1. Introduction
The COVID-19 pandemic is the most serious medical emergency in contemporary history. With the joint effort of nations worldwide, effective vaccines were developed in a short time. Two mRNA-based vaccines, Comirnaty (BNT162b2) and Spikevax (mRNA-1273), have been shown to be considerably safer than adenovirus-based vaccines, especially regarding thrombotic adverse events with or without thrombocytopenia [
1,
2,
3]. As a result, Comirnaty and Spikevax have become the primary vaccinations in use across Western countries, and their effectiveness in preventing severe COVID-19 and mortality is well known [
4]. Despite the extremely rare adverse events in vaccine recipients, the benefits of vaccination vastly outweigh any risks.
Comirnaty and Spikevax contain an mRNA strand enclosed in lipid nanoparticles (LNPs). These LNPs are quite different in composition and long-term stability at 4 °C [
5,
6,
7]. The mRNA sequence encodes a full-length, stabilized SARS-CoV-2 Spike (S) protein, which comprises the extracellular receptor-binding domain and the COOH-terminal transmembrane domain. Despite coding the same protein, the mRNA sequence used in the Comirnaty and Spikevax vaccines differs [
8].
Given the divergence between the two vaccines, we examined whether there was a difference in the levels of S-protein produced by the vaccines. Specifically, this study aimed to verify for the first time whether Comirnaty and Spikevax vaccines from a real-world context resulted in different in vitro levels of S-protein production.
2. Materials and Methods
Three vaccination centers in Perugia (Italy) provided us with residual vaccines present in vials after administration. The vials were collected between September and December 2021, during the first cycle of vaccination (first/second dose). The content of four vials of each vaccine was pooled under sterile conditions and used within 1 h after administration of the last dose.
The non-adherent K562 and Jurkat human cell lines were purchased from ATCC (University Boulevard, Manassas, VA, USA). Cell lines were cultured in RPMI-1640 medium (Thermo Fisher Scientific, Waltham, MA, USA) containing 10% fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) and antibiotics (Thermo Fisher Scientific, Waltham, MA, USA). At 24 h before treatment with 1 and 10 µL of vaccine, cells exhibiting exponential growth were seeded on a 6-well plate at 0.25 × 106 cells/mL (0.5 × 106 cells in 2 mL). Untreated cells were used as the control. After 24 h of culture at 37 °C in a 5% CO2 atmosphere, cells were centrifuged at 300 rcf. To remove residual cell components, the culture supernatant was centrifuged again and stored at −80 °C. Cells were stained with a monoclonal antibody against the receptor binding domain of the S-protein (clone P05DHuRb; Thermo Fisher Scientific, Waltham, MA, USA) for 30 min. Cell surface expression of the S-protein was evaluated using flow cytometry (Attune™ NxT; Thermo Fisher Scientific, Waltham, MA, USA). Free S-protein in the cell culture supernatant was evaluated by ELISA in all the samples on the same day (Human SARS-CoV-2 RBD ELISA Kit; Thermo Fisher Scientific, Waltham, MA, USA). Four Comirnaty batches and two Spikevax batches were used for the measurement of cell surface S-protein expression, and four Comirnaty batches and four Spikevax batches were used for free S-protein measurement in the supernatant. Differences in S-protein expression were analyzed using the unpaired t-test (Prism 9.5.1; Graphpad software, San Diego, CA, USA).
3. Results
K562 and Jurkat human cell lines were treated with 1 and 10 µL of Comirnaty and Spikevax vaccines. We first assessed any potential cytotoxic effects.
Figures S1 and S2 show that the lower vaccine dose exerted nearly no cytotoxic effects on Jurkat cells and marginal cytotoxic effects on K562 cells. Moreover, Spikevax-treated samples displayed a higher proportion of dead cells compared with Comirnaty, particularly following the higher dose.
