
Simultaneous Protective Immune Responses of Ducks against Duck Plague and Fowl Cholera by Recombinant Duck Enteritis Virus Vector Expressing Pasteurella multocida OmpH Gene

 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Immunization and Challenge Exposure
2.3. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
2.4. ELISA Test to Evaluate Immune Response to P. multocida Strain X-73
2.5. ELISA to Evaluate Immune Response to DEV
2.6. Determination of Cellular Response by MTT Proliferation Assays
2.7. Serum Detection Level of IFN-γ and IL-4 Cytokine Assay
2.8. Detection of CD4+ and CD8+ T Lymphocytes in Peripheral Blood by Flow Cytometry
2.9. Statistical Analysis
3. Results
3.1. Virulence and Sustainability of Recombinant DEV-ompH Virus in Tissues
3.2. The Serological Responses against DEV and P. multocida Strain X-73 Measured by ELISA
3.3. Vaccine Efficacy against Lethal DEV and P. multocida Strain X-73 Challenge in Ducks
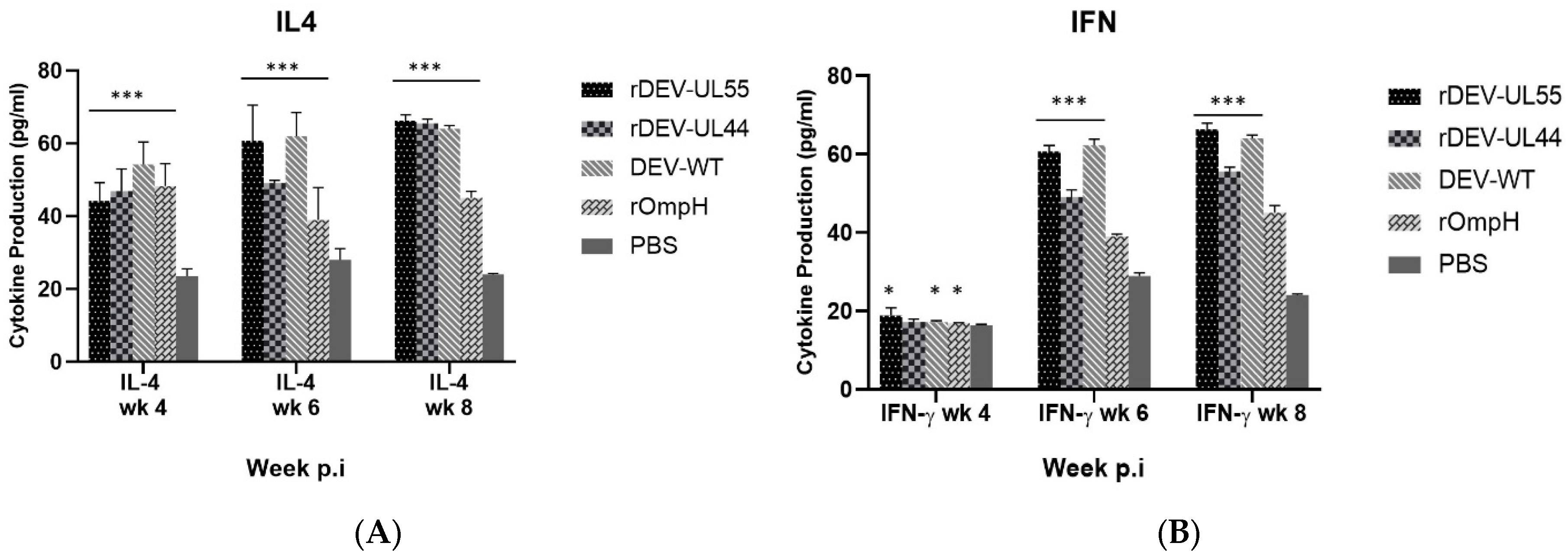
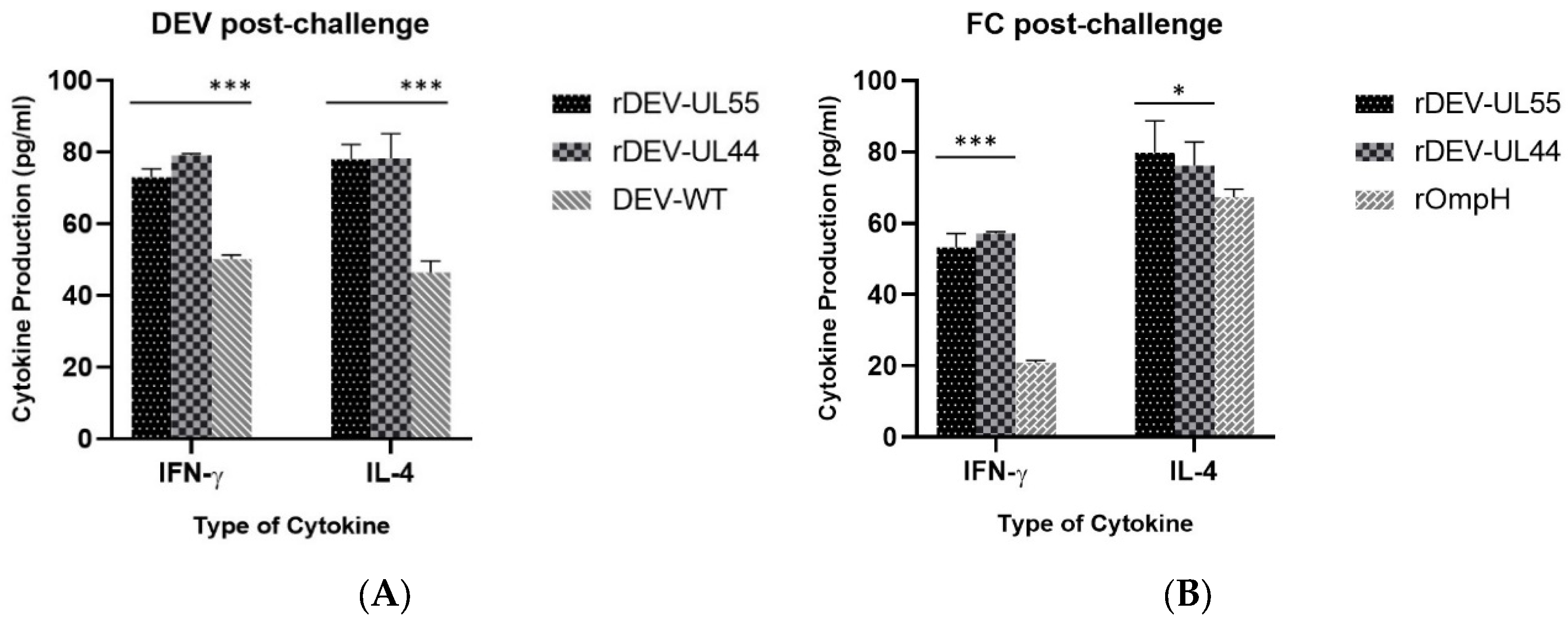
3.4. Detection of IFN-γ and IL-4 in Serum as the Cellular Response to Recombinant DEV-ompH-UL55 and DEV-ompH-UL44
3.5. The Cellular Response to Recombinant DEV-ompH-UL55 and DEV-ompH-UL44 by In Vitro Lymphocyte Proliferation Assay (LPA)
3.6. Analysis of CD4+ and CD8+ T Lymphocytes in Peripheral Blood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaves Hernández, A.J. Poultry and Avian Diseases. In Encyclopedia of Agriculture and Food Systems; Van Alfen, N.K., Ed.; Academic Press: Cambridge, UK, 2014; pp. 504–520. [Google Scholar]
- Samuel, M.D.; Botzler, R.G.; Wobeser, G.A. Avian cholera. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Atkinson, C.T., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 239–269. [Google Scholar]
- Sthitmatee, N.; Numee, S.; Kawamoto, E.; Sasaki, H.; Yamashita, K.; Takahashi, N.; Kataoka, Y.; Sawada, T. Protection of chickens from fowl cholera by vaccination with recombinant adhesive protein of Pasteurella multocida. Vaccine 2008, 26, 2398–2407. [Google Scholar] [CrossRef]
