
Needle-Free Devices and CpG-Adjuvanted DNA Improve Anti-HIV Antibody Responses of Both DNA and Modified Vaccinia Ankara-Vectored Candidate Vaccines
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
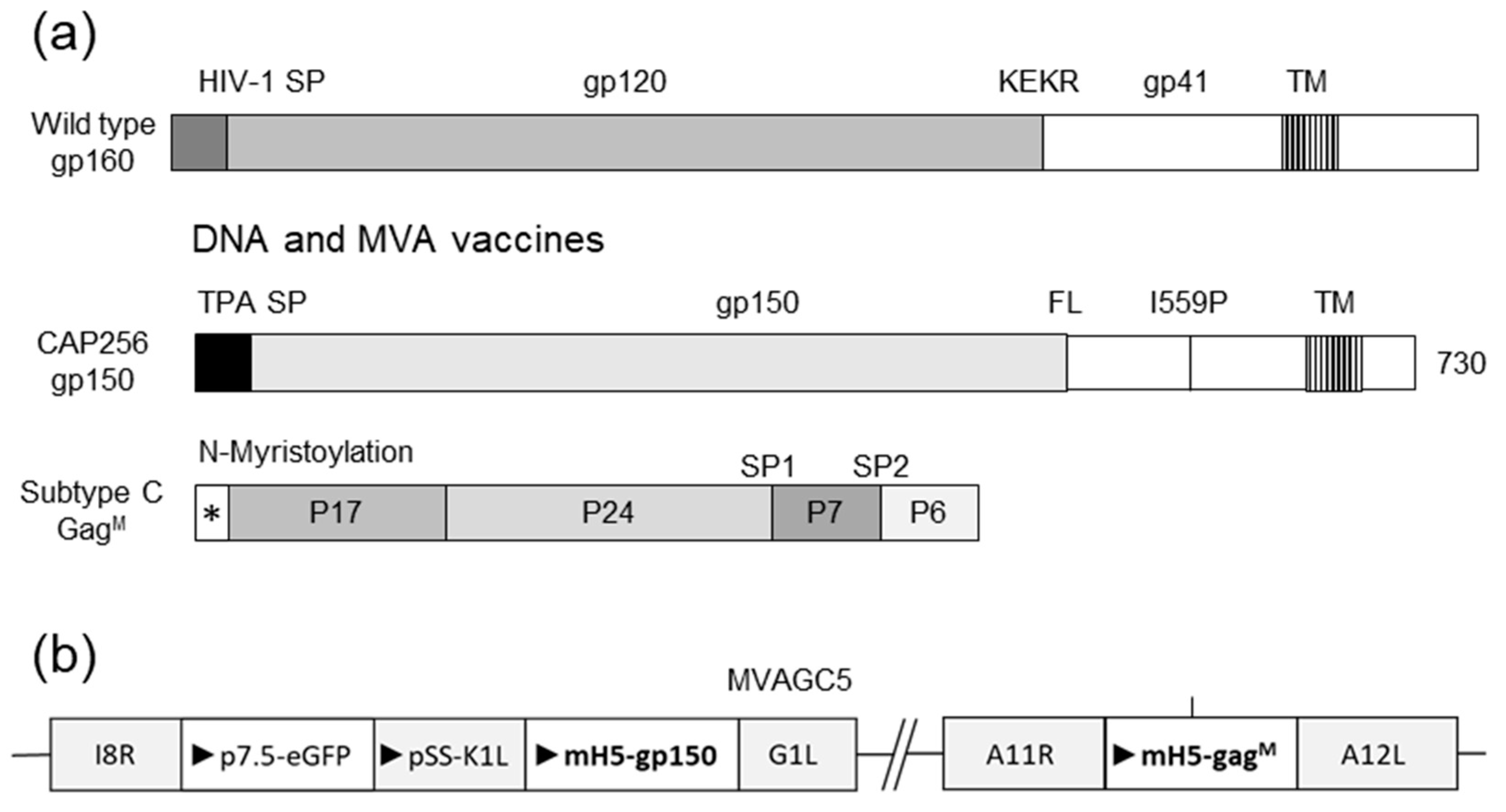
2.1. Vaccines
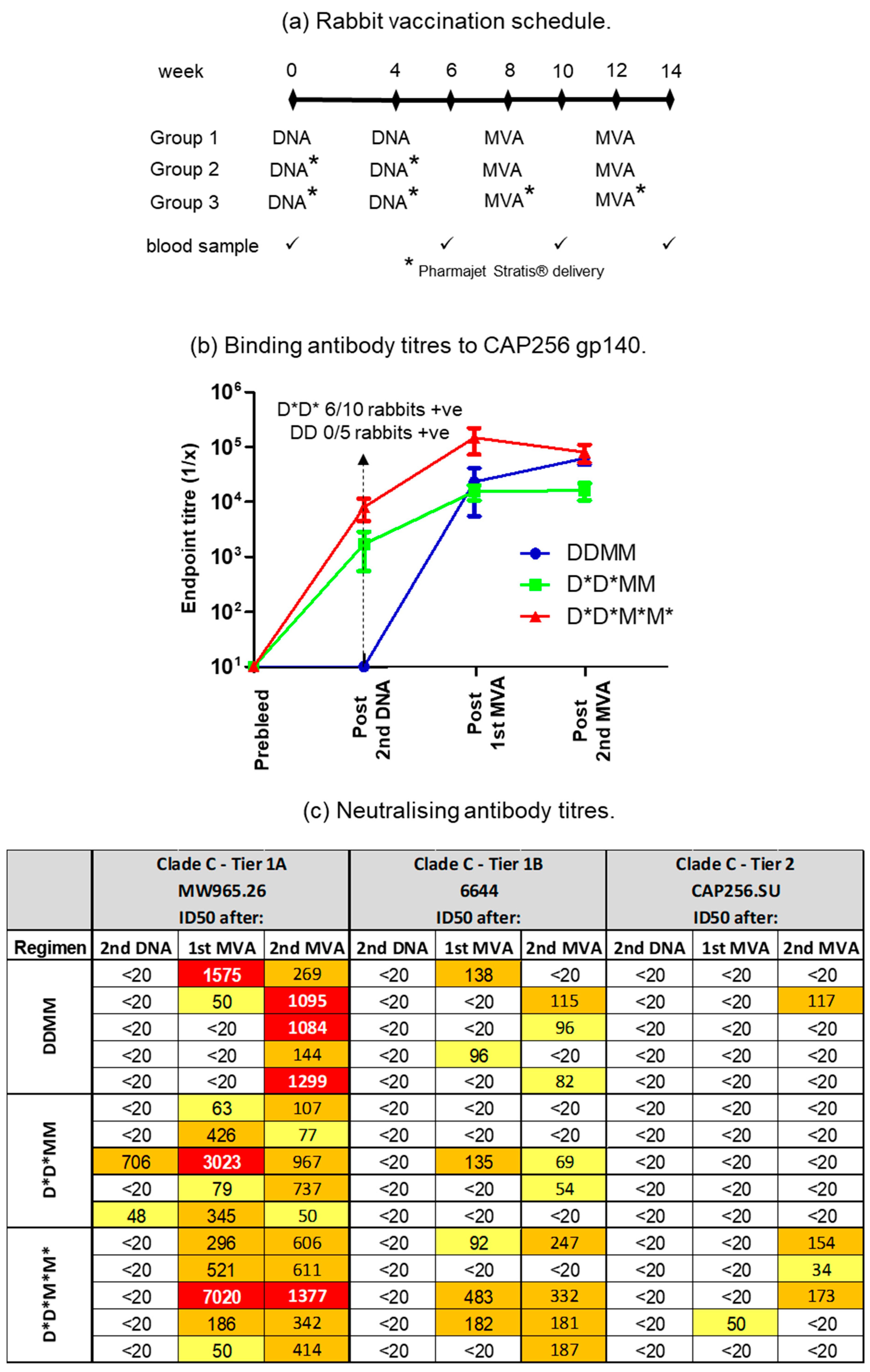
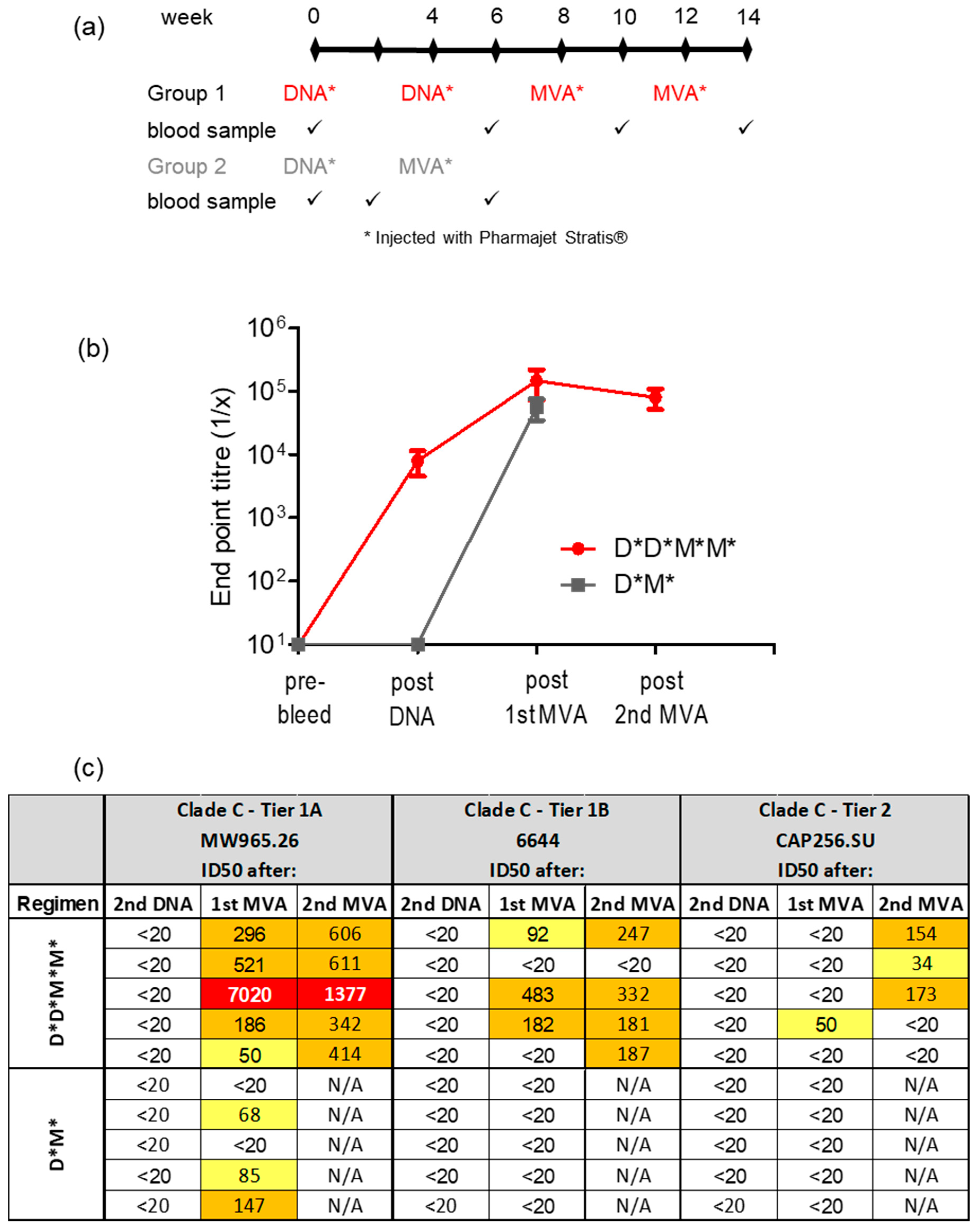
2.2. Rabbit Immunisations
2.3. Anti-Env Enzyme-Linked Immunosorbent Assays (ELISA) and HIV Neutralisation Assays
2.4. Statistical Analysis
3. Results
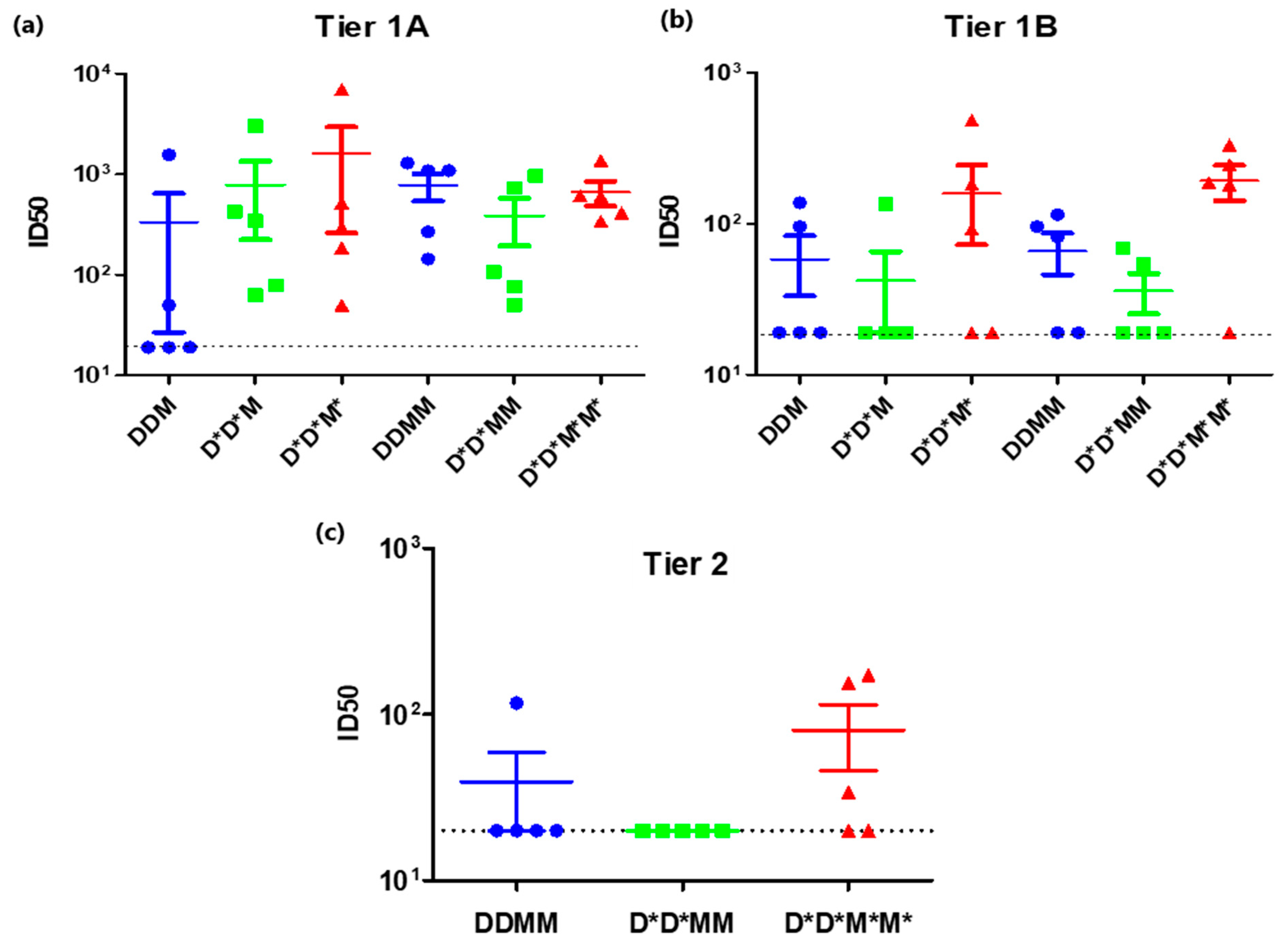
3.1. PharmaJet Stratis Needle-Free Injection Device Improves Immune Responses
3.2. A Shortened Vaccination Regimen Elicits Inferior Neutralizing Antibody Responses
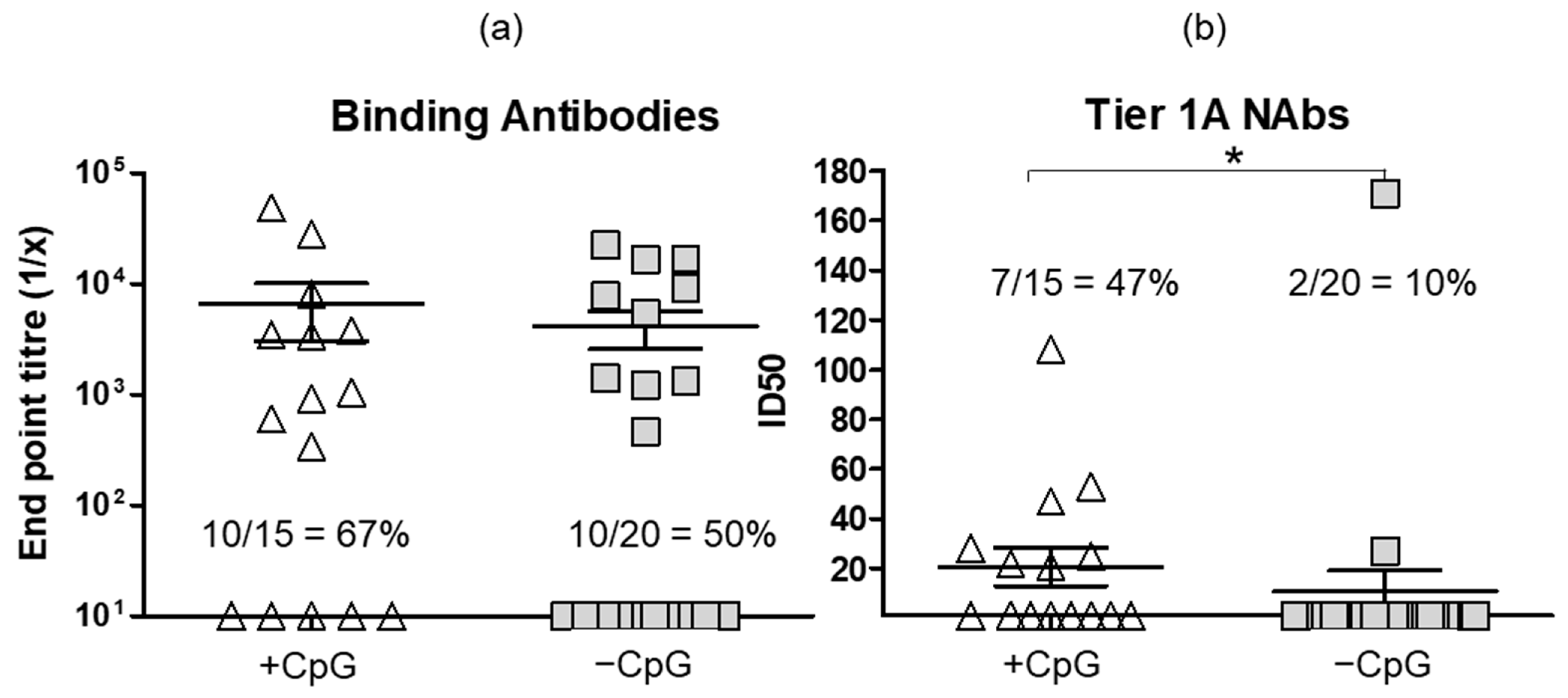
3.3. Administering DNA Vaccines with CpG Adjuvant ODN 1826 Improves Antibody Responses
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuma, K.; Simbayi, L.; Zungu, N.; Moyo, S.; Marinda, E.; Jooste, S.; North, A.; Nadol, P.; Aynalem, G.; Igumbor, E.; et al. The HIV Epidemic in South Africa: Key Findings from 2017 National Population-Based Survey. Int. J. Environ. Res. Public Health 2022, 19, 8125. [Google Scholar] [CrossRef]
- Kim, J.H.; Excler, J.L.; Michael, N.L. Lessons from the RV144 Thai phase III HIV-1 vaccine trial and the search for correlates of protection. Annu. Rev. Med. 2015, 66, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Moodie, Z.; Dintwe, O.; Sawant, S.; Grove, D.; Huang, Y.; Janes, H.; Heptinstall, J.; Omar, F.L.; Cohen, K.; De Rosa, S.C.; et al. Analysis of the HIV Vaccine Trials Network 702 Phase 2b-3 HIV-1 Vaccine Trial in South Africa Assessing RV144 Antibody and T-Cell Correlates of HIV-1 Acquisition Risk. J. Infect. Dis. 2022, 226, 246–257. [Google Scholar] [CrossRef]
- Dispinseri, S.; Tolazzi, M.; Scarlatti, G. The path towards an HIV vaccine. New Microbiol. 2022, 45, 99–103. [Google Scholar]
