Abstract
Age-related changes in the immune system are thought to underlie the vulnerability of elderly individuals to emerging viral diseases, such as coronavirus disease 2019 (COVID-19). In this study, we used a fully validated in vitro approach to determine how age impacts the generation of de novo CD8+ T cell responses against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), the causative agent of COVID-19. Our data revealed a generalized deficit in the ability of elderly individuals to prime the differentiation of naïve precursors into effector CD8+ T cells defined by the expression of interferon (IFN)-γ and the transcription factor T-bet. As a consequence, there was an age-related decline in the diversity of newly generated CD8+ T cell responses targeting a range of typically immunodominant epitopes derived from SARS-CoV-2, accompanied by an overall reduction in the expression frequency of IFN-γ. These findings have potential implications for the development of new strategies to protect the elderly against COVID-19.
1. Introduction
In December 2019, a novel coronavirus, severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), was discovered as the causative agent of an outbreak of lower respiratory tract infections in Wuhan, China. As the virus spread globally, it became clear that elderly individuals responded suboptimally to vaccination and were particularly susceptible to severe disease, with high attendant rates of mortality [1,2,3,4]. These observations suggested an age-related link between antiviral immunity and disease outcome, but a precise understanding of the determinative biological processes has been lacking to date.
Physiological ageing has been linked with a progressive decline in immune functionality, known as immunosenescence, which could account for the increased vulnerability of elderly individuals to emerging infectious agents, such as SARS-CoV-2 [5,6]. In line with this notion, a relative scarcity of naïve T cells has been associated with severe coronavirus disease 2019 (COVID-19) [7]. Naïve T cells are known to become less frequent and more dysfunctional with age [8,9,10]. It is therefore plausible that functional and numerical alterations in the naïve pool could impair the induction of de novo SARS-CoV-2-specific effector/memory T cells in the elderly, consistent with previously described age-related immune deficits in the context of acute COVID-19 [11].
To address this potential mechanistic link, we used an in vitro priming approach to quantify the induction of epitope-specific effector/memory CD8+ T cells from naïve precursors as a function of age. Importantly, all donors were unvaccinated and previously unexposed to SARS-CoV-2, enabling a robust assessment of priming efficacy in the absence of immunological memory. Our data revealed that advanced age was associated with an impaired ability to mount de novo CD8+ T cell responses against a range of immunodominant antigens derived from SARS-CoV-2.
2. Materials and Methods
2.1. Donors
Peripheral blood samples were obtained from anonymized donors (age, 20–69 years; female, n = 2; male, n = 14) via the Blood Bank at Azienda Ospedaliera-Universitaria di Ferrara, Italy. The protocol was approved by the Azienda Unità Sanitaria Locale di Ferrara. All donors expressed human leukocyte antigen (HLA)-A2. Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats via density gradient centrifugation using Ficoll-Paque (GE Healthcare, Milan, Italy) and cryopreserved in fetal bovine serum (FBS; Euroclone, Milan, Italy) containing 10% dimethyl sulfoxide (DMSO; Sigma-Aldrich, Milan, Italy). Samples were obtained either before or during 2020. Donors in the latter group were only included after testing seronegative for SARS-CoV-2. Serostatus was assessed using a SARS-CoV-2 RBD Total Antibody ELISA Kit (Mabtech, Stockholm, Sweden).
2.2. Peptides and Tetramers
HLA-A2-restricted peptides were synthesized in solid phase and purified via high-performance liquid chromatography (>97%). The melanoma-derived EV10 peptide (ELAGIGILTV) was used as a model antigen, given constitutively high cognate precursor frequencies among individuals expressing HLA-A2 [12]. SARS-CoV-2 peptides were selected from different viral proteins on the basis of immunodominance/immunoprevalence [13] and matched to the wildtype strain HKU-001a (Table 1).

Table 1.
SARS-CoV-2 peptides and tetramers restricted by HLA-A2.
Peptides were dissolved initially in DMSO and used at a final concentration of 1 μM. Fluorescent peptide/HLA-A2 tetramers were generated as described previously [14]. All tetramers were used at an identical titer immediately after conjugation to ExtrAvidin-PE (Sigma-Aldrich).
