
Efficacy of Alum-Adjuvanted Peptide and Carbohydrate Conjugate Vaccine Candidates against Group A Streptococcus Pharyngeal Infection in a Non-Human Primate Model

 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Expression and Purification of ADI Protein
2.3. Peptide and Carbohydrate Conjugation to ADI
2.4. Animals
2.5. Immunisation and Challenge
2.6. ELISA
2.7. Statistical Analysis
2.8. Ethics Approvals
3. Results
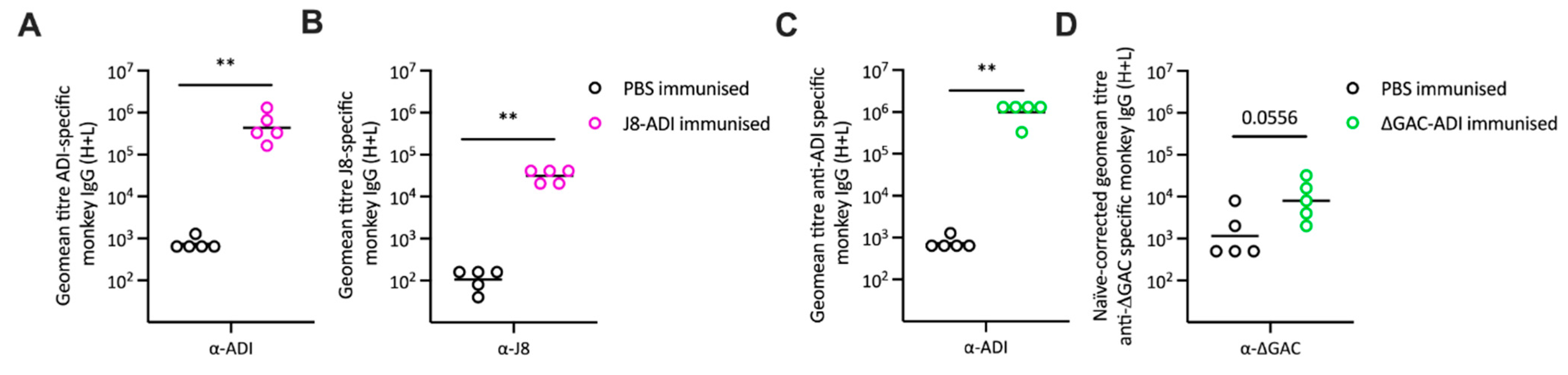
3.1. Antibody Responses to GAS Conjugate Vaccines
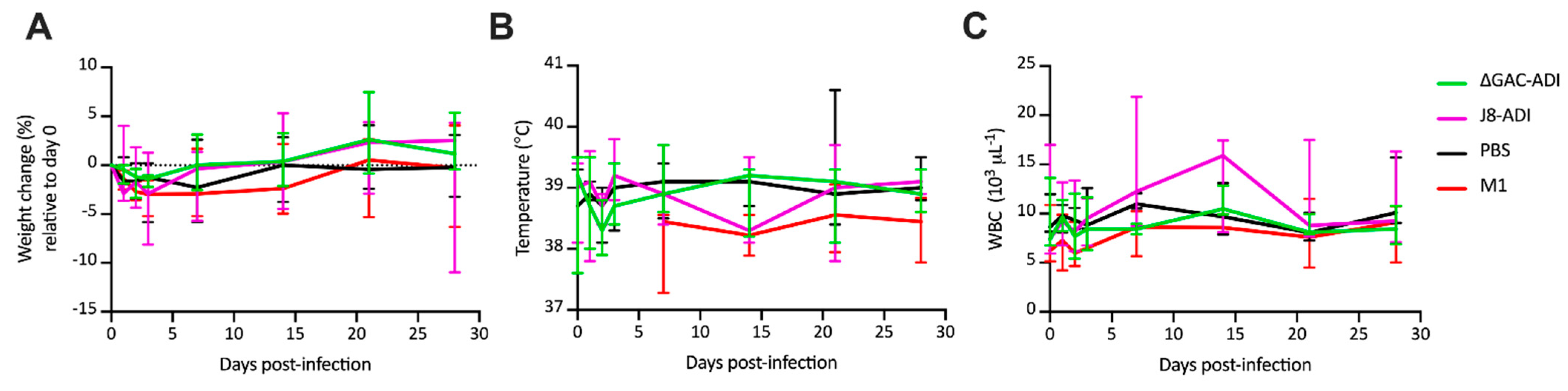
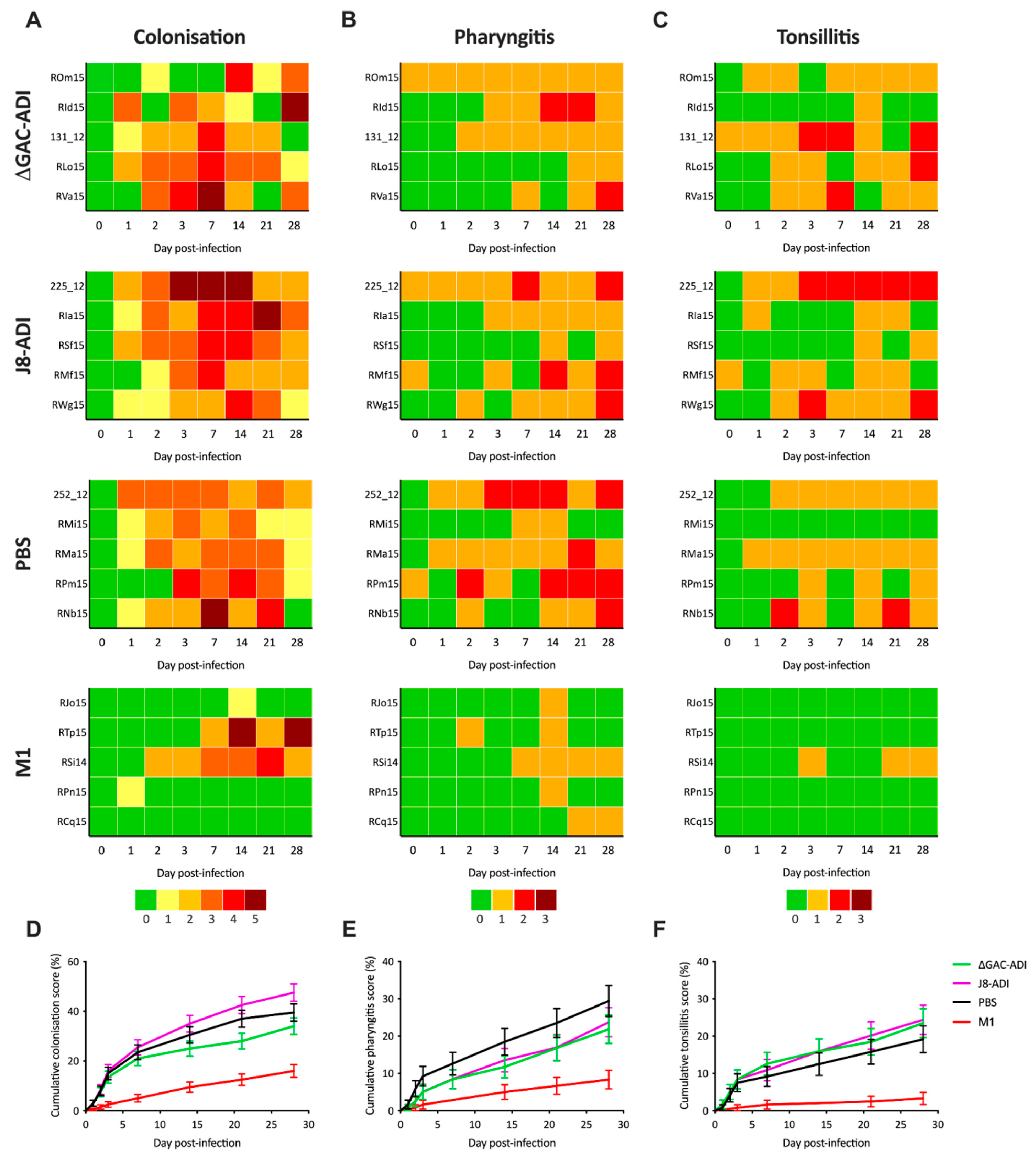
3.2. Efficacy of GAS Conjugate Vaccines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The Global Burden of Group A Streptococcal Diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Watkins, D.A.; Johnson, C.O.; Colquhoun, S.M.; Karthikeyan, G.; Beaton, A.; Bukhman, G.; Forouzanfar, M.H.; Longenecker, C.T.; Mayosi, B.M.; Mensah, G.A.; et al. Global, Regional, and National Burden of Rheumatic Heart Disease, 1990–2015. N. Engl. J. Med. 2017, 377, 713–722. [Google Scholar] [CrossRef]
- Institute for Health Metrics and Evaluation (IHME). University of Washington GBD Results; IHME: Seattle, WA, USA; Available online: http://vizhub.healthdata.org/gbd-results (accessed on 30 June 2022).