- Gilbert, P.B.; Huang, Y.; deCamp, A.C.; Karuna, S.; Zhang, Y.; Magaret, C.A.; Giorgi, E.E.; Korber, B.; Edlefsen, P.T.; Rossenkhan, R.; et al. Neutralization titer biomarker for antibody-mediated prevention of HIV-1 acquisition. Nat. Med. 2022, 28, 1924–1932. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Gilbert, P.B.; Juraska, M.; Montefiori, D.C.; Morris, L.; Karuna, S.T.; Edupuganti, S.; Mgodi, N.M.; deCamp, A.C.; Rudnicki, E.; et al. Two Randomized Trials of Neutralizing Antibodies to Prevent HIV-1 Acquisition. N. Engl. J. Med. 2021, 384, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Julg, B.; Sok, D.; Schmidt, S.D.; Abbink, P.; Newman, R.M.; Broge, T.; Linde, C.; Nkolola, J.; Le, K.; Su, D.; et al. Protective Efficacy of Broadly Neutralizing Antibodies with Incomplete Neutralization Activity against Simian-Human Immunodeficiency Virus in Rhesus Monkeys. J. Virol. 2017, 91, e01187-17. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Kirtley, S.; Gouws-Williams, E.; Ghys, P.D.; WHO-UNAIDS Network for HIV Isolation and Characterisation. Global and regional epidemiology of HIV-1 recombinants in 1990–2015: A systematic review and global survey. Lancet HIV 2020, 7, e772–e781. [Google Scholar] [CrossRef]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D.; Characterisation, W.-U.N.f.H.I. Global and regional molecular epidemiology of HIV-1, 1990-2015: A systematic review, global survey, and trend analysis. Lancet Infect. Dis. 2019, 19, 143–155. [Google Scholar] [CrossRef]
- Hemelaar, J.; Loganathan, S.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Kirtley, S.; WHO-UNAIDS Network for HIV Isolation and Characterization. Country Level Diversity of the HIV-1 Pandemic between 1990 and 2015. J. Virol. 2020, 95, e01580-20. [Google Scholar] [CrossRef]
- Barry, H.; Mutua, G.; Kibuuka, H.; Anywaine, Z.; Sirima, S.B.; Meda, N.; Anzala, O.; Eholie, S.; Betard, C.; Richert, L.; et al. Safety and immunogenicity of 2-dose heterologous Ad26.ZEBOV, MVA-BN-Filo Ebola vaccination in healthy and HIV-infected adults: A randomised, placebo-controlled Phase II clinical trial in Africa. PLoS Med. 2021, 18, e1003813. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Moodley, D.; Hanley, S.; Archary, M.; Hoosain, Z.; Lalloo, U.; Louw, C.; Fairlie, L.; Fouche, L.F.; Masilela, M.S.L.; et al. Immunogenicity and safety of a SARS-CoV-2 recombinant spike protein nanoparticle vaccine in people living with and without HIV-1 infection: A randomised, controlled, phase 2A/2B trial. Lancet HIV 2022, 9, e309–e322. [Google Scholar] [CrossRef]