- Thanasarasakulpong, A.; Poolperm, P.; Tangjitjaroen, W.; Varinrak, T.; Sawada, T.; Pfeiffer, D.; Sthitmatee, N. Comparison of the effect of two purification methods on the immunogenicity of recombinant outer membrane protein H of Pasteurella multocida serovar A:1. Vet. Med. Int. 2016, 2016, 2579345. [Google Scholar] [CrossRef]
- Thanasarasakulpong, A.; Poolperm, P.; Tankaew, P.; Sawada, T.; Sthitmatee, N. Protectivity conferred by immunization with intranasal recombinant outer membrane protein H from Pasteurella multocida serovar A:1 in chickens. J. Vet. Med. Sci. 2015, 77, 321–326. [Google Scholar] [CrossRef]
- Varinrak, T.; Muenthaisong, A.; Apinda, N.; Sawada, T.; Sthitmatee, N. Construction and characterization of an OmpH-deficient mutant of Pasteurella multocida strain X-73. Avian Pathol. 2019, 48, 4–11. [Google Scholar] [CrossRef]
- Varinrak, T.; Poolperm, P.; Sawada, T.; Sthitmatee, N. Cross-protection conferred by immunization with an rOmpH-based intranasal fowl cholera vaccine. Avian Pathol. 2017, 46, 515–525. [Google Scholar] [CrossRef]
- Apinda, N.; Nambooppha, B.; Rittipornlertrak, A.; Tankaew, P.; Punyapornwithaya, V.; Nair, V.; Sawada, T.; Sthitmatee, N. Protection against fowl cholera in ducks immunized with a combination vaccine containing live attenuated duck enteritis virus and recombinant outer membrane protein H of Pasteurella multocida. Avian Pathol. 2020, 49, 221–229. [Google Scholar] [CrossRef]
- Poolperm, P.; Apinda, N.; Kataoka, Y.; Suriyasathaporn, W.; Tragoolpua, K.; Sawada, T.; Sthitmatee, N. Protection against Pasteurella multocida conferred by an intranasal fowl cholera vaccine in Khaki Campbell ducks. Jpn. J. Vet. Res. 2018, 66, 239–250. [Google Scholar]
- Belloc, C.; Dupuis, L.; Deville, S.; Aucouturier, J.; Laval, A. Evaluation of safety and immune response induced by several adjuvants included in Pasteurella multocida vaccines in chickens. Rev. Médecine Vétérinaire 2008, 159, 371–375. [Google Scholar]
- Sandhu, T.S.; Shawky, S. Duck virus enteritis (duck plague). In Diseases of Poultry; Saif, Y.M., Fadly, A.M., Glisson, J.R., McDougald, L.R., Nolan, L.K., Swayne, D.E., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2008; Volume 11, pp. 354–363. [Google Scholar]