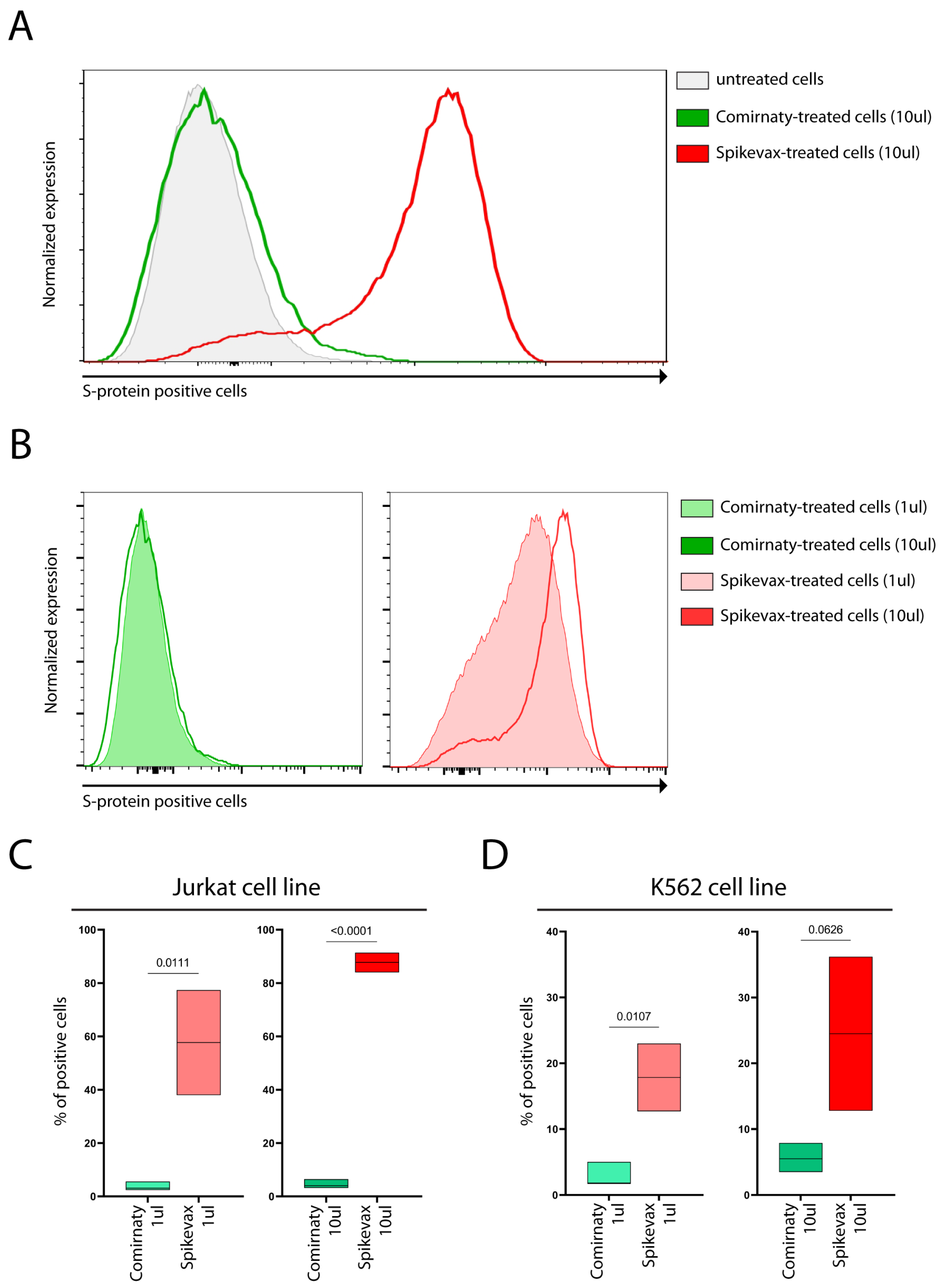
The S-protein was detectable on the cell surface of both cell lines, as shown in
Figure 1A–D, following treatment with both vaccines. Interestingly, a higher percentage of Jurkat cells exhibited cell surface S-protein expression compared with the K562 cells, as shown in
Figure 1C,D.