- Martin, C.; Domingo, C.; Hainaut, M.; Delforge, M.; De Wit, S.; Dauby, N. Seroconversion and persistence of neutralizing antibody response after yellow fever vaccination in patients with perinatally acquired HIV infection. AIDS 2023, 37, 341–346. [Google Scholar] [CrossRef]
- Scheepers, C.; Richardson, S.I.; Moyo-Gwete, T.; Moore, P.L. Antibody class-switching as a strategy to improve HIV-1 neutralization. Trends Mol. Med. 2022, 28, 979–988. [Google Scholar] [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Excler, J.L.; Nitayaphan, S.; Kaewkungwal, J.; Premsri, N.; Kunasol, P.; Karasavvas, N.; Schuetz, A.; Ngauy, V.; et al. Randomized, Double-Blind Evaluation of Late Boost Strategies for HIV-Uninfected Vaccine Recipients in the RV144 HIV Vaccine Efficacy Trial. J. Infect. Dis. 2017, 215, 1255–1263. [Google Scholar] [CrossRef]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef]
- Klasse, P.J.; Moore, J.P. Reappraising the Value of HIV-1 Vaccine Correlates of Protection Analyses. J. Virol. 2022, 96, e0003422. [Google Scholar] [CrossRef] [PubMed]
- Mielke, D.; Stanfield-Oakley, S.; Borate, B.; Fisher, L.H.; Faircloth, K.; Tuyishime, M.; Greene, K.; Gao, H.; Williamson, C.; Morris, L.; et al. Selection of HIV Envelope Strains for Standardized Assessments of Vaccine-Elicited Antibody-Dependent Cellular Cytotoxicity-Mediating Antibodies. J. Virol. 2022, 96, e0164321. [Google Scholar] [CrossRef] [PubMed]
- Moyo, N.; Borthwick, N.J.; Wee, E.G.; Capucci, S.; Crook, A.; Dorrell, L.; Hanke, T. Long-term follow up of human T-cell responses to conserved HIV-1 regions elicited by DNA/simian adenovirus/MVA vaccine regimens. PLoS ONE 2017, 12, e0181382. [Google Scholar] [CrossRef]
- Williamson, A.L.; Rybicki, E.P. Justification for the inclusion of Gag in HIV vaccine candidates. Expert Rev. Vaccines 2016, 15, 585–598. [Google Scholar] [CrossRef]