- Sandhu, T.S.; Shawky, S. Duck virus enteritis (duck plague). In Diseases of Poultry; Saif, Y.M., Fadly, A.M., Glisson, J.R., McDougald, L.R., Nolan, L.K., Swayne, D.E., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2003; Volume 10, pp. 675–683. [Google Scholar]
- Jansen, J.; Kunst, H.; Wemmenhove, R. The active immunization of ducks against duck plague. Tijdschr Diergeneesk 1963, 88, 927–932. [Google Scholar]
- Olobatoke, R.; Mulugeta, S. Effect of dietary garlic powder on layer performance, fecal bacterial load, and egg quality. Poult. Sci. 2011, 90, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Mazurowska-Magdzik, W. Monovalent and combined vaccines. Pros and cons. Prz. Pediatryczny 2001, 31, 103–106. [Google Scholar]
- Teng, M.; Yao, Y.; Nair, V.; Luo, J. Latest advances of virology research using CRISPR/Cas9-based gene-editing technology and its application to vaccine development. Viruses 2021, 13, 779. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, Y.; Shen, Z.; Yao, Y.; Nair, V. Application of CRISPR-Cas9 editing for virus engineering and the development of recombinant viral vaccines. CRISPR J. 2021, 4, 477–490. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, X.; Shen, B.; Lu, Y.; Chen, W.; Ma, J.; Bai, L.; Huang, X.; Zhang, L. Generating rats with conditional alleles using CRISPR/Cas9. Cell Res. 2014, 24, 122–125. [Google Scholar] [CrossRef]
- Vilela, J.; Rohaim, M.A.; Munir, M. Application of CRISPR/Cas9 in understanding avian viruses and developing poultry vaccines. Front. Cell. Infect. Microbiol. 2020, 10, 581504. [Google Scholar] [CrossRef]
- Chen, L.; Yu, B.; Hua, J.; Ye, W.; Ni, Z.; Yun, T.; Deng, X.; Zhang, C. Construction of a full-length infectious bacterial artificial chromosome clone of duck enteritis virus vaccine strain. Virol. J. 2013, 10, 328. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Han, Z.; Wang, Y.; Liang, S.; Jiang, L.; Hu, Y.; Kong, X.; Liu, S. Recombinant duck enteritis viruses expressing major structural proteins of the infectious bronchitis virus provide protection against infectious bronchitis in chickens. Antivir. Res. 2016, 130, 19–26. [Google Scholar] [CrossRef]