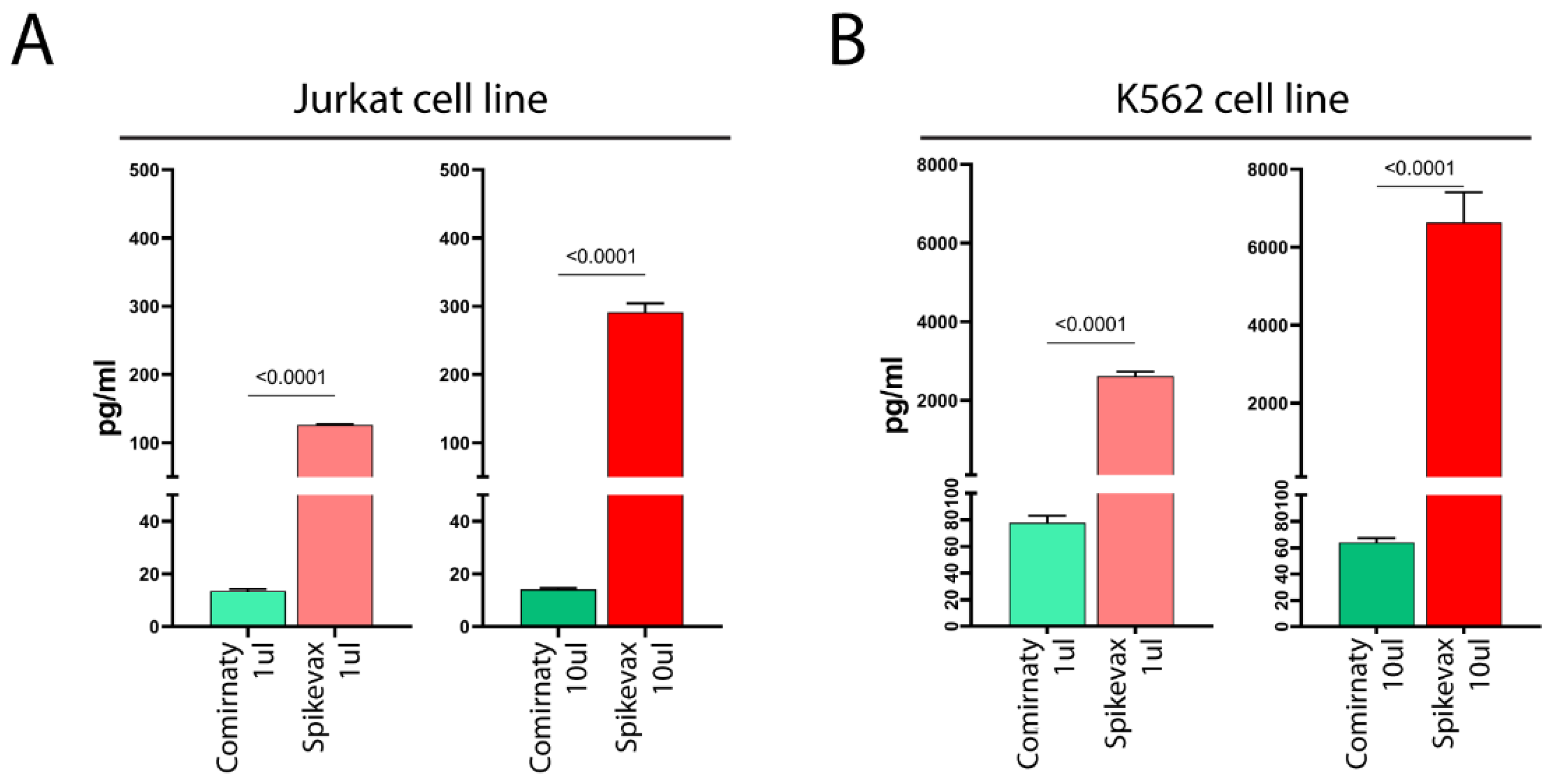
Full-length S-protein, or a truncated form of it, was detected in the supernatant of both vaccine-treated cell lines (
Figure 2A,B), with the K562 cell line showing much higher levels than the Jurkat cell line (
Figure 2A,B). Thus, K562 cells exhibited lower cell surface S-protein expression levels, but higher levels of soluble S-protein than Jurkat cells, possibly suggesting that LNPs of vaccines favored the mRNA entry in both cell lines at similar levels and the observed differences in the detection of S-protein in the cell membrane and culture media reflect differences in the ability of the cells to release the S-protein in the supernatant.