- Fowke, K.R.; Kaul, R.; Rosenthal, K.L.; Oyugi, J.; Kimani, J.; Rutherford, W.J.; Nagelkerke, N.J.; Ball, T.B.; Bwayo, J.J.; Simonsen, J.N.; et al. HIV-1-specific cellular immune responses among HIV-1-resistant sex workers. Immunol. Cell Biol. 2000, 78, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Kaul, R.; Plummer, F.A.; Kimani, J.; Dong, T.; Kiama, P.; Rostron, T.; Njagi, E.; MacDonald, K.S.; Bwayo, J.J.; McMichael, A.J.; et al. HIV-1-specific mucosal CD8+ lymphocyte responses in the cervix of HIV-1-resistant prostitutes in Nairobi. J. Immunol. 2000, 164, 1602–1611. [Google Scholar] [CrossRef]
- van Harmelen, J.H.; Shephard, E.; Thomas, R.; Hanke, T.; Williamson, A.L.; Williamson, C. Construction and characterisation of a candidate HIV-1 subtype C DNA vaccine for South Africa. Vaccine 2003, 21, 4380–4389. [Google Scholar] [CrossRef]
- Gray, G.E.; Mayer, K.H.; Elizaga, M.L.; Bekker, L.G.; Allen, M.; Morris, L.; Montefiori, D.; De Rosa, S.C.; Sato, A.; Gu, N.; et al. Subtype C gp140 Vaccine Boosts Immune Responses Primed by the South African AIDS Vaccine Initiative DNA-C2 and MVA-C HIV Vaccines after More than a 2-Year Gap. Clin. Vaccine Immunol. 2016, 23, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Churchyard, G.; Mlisana, K.; Karuna, S.; Williamson, A.L.; Williamson, C.; Morris, L.; Tomaras, G.D.; De Rosa, S.C.; Gilbert, P.B.; Gu, N.; et al. Sequential Immunization with gp140 Boosts Immune Responses Primed by Modified Vaccinia Ankara or DNA in HIV-Uninfected South African Participants. PLoS ONE 2016, 11, e0161753. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Perkins, S.; Theiler, J.; Bhattacharya, T.; Yusim, K.; Funkhouser, R.; Kuiken, C.; Haynes, B.; Letvin, N.L.; Walker, B.D.; et al. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nat. Med. 2007, 13, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.; Jongwe, T.I.; Douglass, N.; Chege, G.; Williamson, A.L. Heterologous prime-boost vaccination with DNA and MVA vaccines, expressing HIV-1 subtype C mosaic Gag virus-like particles, is highly immunogenic in mice. PLoS ONE 2017, 12, e0173352. [Google Scholar] [CrossRef]
- Doria-Rose, N.A.; Schramm, C.A.; Gorman, J.; Moore, P.L.; Bhiman, J.N.; DeKosky, B.J.; Ernandes, M.J.; Georgiev, I.S.; Kim, H.J.; Pancera, M.; et al. Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature 2014, 509, 55–62. [Google Scholar] [CrossRef]