- Liu, J.; Chen, P.; Jiang, Y.; Wu, L.; Zeng, X.; Tian, G.; Ge, J.; Kawaoka, Y.; Bu, Z.; Chen, H. A duck enteritis virus-vectored bivalent live vaccine provides fast and complete protection against H5N1 avian influenza virus infection in ducks. J. Virol. 2011, 85, 10989–10998. [Google Scholar] [CrossRef]
- Liu, X.; Wei, S.; Liu, Y.; Fu, P.; Gao, M.; Mu, X.; Liu, H.; Xing, M.; Ma, B.; Wang, J. Recombinant duck enteritis virus expressing the HA gene from goose H5 subtype avian influenza virus. Vaccine 2013, 31, 5953–5959. [Google Scholar] [CrossRef]
- Wang, J.; Osterrieder, N. Generation of an infectious clone of duck enteritis virus (DEV) and of a vectored DEV expressing hemagglutinin of H5N1 avian influenza virus. Virus Res. 2011, 159, 23–31. [Google Scholar] [CrossRef]
- Zou, Z.; Hu, Y.; Liu, Z.; Zhong, W.; Cao, H.; Chen, H.; Jin, M. Efficient strategy for constructing duck enteritis virus-based live attenuated vaccine against homologous and heterologous H5N1 avian influenza virus and duck enteritis virus infection. Vet. Res. 2015, 46, 42. [Google Scholar] [CrossRef]
- Zou, Z.; Huang, K.; Wei, Y.; Chen, H.; Liu, Z.; Jin, M. Construction of a highly efficient CRISPR/Cas9-mediated duck enteritis virus-based vaccine against H5N1 avian influenza virus and duck Tembusu virus infection. Sci. Rep. 2017, 7, 1478. [Google Scholar] [CrossRef]
- Apinda, N.; Yao, Y.; Zhang, Y.; Reddy, V.R.A.P.; Chang, P.; Nair, V.; Sthitmatee, N. CRISPR/Cas9 editing of duck enteritis virus genome for the construction of a recombinant vaccine vector expressing ompH gene of Pasteurella multocida in two novel insertion sites. Vaccines 2022, 10, 686. [Google Scholar] [CrossRef]
- Klimpel, G.R. Immune defenses. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston Press: Galveston, TX, USA, 1996. Available online: https://www.ncbi.nlm.nih.gov/books/NBK7627/ (accessed on 16 March 2022).