Notably, we found that Spikevax-induced S-protein expression levels were remarkably higher than those of Comirnaty, both on the cell membrane (
Figure 1) and in culture media (
Figure 2), in both cell lines.
4. Discussion
4.1. Differences in Antibody Response and Adverse Events between Comirnaty and Spikevax Vaccines
Our study demonstrates higher S-protein expression levels in cells treated with Spikevax compared with Comirnaty following an in vitro test. Interestingly, both vaccines showed detectable levels of the S-protein (or truncated forms of it) in the culture media.
A vaccine’s effectiveness, including COVID-19 vaccines, depends on numerous variables, including the activity of the pandemic, vaccine recipients’ demographics, lifestyles, and ethnicities [
9]. To compare the efficacy of different vaccines, the only way is to set up a randomized Phase 3 study comparing the same population receiving either the first or the second vaccine. However, no such studies have been performed. Comparative data from real-world studies suggest that Spikevax and Comirnaty vaccines have similar efficacy [
4]. Interestingly, a study on 1647 vaccine recipients found that the anti-S-protein antibody titer after the second dose was approximately three times higher in Spikevax recipients than in Comirnaty recipients [
10]. The higher mRNA dose in Spikevax (100 µg) compared to the Comirnaty dose (30 µg) potentially causes this difference. However, our results showing the different S-protein expressions by cell lines treated with Spikevax and Comirnaty vaccines may contribute to explaining the higher anti-S-protein antibody titer in Spikevax than in Comirnaty recipients. Notably, the differences in S-protein expression levels appear to be independent of the mRNA doses, because S-protein expression levels do not increase as a result of higher Comirnaty doses (
Figure 1B), as discussed in
Section 4.4.
Circulating S-protein has been shown to cause cardiovascular disease by damaging human heart pericytes through CD147 receptor binding and other unknown mechanism(s), regardless of viral infection [
11,
12] and myocarditis is the most common, though extremely rare, adverse event of mRNA-based vaccines [
13]. Recent studies by Yonker et al. have shown that patients with myocarditis following COVID-19 mRNA vaccines had elevated levels of full-length S-protein, unbound by antibodies (free S-protein), in the plasma [
14], agreeing with our findings that the S-protein is present in the supernatant of vaccine-treated cell lines. Interestingly, the full-length free S-protein was only present in vaccine recipients with myocarditis, strongly suggesting that the S-protein is responsible for vaccine-induced myocarditis [
14,
15].
Notably, myocarditis following vaccination with mRNA-based vaccines affects young males much more frequently than other demographics [
13,
16,
17,
18]. In these subjects, Spikevax shows a higher frequency of myocarditis than Comirnaty, with increased risk ranging from 2.5 to 8 folds in different studies [
16,
17,
18]. The higher mRNA dose of Spikevax compared to Comirnaty is believed to be the reason for the increased incidence of myocarditis. If the different in vitro S-protein expressions by Spikevax and Comirnaty vaccines reflect in vivo conditions, our results could contribute to explaining the disparity in myocarditis frequency.
4.2. The Different Compositions of Comirnaty and Spikevax Vaccines May Account for the Varying Levels of S-Protein Expression
Our study did not explore the precise mechanisms underlying the different levels of S-protein expression induced by the Comirnaty and Spikevax vaccines. However, known information about the vaccines’ mRNA and LNP formulations provides some insights.
Both vaccines consist of a 5′-capped single-stranded mRNA encoding the full-length S-protein enclosed within LNPs. The S-protein produced by the two vaccines is identical and differs from the S-protein in the SARS-CoV-2 genome by two amino acids, which stabilize the resulting S-protein in the prefusion state to prime the host immune system to identify the virus before its entry into the host cell. In both vaccines, the mRNA is synthesized by in vitro transcription using a linear DNA template, and, as mammalian host cells attack unmodified exogenous RNA, all U nucleotides of the mRNA were substituted by N1-methylpseudouridine.