- Moore, P.L.; Gray, E.S.; Sheward, D.; Madiga, M.; Ranchobe, N.; Lai, Z.; Honnen, W.J.; Nonyane, M.; Tumba, N.; Hermanus, T.; et al. Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol. 2011, 85, 3128–3141. [Google Scholar] [CrossRef]
- van Diepen, M.T.; Chapman, R.; Douglass, N.; Galant, S.; Moore, P.L.; Margolin, E.; Ximba, P.; Morris, L.; Rybicki, E.P.; Williamson, A.L. Prime-Boost Immunizations with DNA, Modified Vaccinia Virus Ankara, and Protein-Based Vaccines Elicit Robust HIV-1 Tier 2 Neutralizing Antibodies against the CAP256 Superinfecting Virus. J. Virol. 2019, 93, e02155-18. [Google Scholar] [CrossRef]
- Sahoo, A.; Jones, A.T.; Cheedarla, N.; Gangadhara, S.; Roy, V.; Styles, T.M.; Shiferaw, A.; Walter, K.L.; Williams, L.D.; Shen, X.; et al. A clade C HIV-1 vaccine protects against heterologous SHIV infection by modulating IgG glycosylation and T helper response in macaques. Sci. Immunol. 2022, 7, eabl4102. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Lai, L.; Wrammert, J.; Kabbani, S.; Xu, Y.; Priyamvada, L.; Hill, H.; Goll, J.B.; Jensen, T.L.; Kao, C.; et al. Plasmablast, Memory B Cell, CD4+ T Cell, and Circulating Follicular Helper T Cell Responses to a Non-Replicating Modified Vaccinia Ankara Vaccine. Vaccines 2020, 8, 69. [Google Scholar] [CrossRef]
- Jackson, L.A.; Frey, S.E.; El Sahly, H.M.; Mulligan, M.J.; Winokur, P.L.; Kotloff, K.L.; Campbell, J.D.; Atmar, R.L.; Graham, I.; Anderson, E.J.; et al. Safety and immunogenicity of a modified vaccinia Ankara vaccine using three immunization schedules and two modes of delivery: A randomized clinical non-inferiority trial. Vaccine 2017, 35, 1675–1682. [Google Scholar] [CrossRef]
- McAllister, L.; Anderson, J.; Werth, K.; Cho, I.; Copeland, K.; Le Cam Bouveret, N.; Plant, D.; Mendelman, P.M.; Cobb, D.K. Needle-free jet injection for administration of influenza vaccine: A randomised non-inferiority trial. Lancet 2014, 384, 674–681. [Google Scholar] [CrossRef]
- Bullo, U.F.; Mehraj, J.; Raza, S.M.; Rasool, S.; Ansari, N.N.; Shaikh, A.A.; Phul, Z.A.; Memon, S.A.; Baloch, R.I.; Baloch, Z.A.; et al. An experience of mass administration of fractional dose inactivated polio vaccine through intradermal needle-free injectors in Karachi, Sindh, Pakistan. BMC Public Health 2021, 21, 44. [Google Scholar] [CrossRef]
