 14 | 5 | 4/1 | 3/0 | 3/0 | 0/5 |
| 8 | 10 | 0/10 | 0/10 | 0/10 | 10/10 |
| 15 | 5 | 3/2 | 2/2 | 2/2 | 2/5 |
| 9 | 10 | 0/10 | 0/10 | 0/10 | 10/10 |
| 16 | 5 | 2/3 | 2/2 | 4/0 | 0/5 |
| 10 | 10 | 1/9 | 2/8 | 2/8 | 8/10 |
| 17 | 5 | 2/3 | 3/2 | 5/0 | 0/5 |
| 11 | 10 | 3/7 | 4/6 | 4/6 | 6/10 |
| 18 | 5 | 2/3 | 3/2 | 3/2 | 2/5 |
| 12 | 10 | 1/6 | 3/4 | 3/4 | 4/10 |
| 19 | 5 | 2/3 | 3/2 | 3/2 | 2/5 |
| Group | Number of Samples | Number of Infections | The Ratio of Positive to Negative Samples at Different Time Points (a/b) | Ratio of Negative to TOTAL Samples (b/c) | ||
|---|---|---|---|---|---|---|
| 4 d | 8 d | 12 d | ||||
| 20 | 5 | 0 | 0/5 | 0/5 | 0/5 | 5/5 |
| 21 | 5 | 1 | 1/4 | 1/4 | 1/4 | 4/5 |
| 22 | 5 | 3 | 1/4 | 3/2 | 3/2 | 2/5 |
| 23 | 5 | 5 | 1/4 | 5/0 | 5/0 | 0/5 |
| 24 | 5 | 5 | 5/0 | 5/0 | 5/0 | 0/5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhang, L.; Yang, J.; Zhao, D.; Han, K.; Huang, X.; Liu, Q.; Xiao, Y.; Gu, Y.; Li, Y. An IgY Effectively Prevents Goslings from Virulent GAstV Infection. Vaccines 2022, 10, 2090. https://doi.org/10.3390/vaccines10122090
Zhang M, Zhang L, Yang J, Zhao D, Han K, Huang X, Liu Q, Xiao Y, Gu Y, Li Y. An IgY Effectively Prevents Goslings from Virulent GAstV Infection. Vaccines. 2022; 10(12):2090. https://doi.org/10.3390/vaccines10122090
Chicago/Turabian StyleZhang, Mengran, Lijiao Zhang, Jing Yang, Dongmin Zhao, Kaikai Han, Xinmei Huang, Qingtao Liu, Yichen Xiao, Youfang Gu, and Yin Li. 2022. "An IgY Effectively Prevents Goslings from Virulent GAstV Infection" Vaccines 10, no. 12: 2090. https://doi.org/10.3390/vaccines10122090
APA StyleZhang, M., Zhang, L., Yang, J., Zhao, D., Han, K., Huang, X., Liu, Q., Xiao, Y., Gu, Y., & Li, Y. (2022). An IgY Effectively Prevents Goslings from Virulent GAstV Infection. Vaccines, 10(12), 2090. https://doi.org/10.3390/vaccines10122090