2.3. In Vitro Priming of Human Antigen-Specific CD8+ T Cells
PBMCs were thawed and resuspended in 75-cm2 tissue culture flasks (7 × 107 cells/flask to prime SARS-CoV-2-specific naïve T cells) or 48-well tissue culture plates (2.5 × 105 cells/well to prime EV10-specific naïve T cells) containing AIM-V medium (Thermo Fisher Scientific, Monza, Italy) supplemented with FLT3L (50 ng/mL; Miltenyi Biotec, Bologna, Italy) to mobilize resident dendritic cells (DCs) [13]. After 24 h (day 1), the corresponding peptides were added to the cultures at a final concentration of 1 μM, and DC maturation was induced with TNF-α (1000 U/mL; Miltenyi Biotec), IL-1β (10 ng/mL; Miltenyi Biotec), IL-7 (0.5 ng/mL; R&D Systems, Minneapolis, MN, USA), and prostaglandin E2 (1 μM; Calbiochem, Milan, Italy). On day 2, complement-inactivated FBS (Euroclone) was added at a final v/v ratio of 10%. Medium was replaced on days 4 and 7 with fresh RPMI 1640 (Euroclone) enriched with 10% FBS (Euroclone), non-essential amino acids (1X; Euroclone), and sodium pyruvate (1 mM; Sigma-Aldrich). Antigen-specific CD8+ T cells were characterized on day 10.
2.4. ELISpot Assay
Functional responses were quantified using a human interferon (IFN)-γ ELISpot PLUS (HRP) Kit (Mabtech). PBMCs were primed as described above and seeded in duplicate at 1 × 105 cells/well in 100 μL of fresh RPMI 1640 (Euroclone) enriched with 10% FBS (Euroclone), non-essential amino acids (1X; Euroclone), and sodium pyruvate (1 mM; Sigma-Aldrich). Experimental wells contained peptides at a final concentration of 1 μM. Negative control wells lacked peptide, and positive control wells contained anti-CD3 (clone CD3-2; Mabtech). Plates were incubated for 24 h and developed according to the manufacturer’s instructions (Mabtech). IFN-γ-secreting cells were quantified as spot-forming units (SFUs) per 106 cells using an automated ELISpot Reader (AELVIS, Hannover, Germany). Positive responses were assigned after background subtraction at 50 SFUs per 106 cells (mean). Data were excluded in the absence of a response to anti-CD3.
2.5. Flow Cytometry
Ex vivo analyses were performed using thawed PBMCs. Cells were stained first with LIVE/DEAD Fixable Aqua (Thermo Fisher Scientific) for 10 min at room temperature and then with the following directly conjugated monoclonal antibodies for 15 min at room temperature: anti-CCR7–PE-Cy7 (clone 3D12; BD Biosciences, Milan, Italy), anti-CD8–APC-Cy7 (clone SK1; BD Biosciences), anti-CD27–FITC (clone M-T271; Miltenyi Biotec), anti-CD45RA–PerCP-Cy5.5 (clone HI100; Thermo Fisher Scientific), anti-HLA-DR–APC (clone LN3; Thermo Fisher Scientific), and anti-PD1–PE (clone REA1165; Miltenyi Biotec). Naïve T cells were defined as CCR7+CD27+CD45RA+.
Tetramer staining was performed using 2–3 × 106 primed cells on day 10. Cells were stained serially with LIVE/DEAD Fixable Aqua (Thermo Fisher Scientific) for 10 min at room temperature, individual or pooled tetramers for 15 min at 37 °C, and the following directly conjugated monoclonal antibodies for 15 min at room temperature: anti-CD3–PerCP-Cy5.5 (clone SK7; Thermo Fisher Scientific), anti-CD4–FITC (clone RPA-T4; BD Biosciences), anti-CD8–APC/Fire 750 (clone RPA-T8; BioLegend, Amsterdam, Netherlands), anti-CD14–FITC (clone 18D11; ImmunoTools, Friesoythe, Germany), and anti-CD19–FITC (clone HI19a; ImmunoTools). After fixation/permeabilization using a Transcription Factor Buffer Set (BD Biosciences), cells were stained with anti-Tbet–eFluor 660 (clone 4B10; Thermo Fisher Scientific) for 40 min at 4 °C. Tetramer-labeled CD3+CD8+ T cells were identified among viable cells after exclusion of FITC+ events (dump channel). A positivity threshold was calculated after performing identical experiments in donors lacking expression of HLA-A2.
