
Cross-Neutralization between Bovine Viral Diarrhea Virus (BVDV) Types 1 and 2 after Vaccination with a BVDV-1a Modified-Live-Vaccine

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Inoculum
2.2. Antibody Neutralizing Response
2.3. Statistical Analyses
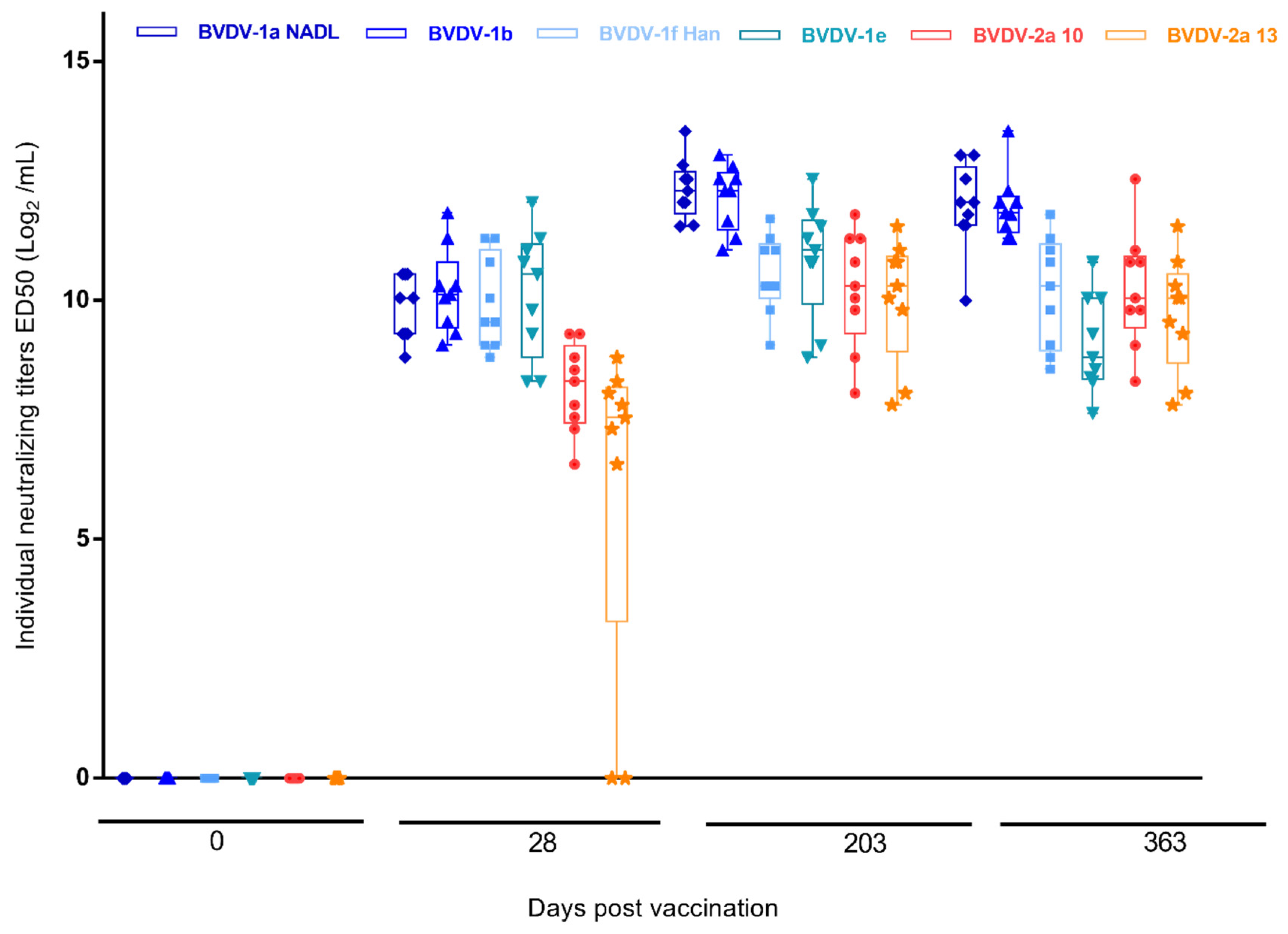
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, D.B.; Meyers, G.; Bukh, J.; Gould, E.A.; Monath, T.; Scott Muerhoff, A.; Pletnev, A.; Rico-Hesse, R.; Stapleton, J.T.; Simmonds, P. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J. Gen. Virol. 2017, 98, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Yeşilbag, K.; Alpay, G.; Becher, P. Variability and global distribution of subgenotypes of bovine viral diarrhea virus. Viruses 2017, 9, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Chen, N.; Guidarini, C.; Xu, Z.; Zhang, J.; Cai, L.; Yuan, S.; Sun, Y.; Metcalfe, L. Prevalence and genetic diversity of bovine viral diarrhea virus in dairy herds of China. Vet. Microbiol. 2020, 242, 108565. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef]
- Fulton, R.W.; Cook, B.J.; Payton, M.E.; Burge, L.J.; Step, D.L. Immune response to Bovine Viral Diarrhea Virus (BVDV) vaccines detecting antibodies to BVDV subtypes 1a, 1b, 2a, and 2c. Vaccine 2020, 38, 4032–4037. [Google Scholar] [CrossRef]
- Platt, R.; Burdett, W.; Roth, J.A. Induction of antigen-specific T-cell subset activation to bovine respiratory disease viruses by a modified-live virus vaccine. Am. J. Vet. Res. 2006, 67, 1179–1184. [Google Scholar] [CrossRef]
- Platt, R.; Kesl, L.; Guidarini, C.; Wang, C.; Roth, J.A. Comparison of humoral and T-cell-mediated immune responses to a single dose of Bovela live double deleted BVDV vaccine or to a field BVDV strain. Vet. Immunol. Immunopathol. 2017, 187, 20–27. [Google Scholar] [CrossRef]
- Theurer, M.E.; Larson, R.L.; White, B.J. Systematic review and meta-analysis of the effectiveness of commercially available vaccines against bovine herpesvirus, Bovine Viral Diarrhea Virus, bovine respiratory syncytial virus, and parainfluenza type 3 virus for mitigation of bovine respiratory disease complex in cattle. J. Am. Vet. Med. Assoc. 2015, 246, 126–142. [Google Scholar]