- Poolperm, P.; Varinrak, T.; Kataoka, Y.; Tragoolpua, K.; Sawada, T.; Sthitmatee, N. Development and standardization of an in-house indirect ELISA for detection of duck antibody to fowl cholera. J. Microbiol. Methods 2017, 142, 10–14. [Google Scholar] [CrossRef]
- Underwood, W.; Anthony, R. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; Retrieved on March 2020; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- Li, L.; Cheng, A.; Wang, M.; Xiang, J.; Yang, X.; Zhang, S.; Zhu, D.; Jia, R.; Luo, Q.; Zhou, Y. Expression and characterization of duck enteritis virus gI gene. Virol. J. 2011, 8, 241. [Google Scholar] [CrossRef]
- Huang, J.; Jia, R.; Wang, M.; Shu, B.; Yu, X.; Zhu, D.; Chen, S.; Yin, Z.; Chen, X.; Cheng, A. An attenuated duck plague virus (DPV) vaccine induces both systemic and mucosal immune responses to protect ducks against virulent DPV infection. Clin. Vaccine Immunol. 2014, 21, 457–462. [Google Scholar] [CrossRef]
- Hein, R.; Koopman, R.; García, M.; Armour, N.; Dunn, J.R.; Barbosa, T.; Martinez, A. Review of Poultry Recombinant Vector Vaccines. Avian Dis. 2021, 65, 438–452. [Google Scholar] [CrossRef]
- Sternberg, N.; Hamilton, D. Bacteriophage P1 site-specific recombination: I. Recombination between loxP sites. J. Mol. Biol. 1981, 150, 467–486. [Google Scholar] [CrossRef]
- Baron, M.D.; Iqbal, M.; Nair, V. Recent advances in viral vectors in veterinary vaccinology. Curr. Opin. Virol. 2018, 29, 1–7. [Google Scholar] [CrossRef]
- Francis, M.J. Spotlight on avian pathology: The importance of recombinant vector platform technologies in poultry vaccination. Avian Pathol. 2021, 50, 109–111. [Google Scholar] [CrossRef]
- de Vries, R.D.; Rimmelzwaan, G.F. Viral vector-based influenza vaccines. Hum. Vaccines Immunother. 2016, 12, 2881–2901. [Google Scholar] [CrossRef]
- Qi, X.-F.; Cheng, A.-C.; Wang, M.-S.; Yang, X.-Y.; Jia, R.-Y.; Chen, X.-Y. Development of an indirect-ELISA kit for detection of antibodies against duck plague virus. Vet. Sci. China 2007, 37, 690–694. [Google Scholar]
- Cai, M.-S.; Deng, S.-X.; Li, M.-L. Comparison of the immune responses in BALB/c mice following immunization with DNA-based and live attenuated vaccines delivered via different routes. Vaccine 2013, 31, 1353–1356. [Google Scholar] [CrossRef]
- Lian, B.; Cheng, A.; Wang, M.; Zhu, D.; Luo, Q.; Jia, R.; Liu, F.; Han, X.; Chen, X. Induction of immune responses in ducks with a DNA vaccine encoding duck plague virus glycoprotein C. Virol. J. 2011, 8, 214. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, A.; Wang, M.; Zhang, S.; Zhu, D.; Jia, R.; Luo, Q.; Chen, Z.; Chen, X. Serologic detection of duck enteritis virus using an indirect ELISA based on recombinant UL55 protein. Avian Dis. 2011, 55, 626–632. [Google Scholar] [CrossRef]
- Yu, X.; Jia, R.; Huang, J.; Shu, B.; Zhu, D.; Liu, Q.; Gao, X.; Lin, M.; Yin, Z.; Wang, M. Attenuated Salmonella typhimurium delivering DNA vaccine encoding duck enteritis virus UL24 induced systemic and mucosal immune responses and conferred good protection against challenge. Vet. Res. 2012, 43, 56. [Google Scholar] [CrossRef]
- Liu, J.; Chen, P.; Jiang, Y.; Deng, G.; Shi, J.; Wu, L.; Lin, Y.; Bu, Z.; Chen, H. Recombinant duck enteritis virus works as a single-dose vaccine in broilers providing rapid protection against H5N1 influenza infection. Antivir. Res. 2013, 97, 329–333. [Google Scholar] [CrossRef]
- Wang, J.; Ge, A.; Xu, M.; Wang, Z.; Qiao, Y.; Gu, Y.; Liu, C.; Liu, Y.; Hou, J. Construction of a recombinant duck enteritis virus (DEV) expressing hemagglutinin of H5N1 avian influenza virus based on an infectious clone of DEV vaccine strain and evaluation of its efficacy in ducks and chickens. Virol. J. 2015, 12, 126. [Google Scholar] [CrossRef]