Data were acquired using a FACSCanto II (BD Biosciences), compensated using CompBeads (BD Biosciences), and analyzed using FlowJo software version 10.8 (FlowJo LLC, Ashland, OR, USA).
2.6. HLA–Peptide Binding Assay
Aliquots of 1 × 106 T2 cells, a lymphoma-derived line expressing low amounts of HLA-A2, were incubated overnight at 26 °C in 1 mL of AIM-V medium containing individual peptides at concentrations ranging from 10−5 M to 10−8 M. Cells were then incubated for a further 4 h at 37 °C, washed extensively in phosphate-buffered saline, and stained with anti-HLA-A2–PE (clone BB7.2; BioLegend). Mean fluorescence intensity was determined via flow cytometry and used to calculate peptide-induced stabilization as the percent increase over baseline expression of HLA-A2 [15,16].
2.7. Statistics
Statistical significance was assessed using the Mann–Whitney U test or the Pearson correlation coefficient in Prism software version 8 (GraphPad, San Diego, CA, USA).
3. Results
3.1. Cohort Description
PBMCs were collected from anonymized adult donors before (2018–2019) or during 2020 (Figure 1a). Donors in the latter group were only included after testing seronegative for SARS-CoV-2. Samples were stratified according to age as “young” (range, 20–52 years; median, 25 years; female, n = 2; male, n = 6) or “old” (range, 65–69 years; median, 68 years; female, n = 0; male, n = 8).
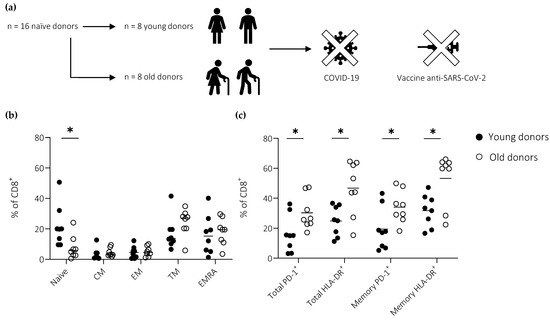
Figure 1.
Activation and differentiation in the peripheral CD8+ T cell compartment as a function of age. (a) Donors were stratified according to age. (b) Frequencies of naïve (CCR7+CD27+CD45RA+), central memory (CM; CCR7+CD27+CD45RA−), transitional memory (TM; CCR7−CD27+CD45RA−), effector memory (EM; CCR7−CD27−CD45RA−), and terminally differentiated EM (EMRA; CCR7−CD27−CD45RA+) CD8+ T cells. (c) Expression frequencies of HLA-DR and PD-1 among total (including naïve) and memory (excluding naïve) CD8+ T cells. (b,c) Horizontal lines represent median values. * p < 0.05 (Mann–Whitney U test).
Basal levels of activation and differentiation in the peripheral CD8+ T cell compartment were characterized using flow cytometry. A proportionate reduction in the size of the naïve pool was observed with age (Figure 1b). Moreover, higher fractions of total (including naïve) and memory (excluding naïve) CD8+ T cells displayed an activated phenotype in old versus young individuals, as defined by the expression frequencies of HLA-DR and PD-1 (Figure 1c).
3.2. Peptide Characterization
HLA-A2-restricted peptide epitopes derived from different viral proteins were used to measure primary CD8+ T cell responses against SARS-CoV-2 (Table 1). The affinity of each peptide for this restriction element was determined using a surface stabilization assay. All peptides bound strongly to HLA-A2. In most cases, stabilization was detected at low peptide concentrations (10−7 M and 10−8 M), although some differences were apparent, especially at high peptide concentrations (10−5 M and 10−6 M), consistent with a binding hierarchy (Figure 2). The lowest relative affinities were observed for the spike-derived peptides YLQ and TLD.
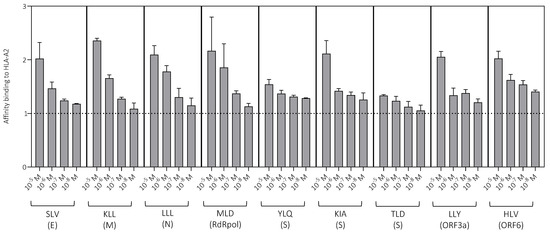
Figure 2.