Despite these similarities, the Spikevax and Comirnaty mRNAs differ in many other design features, such as the 5′- and 3′-untranslated regions and codon optimization [
8]. Translation initiation, which is the rate-limiting step in translation, depends on the 5′-UTR, whereas if the translation initiation of an mRNA is highly efficient, translation elongation becomes rate-limiting and depends on codon optimization strategies. Therefore, differences in Spikevax and Comirnaty 5′-UTR and codons optimization strategies may be involved in regulating the synthesis rate of S-protein by mammalian cells. Notably, the S-protein mean fluorescence intensity (MFI) was only dose-dependent in Spikevax-treated cells (
Figure 1B), possibly suggesting that mRNA features of Comirnaty may not allow rapid translation initiation and/or elongation becoming limiting for S-protein synthesis and independent of the levels of mRNA delivered into cells. However, more studies are needed.
The LNPs of Comirnaty and Spikevax are quite different [
5,
6]. Although both vaccines’ LNPs contain cholesterol and 1,2-distearoyl-snglycero-3-phosphocholine (DSPC), Comirnaty’s LNP contains ALC-0159 (a PEGylated lipid), whereas Spikevax’s LNP includes another PEGylated lipid (1,2-dimyristoyl-rac-glycero-3-methoxyPEG2000). Both contribute to nanoparticle stabilization [
7]. A fourth LNP component is ALC-0315 [(4-hydroxybutyl)azanediyl)bis(hexane-6,1-diyl)bis(2-hexyldecanoate)] in Comirnaty’s LNP. ALC-0315 is an ionizable amino lipid responsible for mRNA compaction and aids mRNA cellular delivery [
7]. The fourth LNP component is not mentioned by Spikevax manufacturer and, reasonably, differs from ALC-0315. Moreover, various vaccine excipients differ between Comirnaty and Spikevax. Therefore, differences in LNP composition and excipients may account for varying degrees of mRNA protection/integrity and mRNA delivery into cells.
4.3. The Differential Storage Conditions and Required Dilution of Comirnaty and Spikevax May Contribute to Explaining the Variations in S-Protein Expression Levels
It is plausible that the LNP composition and excipients play a role in dictating the different storage conditions for Comirnaty and Spikevax recommended by their respective manufacturers. The unpunctured vials of the latter can be stored at temperatures ranging from −50 °C to −15 °C until expiration, chilled and maintained between 2 °C and 8 °C for up to 30 days, and maintained at 8 °C to 25 °C for 24 h [
19]. Meanwhile, unpunctured vials of Comirnaty can be stored at temperatures ranging from −90 °C to −60 °C until expiration, kept between 2 °C and 8 °C for up to 70 days, and held at 8 °C to 25 °C for 12 h [
20]. The differential requirements of the Spikevax vaccine may suggest greater stability and efficacy in real-world settings, in line with our present study findings.
We observed that the Comirnaty vaccine required dilution prior to administration [
21]. In contrast, there was no requirement for dilution of the Spikevax vaccine [
22]. As Italy is a well-equipped country with well-trained healthcare personnel, we have no reason to conclude that the storage and possible dilution procedures of the vaccines in the tested samples were performed incorrectly. Nevertheless, this observation may hold implications for the LNP stability of Comirnaty in real-world contexts when compared with Spikevax.
4.4. The Differential Levels of In Vitro S-Protein Expression Do Not Depend on the Different mRNA Amounts in Comirnaty and Spikevax Vaccines
As previously stated in
Section 4.1, Comirnaty and Spikevax vaccines utilized different mRNA doses during their administration. Specifically, when the ones that we tested were used (representing the first approved vaccines used in the first cycle of vaccination) adult subjects received 30 µg mRNA in 0.3 mL of Comirnaty and 100 µg mRNA in 0.5 mL of Spikevax. We supposed that vaccine manufacturers decided to use different mRNA doses to obtain similar levels of vaccine-induced S-protein.
To verify this hypothesis, we conducted experiments utilizing 1 and 10 µL of both vaccines. In this way, we tested the effects of 0.1 and 1 µg mRNA present in the Comirnaty vaccine, and 0.2 and 2 µg mRNA present in the Spikevax vaccine. Therefore, when comparing the effects of 1 µL of Comirnaty and Spikevax, we compared the effect of a different dose of Comirnaty (0.1 µg of mRNA) and Spikevax (0.2 µg of mRNA), so that the different levels of S-protein observed on the cell membrane and culture media may be explained by the different mRNA dose. The same reasoning can be applied to the effects of 10 µL of Comirnaty and Spikevax.