- Gaudinski, M.R.; Houser, K.V.; Morabito, K.M.; Hu, Z.; Yamshchikov, G.; Rothwell, R.S.; Berkowitz, N.; Mendoza, F.; Saunders, J.G.; Novik, L.; et al. Safety, tolerability, and immunogenicity of two Zika virus DNA vaccine candidates in healthy adults: Randomised, open-label, phase 1 clinical trials. Lancet 2018, 391, 552–562. [Google Scholar] [CrossRef]
- Kwilas, S.; Kishimori, J.M.; Josleyn, M.; Jerke, K.; Ballantyne, J.; Royals, M.; Hooper, J.W. A hantavirus pulmonary syndrome (HPS) DNA vaccine delivered using a spring-powered jet injector elicits a potent neutralizing antibody response in rabbits and nonhuman primates. Curr. Gene Ther. 2014, 14, 200–210. [Google Scholar] [CrossRef]
- Suschak, J.J.; Bixler, S.L.; Badger, C.V.; Spik, K.W.; Kwilas, S.A.; Rossi, F.D.; Twenhafel, N.; Adams, M.L.; Shoemaker, C.J.; Spiegel, E.; et al. A DNA vaccine targeting VEE virus delivered by needle-free jet-injection protects macaques against aerosol challenge. NPJ Vaccines 2022, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Jin, S.; Gilmartin, L.; Toth, I.; Hussein, W.M.; Stephenson, R.J. Advances in Infectious Disease Vaccine Adjuvants. Vaccines 2022, 10, 1120. [Google Scholar] [CrossRef]
- Facciola, A.; Visalli, G.; Lagana, A.; Di Pietro, A. An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines 2022, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Kayraklioglu, N.; Horuluoglu, B.; Klinman, D.M. CpG Oligonucleotides as Vaccine Adjuvants. Methods Mol. Biol. 2021, 2197, 51–85. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.; Pandey, R.M.; Seth, P. Augmentation of HIV-1 subtype C vaccine constructs induced immune response in mice by CpG motif 1826-ODN. Viral. Immunol. 2005, 18, 213–223. [Google Scholar] [CrossRef]
- Pun, P.B.; Bhat, A.A.; Mohan, T.; Kulkarni, S.; Paranjape, R.; Rao, D.N. Intranasal administration of peptide antigens of HIV with mucosal adjuvant CpG ODN coentrapped in microparticles enhances the mucosal and systemic immune responses. Int. Immunopharmacol. 2009, 9, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.L.; Sheward, D.; Nonyane, M.; Ranchobe, N.; Hermanus, T.; Gray, E.S.; Abdool Karim, S.S.; Williamson, C.; Morris, L. Multiple pathways of escape from HIV broadly cross-neutralizing V2-dependent antibodies. J. Virol. 2013, 87, 4882–4894. [Google Scholar] [CrossRef]