Peptide binding to HLA-A2. Peptide binding affinity was measured across a range of concentrations using a surface stabilization assay. Data are shown as median + SEM. The dotted line indicates no binding (1). E, envelope; M, membrane; ORF, open reading frame; RdRpol, RNA-dependent RNA polymerase; S, spike.
3.3. Ageing Impairs the Induction of Effector CD8+ T Cells Defined by the Expression of T-bet
PBMCs were stimulated in vitro with the model peptide EV10 and a panel of peptides derived from SARS-CoV-2 (Table 1), all restricted by HLA-A2 (Figure 3a). Epitope-specific CD8+ T cells were quantified via tetramer staining on day 10. This experimental approach to the measurement of priming efficacy has been fully validated in humans and mice [10,17].
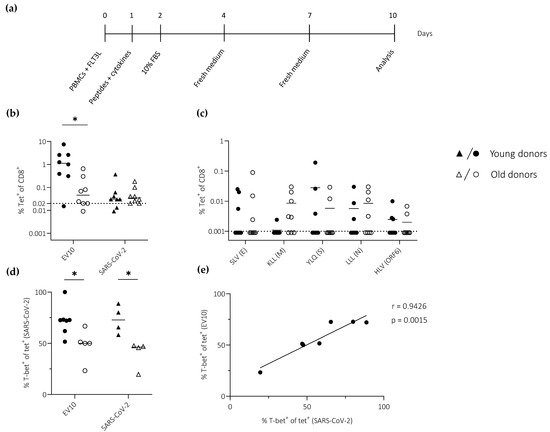
Figure 3.
T-bet expression among primed CD8+ T cells specific for EV10 or SARS-CoV-2. (a) Schematic representation of the in vitro priming approach. (b,c) Epitope-specific CD8+ T cell frequencies quantified using the EV10 tetramer or SARS-CoV-2 tetramers in a single pool (n = 5) (b) or individually (c). Dotted lines indicate the limit of detection. (d) T-bet expression frequencies among tetramer-labeled CD8+ T cells quantified as in (b). (e) Pearson correlation showing T-bet expression frequencies among EV10-specific CD8+ T cells versus T-bet expression frequencies among SARS-CoV-2-specific CD8+ T cells. (b–d) Horizontal lines represent median values. * p < 0.05 (Mann–Whitney U test).
EV10-specific CD8+ T cells from young donors expanded to a substantially greater extent than EV10-specific CD8+ T cells from old donors, resulting in a median frequency difference that exceeded one order of magnitude (Figure 3b). In contrast, only very low frequencies of SARS-CoV-2-specific CD8+ T cells (often <0.1%) were detected using a pool of tetramers (n = 5), irrespective of age (Figure 3b). Similar results were obtained using each tetramer individually (Figure 3c).
To extend these findings, we measured the expression of T-bet, a transcription factor that plays a key role in effector differentiation, among primed CD8+ T cells specific for EV10 or SARS-CoV-2. T-bet was expressed at lower frequencies among primed CD8+ T cells from old versus young donors, irrespective of specificity (Figure 3d). Moreover, a direct correlation was observed between T-bet expression frequencies among EV10-specific CD8+ T cells and T-bet expression frequencies among SARS-CoV-2-specific CD8+ T cells (Figure 3e).
Collectively, these data suggested that age rather than specificity was a key determinant of effector CD8+ T cell differentiation, as defined by the expression of T-bet.
3.4. Ageing Impairs the Induction of Effector CD8+ T Cells Defined by the Expression of IFN-γ
To assess the functional correlates of differential T-bet expression patterns among epitope-specific CD8+ T cells primed via our in vitro approach, we used ELISpot assays to quantify the secretion of IFN-γ.