However, our results also demonstrate that the varying levels of S-protein expression on the cell membrane and culture media are not solely attributable to differences in mRNA dose. Indeed, when comparing S-protein expression in cell lines treated with 1 µL Spikevax (0.2 µg mRNA) and 10 µL Comirnaty (1 µg mRNA), we observed a substantially lower level of S-protein expression in Comirnaty-treated cells (
Figure 1C,D and
Figure 2), despite the mRNA dose being five-fold higher than in Spikevax-treated cells. Consequently, it can be concluded that differences in S-protein levels following Spikevax and Comirnaty treatment are not due to differences in mRNA dose, but instead result from differences in mRNA sequence and/or LNP composition between the vaccines, as analyzed in
Section 4.1.
Notably, dose escalation of the Spikevax vaccine increases S-protein expression (
Figure 1B (right panel) and
Figure 2), whereas dose escalation of the Comirnaty vaccine does not (
Figure 1B (left panel) and
Figure 2).
4.5. Soluble S-Protein: Possible Explanations
The presence of S-proteins in the supernatant is unexpected given their hydrophobic -COOH terminal transmembrane domain that should not allow protein exit from the cells. Our observations, however, demonstrate the presence of S-protein (or truncated forms of it) in the supernatant, consistent with previous in vivo studies of vaccinated individuals [
14,
23,
24].
Notably, K562 cells exhibited lower cell surface S-protein expression levels but higher levels of soluble S-protein than Jurkat cells, suggesting a varied ability of cells to release S-protein into the supernatant. Therefore, one possible hypothesis is that the S-protein can be shed by cells and the soluble S-protein represents the shed S-protein.
Alternatively, the assessment report of the Spikevax vaccine by the European Medicine Agency (EMA) reports that “the applicant described short mRNA species that can occur because of abortive transcription or premature termination of transcription. As the majority of these short mRNAs do not contain a Poly A tail, the manufacturing process includes chromatography steps, which aim at removing these impurities to a large extent.” However, “additional bands are observed by an in vitro translation assay” performed using the purified mRNA [
6]. A similar problem was reported by Comirnaty’s manufacturer, but, in the case of Comirnaty, the chromatography steps are not performed [
5]. Therefore, it can be hypothesized that some of the short mRNA species present in the LNP of Spikevax and Comirnaty are translated into truncated S-proteins that do not have the transmembrane domain and can exit from the cells, explaining the in vitro finding of the present study and the in vivo finding from other authors [
14,
24].
If the rare adverse events of mRNA-based vaccines are due to the soluble S-protein, further studies are warranted to optimize the safety of mRNA-based vaccines and produce new vaccines even safer than the present ones.
4.6. Study Limitation
Further exploration into the underlying mechanisms responsible for the differential levels of S-protein expression observed with Spikevax and Comirnaty would be valuable, particularly to understand whether these differences are attributable to variations in LNPs, mRNA optimization, or both. However, our study did not test identical LNPs carrying the mRNA of either vaccine or identical mRNA delivered by LNPs present in either Spikevax or Comirnaty.
Additionally, it is important to note that the behavior of cell lines may not accurately reflect that of cells within the human body. In addition, our study utilized cell lines derived from the hematological compartment, with adherent cell lines intentionally omitted to prevent confounding biases due to detachment procedures. However, vaccines are injected into a muscle and their main target may be myocytes.
5. Conclusions
In conclusion, this study has compared the levels of S-protein expression in cells treated with SARS-CoV-2 mRNA-based vaccines used in the real world. Our results demonstrate that the Spikevax vaccine induces a higher level of S-protein expression in cells compared to the Comirnaty vaccine. These differences may be attributed to variations in LNP efficacy favoring mRNA entry in the cells, the mRNA sequence itself and/or loss of LNP and mRNA integrity during transport, storage, or dilution.