- Sharma, S.K.; de Val, N.; Bale, S.; Guenaga, J.; Tran, K.; Feng, Y.; Dubrovskaya, V.; Ward, A.B.; Wyatt, R.T. Cleavage-independent HIV-1 Env trimers engineered as soluble native spike mimetics for vaccine design. Cell Rep. 2015, 11, 539–550. [Google Scholar] [CrossRef]
- Sanders, R.W.; Vesanen, M.; Schuelke, N.; Master, A.; Schiffner, L.; Kalyanaraman, R.; Paluch, M.; Berkhout, B.; Maddon, P.J.; Olson, W.C.; et al. Stabilization of the soluble, cleaved, trimeric form of the envelope glycoprotein complex of human immunodeficiency virus type 1. J. Virol. 2002, 76, 8875–8889. [Google Scholar] [CrossRef]
- Dowd, K.A.; Ko, S.Y.; Morabito, K.M.; Yang, E.S.; Pelc, R.S.; DeMaso, C.R.; Castilho, L.R.; Abbink, P.; Boyd, M.; Nityanandam, R.; et al. Rapid development of a DNA vaccine for Zika virus. Science 2016, 354, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Lassauniere, R.; Polacek, C.; Gram, G.J.; Frische, A.; Tingstedt, J.L.; Kruger, M.; Dorner, B.G.; Cook, A.; Brown, R.; Orekov, T.; et al. Preclinical evaluation of a candidate naked plasmid DNA vaccine against SARS-CoV-2. NPJ Vaccines 2021, 6, 156. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jiang, S.; Ye, S.; Deng, Y.; Ma, S.; Li, C.P. CpG oligodeoxynucleotide ligand potentiates the activity of the pVAX1-Sj26GST. Biomed. Rep. 2013, 1, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Khodadadi, M.; Ghaffarifar, F.; Dalimi, A.; Ahmadpour, E. Immunogenicity of in-silico designed multi-epitope DNA vaccine encoding SAG1, SAG3 and SAG5 of Toxoplasma gondii adjuvanted with CpG-ODN against acute toxoplasmosis in BALB/c mice. Acta Trop. 2021, 216, 105836. [Google Scholar] [CrossRef]
- Yoshimatsu, H.; Kataoka, K.; Fujihashi, K.; Miyake, T.; Ono, Y. A nasal double DNA adjuvant system induces atheroprotective IgM antibodies via dendritic cell-B-1a B cell interactions. Vaccine 2022, 40, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Becker, E.; Stallmann, S.; Waldhuber, A.; Rommler-Dreher, F.; Albrecht, S.; Mohr, F.; Hegemann, J.H.; Miethke, T. Vaccination