- Tang, J.; Yin, D.; Wang, R.; Zhou, Q.; Zhou, X.; Xing, X.; Liu, H.-M.; Liu, G.; Wang, G. A recombinant adenovirus expressing the E protein of duck Tembusu virus induces protective immunity in duck. J. Vet. Med. Sci. 2018, 81, 314–320. [Google Scholar] [CrossRef]
- Petersone, L.; Edner, N.M.; Ovcinnikovs, V.; Heuts, F.; Ross, E.M.; Ntavli, E.; Wang, C.J.; Walker, L.S. T cell/B cell collaboration and autoimmunity: An intimate relationship. Front. Immunol. 2018, 9, 1941. [Google Scholar] [CrossRef]
- Jennings, S.R.; Bonneau, R.H.; Smith, P.M.; Wolcott, R.M.; Chervenak, R. CD4-positive T lymphocytes are required for the generation of the primary but not the secondary CD8-positive cytolytic T lymphocyte response to herpes simplex virus in C57BL/6 mice. Cell. Immunol. 1991, 133, 234–252. [Google Scholar] [CrossRef]
- Baumjohann, D.; Preite, S.; Reboldi, A.; Ronchi, F.; Ansel, K.M.; Lanzavecchia, A.; Sallusto, F. Persistent antigen and germinal center B cells sustain T follicular helper cell responses and phenotype. Immunity 2013, 38, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Phares, T.W.; Stohlman, S.A.; Hwang, M.; Min, B.; Hinton, D.R.; Bergmann, C.C. CD4 T cells promote CD8 T cell immunity at the priming and effector site during viral encephalitis. J. Virol. 2012, 86, 2416–2427. [Google Scholar] [CrossRef]
- Sabarth, N.; Chamberlain, L.; Brett, S.; Tite, J.; Craigen, J. Induction of homologous rather than heterologous antigen-specific CD4 T cell responses is critical for functional CD8 T cell responses in mice transgenic for a foreign antigen. J. Immunol. 2010, 185, 4590–4601. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Ni, G.; Damania, B. Innate sensing of DNA virus genomes. Annu. Rev. Virol. 2018, 5, 341–362. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vaccination Formulation | Challenge Exposure (IM) Duck/Group | |
|---|---|---|---|
| Virulent Local Type DEV (100-fold DLD50) | P. multocida X-73 (3.5 × 103 CFU/mL) | ||
| 1 | rDEV-Omp-UL55 10 5 PFU a | 10 | 10 |
| 2 | rDEV-Omp-UL44 10 5 PFU a | 10 | 10 |
| 3 | DEV-WT 10 3.5 PFU b | 10 | 10 |
| 4 | rOmpH 100 µg/mL | 10 | 10 |
| 5 | PBS | 10 | 10 |
| Total | 100 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apinda, N.; Muenthaisong, A.; Chomjit, P.; Sangkakam, K.; Nambooppha, B.; Rittipornlertrak, A.; Koonyosying, P.; Yao, Y.; Nair, V.; Sthitmatee, N. Simultaneous Protective Immune Responses of Ducks against Duck Plague and Fowl Cholera by Recombinant Duck Enteritis Virus Vector Expressing Pasteurella multocida OmpH Gene. Vaccines 2022, 10, 1358. https://doi.org/10.3390/vaccines10081358
Apinda N, Muenthaisong A, Chomjit P, Sangkakam K, Nambooppha B, Rittipornlertrak A, Koonyosying P, Yao Y, Nair V, Sthitmatee N. Simultaneous Protective Immune Responses of Ducks against Duck Plague and Fowl Cholera by Recombinant Duck Enteritis Virus Vector Expressing Pasteurella multocida OmpH Gene. Vaccines. 2022; 10(8):1358. https://doi.org/10.3390/vaccines10081358
Chicago/Turabian StyleApinda, Nisachon, Anucha Muenthaisong, Paweena Chomjit, Kanokwan Sangkakam, Boondarika Nambooppha, Amarin Rittipornlertrak, Pongpisid Koonyosying, Yongxiu Yao, Venugopal Nair, and Nattawooti Sthitmatee. 2022. "Simultaneous Protective Immune Responses of Ducks against Duck Plague and Fowl Cholera by Recombinant Duck Enteritis Virus Vector Expressing Pasteurella multocida OmpH Gene" Vaccines 10, no. 8: 1358. https://doi.org/10.3390/vaccines10081358
APA StyleApinda, N., Muenthaisong, A., Chomjit, P., Sangkakam, K., Nambooppha, B., Rittipornlertrak, A., Koonyosying, P., Yao, Y., Nair, V., & Sthitmatee, N. (2022). Simultaneous Protective Immune Responses of Ducks against Duck Plague and Fowl Cholera by Recombinant Duck Enteritis Virus Vector Expressing Pasteurella multocida OmpH Gene. Vaccines, 10(8), 1358. https://doi.org/10.3390/vaccines10081358