IFN-γ response frequencies were consistently higher among young versus old donors after stimulation with peptides derived from SARS-CoV-2 (n = 9) (Figure 4a). In the latter group, no responses were observed against the spike-derived peptides KIA and TLD, and responses against the non-spike-derived peptides SLV, MLD, LLY, and HLV were limited to a single donor in each case (Figure 4b). Accordingly, there was an age-related difference in the frequency of epitope recognition, albeit just below the threshold required for significance (Figure 4c). Of note, T-bet expression frequencies among SARS-CoV-2-specific CD8+ T cells correlated directly with epitope recognition frequencies (immunoprevalence), measured as a binary outcome for each peptide (Figure 4d), and overall response frequencies (immunodominance), measured as cumulative SFUs (Figure 4e). No correlation was detected between epitope recognition frequencies and peptide binding affinities for HLA-A2 (Figure S1).
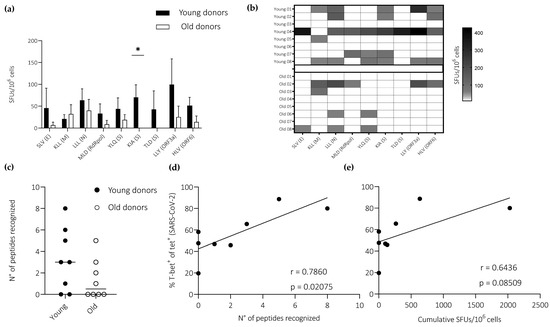
Figure 4.
IFN-γ expression among primed CD8+ T cells specific for SARS-CoV-2. (a) Epitope-specific response frequencies quantified using ELISpot assays to capture IFN-γ after stimulation with peptides derived from SARS-CoV-2. Responses above 50 SFUs per 106 cells were considered positive. Data are shown as median + SEM. (b) Data from (a) shown as a heatmap. (c) The total number of peptides recognized by each donor. Horizontal lines represent median values. (d,e) Pearson correlations showing T-bet expression frequencies among SARS-CoV-2-specific CD8+ T cells versus epitope recognition frequencies, measured as a binary outcome for each peptide (d), and overall response frequencies, measured as cumulative SFUs (e). Dots correspond with the data shown in Figure 3d (young, n = 4; old, n = 4). (a) * p < 0.05 (Mann–Whitney U test).
Collectively, these data suggested that age rather than specificity was a key determinant of effector CD8+ T cell differentiation, as defined by the induction of IFN-γ.
4. Discussion
Ageing is known to alter the composition and functionality of the immune system [8,10,18]. These changes limit the ability of elderly individuals to mount de novo cellular immune responses against previously unencountered pathogens in the context of natural infection and/or vaccination [9,19]. T cells are particularly susceptible to the ravages of age, potentially explaining the vulnerability of elderly individuals to emerging viral infections, such as SARS-CoV-2 [2]. This notion has gained traction following descriptions of an inverse correlation between the size of the naïve pool and the severity of COVID-19 [7,20,21].
In this study, we used an in vitro approach to prime epitope-specific CD8+ T cells from healthy donors stratified by age, ensuring a complete absence of prior exposure to antigens derived from SARS-CoV-2. We found that age was associated with reductions in the diversity and magnitude of functional CD8+ T cell responses elicited against SARS-CoV-2, measured via the recall induction of IFN-γ. Of note, we also found that epitope recognition frequencies were not associated with peptide binding affinities for HLA-A2, consistent with previous work [22,23,24].
In young donors, EV10 elicited substantially more robust CD8+ T cell responses than any of the peptides derived from SARS-CoV-2, most likely reflecting preservation of the naïve pool and the constitutively high precursor frequencies associated with the expression of HLA-A2 [13]. In contrast, viral specificities were barely detectable via tetramer staining on day 10, irrespective of age. Similar findings have been reported previously [25,26]. SARS-CoV-2-specific responses were nonetheless detected functionally. This discrepancy could reflect lower thresholds for activation versus tetramer binding and/or differences in the respective limits of detection [27]. Moreover, the functional data revealed an age-related deficit in the mobilization of effector CD8+ T cells against various epitopes derived from SARS-CoV-2, consistent with the lower T-bet expression frequencies observed among tetramer-labeled CD8+ T cells from old versus young donors at the same time point, an observation that extended to the model antigen EV10.