with the polymorphic membrane protein A reduces Chlamydia muridarum induced genital tract pathology. Vaccine 2017, 35, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Yao, J.; Feng, G. Protective effect of DNA vaccine encoding pseudomonas exotoxin A and PcrV against acute pulmonary P. aeruginosa Infection. PLoS ONE 2014, 9, e96609. [Google Scholar] [CrossRef] [PubMed]
- Jie, J.; Zhang, Y.; Zhou, H.; Zhai, X.; Zhang, N.; Yuan, H.; Ni, W.; Tai, G. CpG ODN1826 as a Promising Mucin1-Maltose-Binding Protein Vaccine Adjuvant Induced DC Maturation and Enhanced Antitumor Immunity. Int. J. Mol. Sci. 2018, 19, 920. [Google Scholar] [CrossRef] [PubMed]
- Babiarova, K.; Kutinova, L.; Zurkova, K.; Krystofova, J.; Brabcova, E.; Hainz, P.; Musil, J.; Nemeckova, S. Immunization with WT1-derived peptides by tattooing inhibits the growth of TRAMP-C2 prostate tumor in mice. J. Immunother. 2012, 35, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.J.; Ma, B.B.; He, W.; Xu, D.; Wang, X.H. CpG oligodeoxynucleotide stimulates protective innate immunity against human renal cell carcinoma xenografted in nude mice. J. Immunother. 2011, 34, 535–541. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapman, R.; van Diepen, M.; Douglass, N.; Hermanus, T.; Moore, P.L.; Williamson, A.-L. Needle-Free Devices and CpG-Adjuvanted DNA Improve Anti-HIV Antibody Responses of Both DNA and Modified Vaccinia Ankara-Vectored Candidate Vaccines. Vaccines 2023, 11, 376. https://doi.org/10.3390/vaccines11020376
Chapman R, van Diepen M, Douglass N, Hermanus T, Moore PL, Williamson A-L. Needle-Free Devices and CpG-Adjuvanted DNA Improve Anti-HIV Antibody Responses of Both DNA and Modified Vaccinia Ankara-Vectored Candidate Vaccines. Vaccines. 2023; 11(2):376. https://doi.org/10.3390/vaccines11020376
Chicago/Turabian StyleChapman, Rosamund, Michiel van Diepen, Nicola Douglass, Tandile Hermanus, Penny L. Moore, and Anna-Lise Williamson. 2023. "Needle-Free Devices and CpG-Adjuvanted DNA Improve Anti-HIV Antibody Responses of Both DNA and Modified Vaccinia Ankara-Vectored Candidate Vaccines" Vaccines 11, no. 2: 376. https://doi.org/10.3390/vaccines11020376