- Bolin, S.R.; Ridpath, J.F. Assessment of protection from systemic infection or disease a_orded by low to intermediate titers of passively acquired neutralizing antibody against bovine viral diarrhea virus in calves. Am. J. Veter. Res. 1995, 56, 755–759. [Google Scholar]
- Howard, C.J.; Clarke, M.C.; Brownlie, J. Protection against respiratory infection with bovine virus diarrhea virus by passively acquired antibody. Veter. Microbiol. 1989, 19, 195–203. [Google Scholar] [CrossRef]
- Bolin, S.R. Immunogens of bovine viral diarrhea virus. Veter. Microbiol. 1993, 37, 263–271. [Google Scholar] [CrossRef]
- Al-Kubati, A.A.G.; Hussen, J.; Kandeel, M.; Al-Mubarak, A.I.A.; Hemida, M.G. Recent Advances on the Bovine Viral Diarrhea Virus Molecular Pathogenesis, Immune Response, and Vaccines Development. Front. Vet. Sci. 2021, 8, 665128. [Google Scholar] [CrossRef]
- Pecora, A.; Malacari, D.; Ridpath, J.; Aguirreburualde, M.P.; Combessies, G.; Odeón, A.; Romera, S.; Golemba, M.; Wigdorovitz, A. First finding of genetic and antigenic diversity in 1b-BVDV isolates from Argentina. Res. Vet. Sci. 2014, 96, 204–212. [Google Scholar] [CrossRef]
- Alpay, G.; Ye¸silbag, K. Serological relationships among subgroups in bovine viral diarrhea virus genotype 1 (BVDV-1). Vet. Microbiol. 2015, 175, 1–6. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Fulton, R.W.; Kirkland, P.D.; Neill, J.D. Prevalence and antigenic differences observed between bovine viral diarrhea virus subgenotypes isolated from cattle in Australia and feedlots in the Southwestern United States. J. Vet. Diagn. Investig. 2010, 22, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, E.; Righi, C.; Boldini, M.; Bazzucchi, M.; Pezzoni, G.; Gradassi, M.; Petrini, S.; Lelli, D.; Ventura, G.; Pierini, I.; et al. Cross-reactivity antibody response after vaccination with modified live and killed bovine viral diarrhoea virus (BVD) vaccines. Vaccines 2020, 8, 374. [Google Scholar] [CrossRef]
- Agence Nationale de Sécurité Sanitaire de l’Alimentation, de l’Environnement et du Travail, Anses—Index des Médicaments Vétérinaires Autorisés en France. Available online: http://www.ircp.anmv.anses.fr/results.aspx (accessed on 22 November 2021).