The observed findings have significant implications. Firstly, they may help explain the slight differences in the efficacy and safety of the Comirnaty and Spikevax vaccines. Secondly, they suggest the importance of dose–response studies, not only for new drugs but also for new vaccines. Lastly, our results might support the development of new-generation vaccines.
Supplementary Materials
The following supporting information can be downloaded at:
https://www.mdpi.com/article/10.3390/vaccines11040879/s1, Figure S1: Flow cytometric analysis of Jurkat cell line treated with mRNA-based COVID-19 vaccines, Figure S2: cytometric analysis of K562 cell line treated with mRNA-based COVID-19 vaccines.
Author Contributions
L.C. conceptualized the study, performed the experiments, and analyzed and interpreted the data. M.N.A. performed the experiments and analyzed and interpreted the data. A.M. conceptualized the study and discussed the results. G.M. conceptualized the study and discussed the results. G.N. conceptualized the study, discussed the results, and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministero dell’Università e della Ricerca (MUR), grant number FISR2020IP_03103.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We thank the vaccination centers of the USL Umbria 1 district who provided the residual vaccine present in vials after administration and Alfredo Notargiacomo, medical doctor and director of the USL Umbria 1 district, who made the study possible.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cari, L.; Alhosseini, M.N.; Fiore, P.; Pierno, S.; Pacor, S.; Bergamo, A.; Sava, G.; Nocentini, G. Cardiovascular, neurological, and pulmonary events following vaccination with the BNT162b2, ChAdOx1 nCoV-19, and Ad26.COV2.S vaccines: An analysis of European data. J. Autoimmun. 2021, 125, 102742. [Google Scholar] [CrossRef] [PubMed]
- Cari, L.; Naghavi Alhosseini, M.; Bergamo, A.; Pacor, S.; Pierno, S.; Sava, G.; Nocentini, G. Thrombotic events with or without thrombocytopenia in recipients of adenovirus-based COVID-19 vaccines. Front. Cardiovasc. Med. 2022, 9, 967926. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Fernández, M.; Melguizo-Rodríguez, L.; Costela-Ruiz, V.J.; González-Acedo, A.; Ramos-Torrecillas, J.; Illescas-Montes, R. The current status of COVID-19 vaccines. A scoping review. Drug Discov. Today 2022, 27, 103336. [Google Scholar] [CrossRef] [PubMed]
- Higdon, M.M.; Baidya, A.; Walter, K.K.; Patel, M.K.; Issa, H.; Espié, E.; Feikin, D.R.; Knoll, M.D. Duration of effectiveness of vaccination against COVID-19 caused by the omicron variant. Lancet Infect. Dis. 2022, 22, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- EMA—European Medicines Agency. Comirnaty: EPAR—Public Assessment Report. Available online: https://www.ema.europa.eu/en/documents/assessment-report/comirnaty-epar-public-assessment-report_en.pdf (accessed on 14 February 2023).
- EMA—European Medicines Agency. Spikevax (Previously COVID-19 Vaccine Moderna): EPAR—Public Assessment Report. Available online: https://www.ema.europa.eu/en/documents/assessment-report/spikevax-previously-covid-19-vaccine-moderna-epar-public-assessment-report_en.pdf (accessed on 14 February 2023).