These collective observations confirmed and extended previous findings based on the use of peptide matrices to assess priming efficacy [13]. In particular, we used a cleaner approach focused on single epitopes to identify functional responses and further incorporated phenotypic assessments to provide additional mechanistic information, namely the expression of HLA-DR, PD-1, and T-bet. Accordingly, we generated a comprehensive dataset showing that age-related changes in the basal activation status and overall size of the naïve T cell pool were linked with an impaired ability to mount de novo CD8+ T cell responses against previously unencountered pathogens, exemplified in this study by SARS-CoV-2.
Immunosenescence is thought to underlie the vulnerability of elderly individuals to COVID-19 [28]. The data presented here revealed a global decline in priming efficacy with age, which led to a reduction in the diversity of functional CD8+ T cell responses elicited in vitro against a panel of typically immunodominant epitopes derived from SARS-CoV-2. Although further studies will be required to link these findings with immune control of viral replication and vaccine efficacy, our work has established a testable mechanistic paradigm that could expedite the development of new interventions designed to enhance and/or preserve the operational integrity of the naïve T cell pool as a means of protecting the elderly against potentially fatal episodes of COVID-19.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/vaccines11010154/s1: Figure S1. Correlation between peptide binding affinity and immunoprevalence.
Author Contributions
Conceptualization, A.C., F.N. and R.G.; methodology, D.P., B.D., E.G., V.A. (Victor Appay) and S.P.; formal analysis, D.P., B.D., E.G. and F.N.; resources, V.A. (Valentina Albanese), S.L.-L., D.A.P. and S.P.; data curation, D.P.; writing—original draft, D.P. and F.N.; writing—review and editing, D.P., D.A.P., V.A. (Victor Appay), A.C., F.N. and R.G.; supervision, D.A.P., S.P., A.C., F.N. and R.G.; project administration, S.P. and F.N.; funding acquisition, D.A.P., S.P. and F.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the ICE COVID-19 Project supported by the University of Ferrara (Bando Anno 2020 per Progetti di Ricerca Finanziati con il Contributo 5 × 1000 Anno 2018) and by the CRASH CAGE One Health TARGET Project at the Center for International Health (CIH), Ludwig-Maximilians-University, Munich. The CIH is supported by the German Academic Exchange Service (DAAD) with funds from the German Federal Ministry for Economic Cooperation and Development (BMZ). D.A.P. was funded by the National Institute for Health Research (COV-LT2-0041).
Institutional Review Board Statement
This study was approved by the Azienda Unità Sanitaria Locale di Ferrara.
Informed Consent Statement
Written informed consent was not required in accordance with institutional guidance and national legislation.
Data Availability Statement
All original data presented in this study are included in the article/Supplementary Materials. Further inquiries should be directed to the corresponding author (nclfnc1@unife.it).
Acknowledgments