- Rivas, J.; Hasanaj, A.; Deblon, C.; Gisbert, P.; Garigliany, M.-M. Genetic diversity of Bovine Viral Diarrhea Virus in cattle in France between 2018 and 2020. Front. Vet. Sci. 2022, 9, 1028866. [Google Scholar] [CrossRef]
- Meyer, G.; Deplanche, M.; Roux, D.; Moulignie, M.; Picard-Hagen, N.; Lyazrhi, F.; Raboisson, D.; Mathevet, P.; Schelcher, F. Fetal protection against bovine viral diarrhoea type 1 virus infection after one administration of a live-attenuated vaccine. Vet. J. 2012, 192, 242–245. [Google Scholar] [CrossRef]
- Achard, D.; Munoz-Bielsa, J.; Pinho, P.; Febery, E. Determination of the fetal protection in pregnant heifers challenged with bovine viral diarrhea type 1 virus twelve months after one administration of a live-attenuated vaccine. In Proceedings of the 30th World Buiatrics Congress, Sapporo, Japan, 27–31 August 2018. [Google Scholar]
- Leyh, R.D.; Fulton, R.W.; Stegner, J.E.; Goodyear, M.D.; Witte, S.B.; Taylor, L.P.; Johnson, B.J.; Step, D.L.; Ridpath, J.F.; Holland, B.P. Fetal protection in heifers vaccinated with a modified-live virus vaccine containing bovine viral diarrhea virus subtypes 1a and 2a and exposed during gestation to cattle persistently infected with bovine viral diarrhea virus subtype 1b. Am. J. Vet. Res. 2011, 72, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Walz, P.H.; Riddell, K.P.; Newcomer, B.W.; Neill, J.D.; Falkenberg, S.M.; Cortese, V.S.; Scruggs, D.W.; Short, T.H. Comparison of reproductive protection against bovine viral diarrhea virus provided by multivalent viral vaccines containing inactivated fractions of bovine viral diarrhea virus 1 and 2. Vaccine 2018, 36, 3853–3860. [Google Scholar] [CrossRef]
- Mosena, A.C.S.; Falkenberg, S.M.; Ma, H.; Casas, E.; Dassanayake, R.P.; Walz, P.H.; Canal, C.W.; Neill, J.D. Multivariate analysis as a method to evaluate antigenic relationships between BVDV vaccine and field strains. Vaccine 2020, 38, 5764–5772. [Google Scholar] [CrossRef] [PubMed]
- Hamers, C.; Di Valentin, E.; Lecomte, C.; Lambot, M.; Joris, E.; Genicot, B.; Pastoret, P.P. Virus neutralising antibodies against 22 bovine viral diarrhoea virus isolates in vaccinated calves. Vet. J. 2002, 163, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Meyer, G.; Combes, M.; Teillaud, A.; Pouget, C.; Bethune, M.-A.; Cassard, H. Vaccination of Sheep with Bovine Viral Diarrhea Vaccines Does Not Protect against Fetal Infection after Challenge of Pregnant Ewes with Border Disease Virus. Vaccines 2021, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Minami, F.; Nagai, M.; Ito, M.; Matsuda, T.; Takai, H.; Jinkawa, Y.; Shimano, T.; Hayashi, M.; Seki, Y.; Sakoda, Y.; et al. Reactivity and prevalence of neutralizing antibodies against Japanese strains of bovine viral diarrhea virus subgenotypes. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 35–39. [Google Scholar] [CrossRef]