- Moghimi, S.M. Allergic Reactions and Anaphylaxis to LNP-Based COVID-19 Vaccines. Mol. Ther. 2021, 29, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. Detailed Dissection and Critical Evaluation of the Pfizer/BioNTech and Moderna mRNA Vaccines. Vaccines 2021, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.-Y.; Li, Y.-D.; Huang, H.-C.; Chan, T.E.H.; Chow, S.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. COVID-19 vaccine update: Vaccine effectiveness, SARS-CoV-2 variants, boosters, adverse effects, and immune correlates of protection. J. Biomed. Sci. 2022, 29, 82. [Google Scholar] [CrossRef] [PubMed]
- Steensels, D.; Pierlet, N.; Penders, J.; Mesotten, D.; Heylen, L. Comparison of SARS-CoV-2 Antibody Response Following Vaccination With BNT162b2 and mRNA-1273. JAMA 2021, 326, 1533. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D. SARS-CoV-2 spike protein causes cardiovascular disease independent of viral infection. Clin. Sci. 2022, 136, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Avolio, E.; Carrabba, M.; Milligan, R.; Kavanagh Williamson, M.; Beltrami, A.P.; Gupta, K.; Elvers, K.T.; Gamez, M.; Foster, R.R.; Gillespie, K.; et al. The SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: A potential non-infective mechanism of COVID-19 microvascular disease. Clin. Sci. 2021, 135, 2667–2689. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Kamat, I.; Hotez, P.J. Myocarditis With COVID-19 mRNA Vaccines. Circulation 2021, 144, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.M.; Swank, Z.; Bartsch, Y.C.; Burns, M.D.; Kane, A.; Boribong, B.P.; Davis, J.P.; Loiselle, M.; Novak, T.; Senussi, Y.; et al. Circulating Spike Protein Detected in Post–COVID-19 mRNA Vaccine Myocarditis. Circulation 2023, 147, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Terpos, E.; Alexopoulos, H.; Politou, M.; Paraskevis, D.; Scorilas, A.; Kastritis, E.; Andreakos, E.; Dimopoulos, M.A. Adverse effects of COVID-19 mRNA vaccines: The spike hypothesis. Trends Mol. Med. 2022, 28, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Karlstad, Ø.; Hovi, P.; Husby, A.; Härkänen, T.; Selmer, R.M.; Pihlström, N.; Hansen, J.V.; Nohynek, H.; Gunnes, N.; Sundström, A.; et al. SARS-CoV-2 Vaccination and Myocarditis in a Nordic Cohort Study of 23 Million Residents. JAMA Cardiol. 2022, 7, 600. [Google Scholar] [CrossRef] [PubMed]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.A.C.; et al. Risk of Myocarditis After Sequential Doses of COVID-19 Vaccine and SARS-CoV-2 Infection by Age and Sex. Circulation 2022, 146, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Heidecker, B.; Dagan, N.; Balicer, R.; Eriksson, U.; Rosano, G.; Coats, A.; Tschöpe, C.; Kelle, S.; Poland, G.A.; Frustaci, A.; et al. Myocarditis following COVID-19 vaccine: Incidence, presentation, diagnosis, pathophysiology, therapy, and outcomes put into perspective. A clinical consensus document supported by the Heart Failure Association of the European Society of Cardio. Eur. J. Heart Fail. 2022, 24, 2000–2018. [Google Scholar] [CrossRef] [PubMed]
- CDC Moderna COVID-19 Vaccine: Storage and Handling Summary. Available online: https://www.cdc.gov/vaccines/covid-19/info-by-product/moderna/downloads/storage-summary.pdf (accessed on 14 February 2023).
- CDC Pfizer-BioNTech COVID-19 Vaccine: Storage and Handling Summary. Available online: https://www.cdc.gov/vaccines/covid-19/info-by-product/pfizer/downloads/storage-summary.pdf (accessed on 14 February 2023).
- EMA—European Medicines Agency. Comirnaty: EPAR—Product Information. Available online: https://www.ema.europa.eu/en/documents/product-information/comirnaty-epar-product-information_en.pdf (accessed on 14 February 2023).
- EMA—European Medicines Agency. Spikevax (Previously COVID-19 Vaccine Moderna): EPAR—Product Information. Available online: https://www.ema.europa.eu/en/documents/product-information/spikevax-previously-covid-19-vaccine-moderna-epar-product-information_en.pdf (accessed on 14 February 2023).
- Bansal, S.; Perincheri, S.; Fleming, T.; Poulson, C.; Tiffany, B.; Bremner, R.M.; Mohanakumar, T. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer–BioNTech) Vaccination prior to Development of Antibodies: A Novel Mechanism for Immune Activation by mRNA Vaccines. J. Immunol. 2021, 207, 2405–2410. [Google Scholar] [CrossRef] [PubMed]
- Ogata, A.F.; Cheng, C.-A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of mRNA-1273 Vaccine Recipients. Clin. Infect. Dis. 2022, 74, 715–718. [Google Scholar] [CrossRef] [PubMed]
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


 ,
, 
{kind=link}
{kind=link}