We are grateful to Deepak Paudel (Nepal Public Health Association) and Maria Teresa Solis-Soto (O’Higgins University) for helpful discussions and to Maurizio Govoni (Ferrara University Hospital), Roberta Piva (University of Ferrara), and the Ferrara Blood Bank (AVIS) for assistance with sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nicoli, F.; Paudel, D.; Solis-Soto, M.T. Old and new coronaviruses in the elderly. Aging 2021, 13, 12295–12296. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, F.; Solis-Soto, M.T.; Paudel, D.; Marconi, P.; Gavioli, R.; Appay, V.; Caputo, A. Age-related decline of de novo T cell responsiveness as a cause of COVID-19 severity. Geroscience 2020, 42, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Arregoces-Castillo, L.; Fernandez-Nino, J.; Rojas-Botero, M.; Palacios-Clavijo, A.; Galvis-Pedraza, M.; Rincon-Medrano, L.; Pinto-Alvarez, M.; Ruiz-Gomez, F.; Trejo-Valdivia, B. Effectiveness of COVID-19 vaccines in older adults in Colombia: A retrospective, population-based study of the ESPERANZA cohort. Lancet Healthy Longev. 2022, 3, e242–e252. [Google Scholar] [CrossRef] [PubMed]
- Verity, R.; Okell, L.C.; Dorigatti, I.; Winskill, P.; Whittaker, C.; Imai, N.; Cuomo-Dannenburg, G.; Thompson, H.; Walker, P.G.T.; Fu, H.; et al. Estimates of the severity of coronavirus disease 2019: A model-based analysis. Lancet Infect. Dis. 2020, 20, 669–677. [Google Scholar] [CrossRef]
- Hazeldine, J.; Lord, J.M. Innate immunesenescence: Underlying mechanisms and clinical relevance. Biogerontology 2015, 16, 187–201. [Google Scholar] [CrossRef]
- Cunha, L.L.; Perazzio, S.F.; Azzi, J.; Cravedi, P.; Riella, L.V. Remodeling of the immune response with aging: Immunosenescence and its potential impact on COVID-19 immune response. Front. Immunol. 2020, 11, 1748. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Nicoli, F.; Cabral-Piccin, M.P.; Papagno, L.; Gallerani, E.; Fusaro, M.; Folcher, V.; Dubois, M.; Clave, E.; Vallet, H.; Frere, J.J.; et al. Altered basal lipid metabolism underlies the functional impairment of naive CD8+ T cells in elderly humans. J. Immunol. 2022, 208, 562–570. [Google Scholar] [CrossRef]
- Nicoli, F.; Clave, E.; Wanke, K.; von Braun, A.; Bondet, V.; Alanio, C.; Douay, C.; Baque, M.; Lependu, C.; Marconi, P.; et al. Primary immune responses are negatively impacted by persistent herpesvirus infections in older people: Results from an observational study on healthy subjects and a vaccination trial on subjects aged more than 70 years old. EBioMedicine 2022, 76, 103852. [Google Scholar] [CrossRef]
- Briceno, O.; Lissina, A.; Wanke, K.; Afonso, G.; von Braun, A.; Ragon, K.; Miquel, T.; Gostick, E.; Papagno, L.; Stiasny, K.; et al. Reduced naive CD8+ T-cell priming efficacy in elderly adults. Aging Cell 2016, 15, 14–21. [Google Scholar] [CrossRef]
- Westmeier, J.; Paniskaki, K.; Karakose, Z.; Werner, T.; Sutter, K.; Dolff, S.; Overbeck, M.; Limmer, A.; Liu, J.; Zheng, X.; et al. Impaired cytotoxic CD8+ T cell response in elderly COVID-19 patients. mBio 2020, 11, e02243-20. [Google Scholar] [CrossRef]
- Pittet, M.J.; Valmori, D.; Dunbar, P.R.; Speiser, D.E.; Lienard, D.; Lejeune, F.; Fleischhauer, K.; Cerundolo, V.; Cerottini, J.C.; Romero, P. High frequencies of naive Melan-A/MART-1-specific CD8+ T cells in a large proportion of human histocompatibility leukocyte antigen (HLA)-A2 individuals. J. Exp. Med. 1999, 190, 705–715. [Google Scholar] [CrossRef]
- Gallerani, E.; Proietto, D.; Dallan, B.; Campagnaro, M.; Pacifico, S.; Albanese, V.; Marzola, E.; Marconi, P.; Caputo, A.; Appay, V.; et al. Impaired priming of SARS-CoV-2-specific naive CD8+ T cells in older subjects. Front. Immunol. 2021, 12, 693054. [Google Scholar] [CrossRef]
- Price, D.A.; Brenchley, J.M.; Ruff, L.E.; Betts, M.R.; Hill, B.J.; Roederer, M.; Koup, R.A.; Migueles, S.A.; Gostick, E.; Wooldridge, L.; et al. Avidity for antigen shapes clonal dominance in CD8+ T cell populations specific for persistent DNA viruses. J. Exp. Med. 2005, 202, 1349–1361. [Google Scholar] [CrossRef]