- Fulton, R.W.; Ridpath, J.F.; Confer, A.W.; Saliki, J.T.; Burge, L.J.; Payton, M.E. Bovine viral diarrhoea virus antigenic diversity: Impact on disease and vaccination programmes. Biologicals 2003, 31, 89–95. [Google Scholar] [CrossRef]
- Becher, P.; Ramirez, R.A.; Orlich, M.; Rosales, S.C.; Konig, M.; Schweizer, M.; Stalder, H.; Schirrmeier, H.; Theil, H.J. Genetic and antigenic characterization of novel pestivirus genotypes: Implications for classification. Virology 2003, 311, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Bachofen, C.; Stalder, H.; Braun, U.; Hilbe, M.; Ehrensperger, F.; Peterhans, E. Co-existence of genetically and antigenically diverse bovine viral diarrhoea viruses in an endemic situation. Vet. Microbiol. 2008, 131, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Mosena, A.C.S.; Falkenberg, S.M.; Ma, H.; Casas, E.; Dassanayake, R.P.; Booth, R.; De Mia, G.M.; Schwizer, M.; Canal, C.W.; Neill, J.D. Use of multivariate analysis to evaluate antigenic relationships between US BVDV vaccine strains and non-US genetically divergent isolates. J. Virol. Methods 2022, 299, 114328. [Google Scholar] [CrossRef]
- Riitho, V.; Strong, R.; Larska, M.; Graham, S.P.; Steinbach, F. Bovine pestivirus heterogeneity and its potential impact on vaccination and diagnosis. Viruses 2020, 12, 1134. [Google Scholar] [CrossRef]
- Beer, M.; Hehnen, H.R.; Wolfmeyer, A.; Poll, G.; Kaaden, O.R.; Wolf, G. A new inactivated BVDV genotype I and II vaccine. An immunization and challenge study with BVDV genotype, I. Vet. Microbiol. 2000, 77, 195–208. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Neill, J.D.; Frey, M.; Landgraf, J.G. Phylogenetic, antigenic and clinical characterization of type 2 BVDV from North America. Vet. Microbiol. 2000, 77, 145–155. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grange, G.; Mindeguia, M.; Gisbert, P.; Meyer, G. Cross-Neutralization between Bovine Viral Diarrhea Virus (BVDV) Types 1 and 2 after Vaccination with a BVDV-1a Modified-Live-Vaccine. Vaccines 2023, 11, 1204. https://doi.org/10.3390/vaccines11071204
Grange G, Mindeguia M, Gisbert P, Meyer G. Cross-Neutralization between Bovine Viral Diarrhea Virus (BVDV) Types 1 and 2 after Vaccination with a BVDV-1a Modified-Live-Vaccine. Vaccines. 2023; 11(7):1204. https://doi.org/10.3390/vaccines11071204
Chicago/Turabian StyleGrange, Geromine, Marie Mindeguia, Philippe Gisbert, and Gilles Meyer. 2023. "Cross-Neutralization between Bovine Viral Diarrhea Virus (BVDV) Types 1 and 2 after Vaccination with a BVDV-1a Modified-Live-Vaccine" Vaccines 11, no. 7: 1204. https://doi.org/10.3390/vaccines11071204
APA StyleGrange, G., Mindeguia, M., Gisbert, P., & Meyer, G. (2023). Cross-Neutralization between Bovine Viral Diarrhea Virus (BVDV) Types 1 and 2 after Vaccination with a BVDV-1a Modified-Live-Vaccine. Vaccines, 11(7), 1204. https://doi.org/10.3390/vaccines11071204