- Prakash, S.; Srivastava, R.; Coulon, P.G.; Dhanushkodi, N.R.; Chentoufi, A.A.; Tifrea, D.F.; Edwards, R.A.; Figueroa, C.J.; Schubl, S.D.; Hsieh, L.; et al. Genome-wide B cell, CD4+, and CD8+ T cell epitopes that are highly conserved between human and animal coronaviruses, identified from SARS-CoV-2 as targets for preemptive pan-coronavirus vaccines. J. Immunol. 2021, 206, 2566–2582. [Google Scholar] [CrossRef]
- Lee, E.; Sandgren, K.; Duette, G.; Stylianou, V.V.; Khanna, R.; Eden, J.S.; Blyth, E.; Gottlieb, D.; Cunningham, A.L.; Palmer, S. Identification of SARS-CoV-2 nucleocapsid and spike T-cell epitopes for assessing T-cell immunity. J. Virol. 2021, 95, e02002-20. [Google Scholar] [CrossRef]
- Gutjahr, A.; Papagno, L.; Nicoli, F.; Lamoureux, A.; Vernejoul, F.; Lioux, T.; Gostick, E.; Price, D.A.; Tiraby, G.; Perouzel, E.; et al. Cutting Edge: A dual TLR2 and TLR7 ligand induces highly potent humoral and cell-mediated immune responses. J. Immunol. 2017, 198, 4205–4209. [Google Scholar] [CrossRef]
- Fali, T.; Fabre-Mersseman, V.; Yamamoto, T.; Bayard, C.; Papagno, L.; Fastenackels, S.; Zoorab, R.; Koup, R.A.; Boddaert, J.; Sauce, D.; et al. Elderly human hematopoietic progenitor cells express cellular senescence markers and are more susceptible to pyroptosis. JCI Insight 2018, 3, e95319. [Google Scholar] [CrossRef]
- Zhang, H.; Weyand, C.M.; Goronzy, J.J.; Gustafson, C.E. Understanding T cell aging to improve anti-viral immunity. Curr. Opin. Virol. 2021, 51, 127–133. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’Andrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369, eabc8511. [Google Scholar] [CrossRef]
- Witkowski, J.M. Immune system aging and the aging-related diseases in the COVID-19 era. Immunol. Lett. 2022, 243, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Szeto, C.; Nguyen, A.T.; Lobos, C.A.; Chatzileontiadou, D.S.M.; Jayasinghe, D.; Grant, E.J.; Riboldi-Tunnicliffe, A.; Smith, C.; Gras, S. Molecular basis of a dominant SARS-CoV-2 spike-derived epitope presented by HLA-A*02:01 recognised by a public TCR. Cells 2021, 10, 2646. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, P.; Nguyen, T.H.O.; Rowntree, L.C.; Juno, J.A.; Wheatley, A.K.; Kent, S.J.; Kedzierska, K.; Rossjohn, J.; Petersen, J. Structural basis of biased T cell receptor recognition of an immunodominant HLA-A2 epitope of the SARS-CoV-2 spike protein. J. Biol. Chem. 2021, 297, 101065. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Xiao, C.; Wang, Z.; Zhu, G.; Mao, L.; Chen, X.; Gao, L.; Deng, J.; Su, J.; Su, H.; et al. CD8+ T-cell epitope variations suggest a potential antigen HLA-A2 binding deficiency for spike protein of SARS-CoV-2. Front. Immunol. 2022, 12, 764949. [Google Scholar] [CrossRef]
- Rowntree, L.C.; Petersen, J.; Juno, J.A.; Chaurasia, P.; Wragg, K.; Koutsakos, M.; Hensen, L.; Wheatley, A.K.; Kent, S.J.; Rossjohn, J.; et al. SARS-CoV-2-specific CD8+ T-cell responses and TCR signatures in the context of a prominent HLA-A*24:02 allomorph. Immunol. Cell Biol. 2021, 99, 990–1000. [Google Scholar] [CrossRef]
- Habel, J.R.; Nguyen, T.H.O.; van de Sandt, C.E.; Juno, J.A.; Chaurasia, P.; Wragg, K.; Koutsakos, M.; Hensen, L.; Jia, X.; Chua, B.; et al. Suboptimal SARS-CoV-2-specific CD8+ T cell response associated with the prominent HLA-A*02:01 phenotype. Proc. Natl. Acad. Sci. USA 2020, 117, 24384–24391. [Google Scholar] [CrossRef]
- Ranieri, E.; Popescu, I.; Gigante, M. CTL ELISPOT assay. Methods Mol. Biol. 2014, 1186, 75–86. [Google Scholar] [CrossRef]
- Chen, Y.; Klein, S.L.; Garibaldi, B.T.; Li, H.; Wu, C.; Osevala, N.M.; Li, T.; Margolick, J.B.; Pawelec, G.; Leng, S.X. Aging in COVID-19: Vulnerability, immunity and intervention. Ageing Res. Rev. 2021, 65, 101205. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).