- Sheel, M.; Moreland, N.J.; Fraser, J.D.; Carapetis, J. Development of Group A Streptococcal Vaccines: An Unmet Global Health Need. Expert Rev. Vaccines 2016, 15, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Dale, J.B.; Walker, M.J. Update on Group A Streptococcal Vaccine Development. Curr. Opin. Infect. Dis. 2020, 33, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Ajay Castro, S.; Dorfmueller, H.C. Update on the Development of Group A Streptococcus Vaccines. NPJ Vaccines 2023, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S. Conjugate Vaccines. Expert Rev. Vaccines 2003, 2, 633–648. [Google Scholar] [CrossRef]
- Micoli, F.; Del Bino, L.; Alfini, R.; Carboni, F.; Romano, M.R.; Adamo, R. Glycoconjugate Vaccines: Current Approaches towards Faster Vaccine Design. Expert Rev. Vaccines 2019, 18, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, H.; Michon, F.; Nelson, D.; Dong, W.; Fuchs, K.; Manjarrez, R.C.; Sarkar, A.; Uitz, C.; Viteri-Jackson, A.; Suarez, R.S.R.; et al. Group A Streptococcus (GAS) Carbohydrate as an Immunogen for Protection against GAS Infection. J. Infect. Dis. 2006, 193, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, Y.; Wang, G.; Feng, S.; Guo, Z.; Gu, G. Group A Streptococcus Cell Wall Oligosaccharide-Streptococcal C5a Peptidase Conjugates as Effective Antibacterial Vaccines. ACS Infect. Dis. 2020, 6, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, E.; Kis, Z.; Ozanne, J.; Di Benedetto, R.; Ricchetti, B.; Massai, L.; Carducci, M.; Oldrini, D.; Gasperini, G.; Aruta, M.G.; et al. GMMA as an Alternative Carrier for a Glycoconjugate Vaccine against Group A Streptococcus. Vaccines 2022, 10, 1034. [Google Scholar] [CrossRef]
- Rivera-Hernandez, T.; Pandey, M.; Henningham, A.; Cole, J.; Choudhury, B.; Cork, A.J.; Gillen, C.M.; Ghaffar, K.A.; West, N.P.; Silvestri, G.; et al. Differing Efficacies of Lead Group A Streptococcal Vaccine Candidates and Full-Length M Protein in Cutaneous and Invasive Disease Models. MBio 2016, 7, e00618-16. [Google Scholar] [CrossRef] [PubMed]
- Henningham, A.; Ericsson, D.J.; Langer, K.; Casey, L.W.; Jovcevski, B.; Chhatwal, G.S.; Aquilina, J.A.; Batzloff, M.R.; Kobe, B.; Walker, M.J. Structure-Informed Design of an Enzymatically Inactive Vaccine Component for Group A Streptococcus. MBio 2013, 4, e00509-13. [Google Scholar] [CrossRef] [PubMed]
- Good, M.F.; Pandey, M.; Batzloff, M.R.; Tyrrell, G.J. Strategic Development of the Conserved Region of the M Protein and Other Candidates as Vaccines to Prevent Infection with Group A Streptococci. Expert Rev. Vaccines 2015, 14, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Sekuloski, S.; Batzloff, M.R.; Griffin, P.; Parsonage, W.; Elliott, S.; Hartas, J.; O’Rourke, P.; Marquart, L.; Pandey, M.; Rubin, F.A.; et al. Evaluation of Safety and Immunogenicity of a Group A Streptococcus Vaccine Candidate (MJ8VAX) in a Randomized Clinical Trial. PLoS ONE 2018, 13, e0198658. [Google Scholar] [CrossRef] [PubMed]
- van Sorge, N.M.; Cole, J.N.; Kuipers, K.; Henningham, A.; Aziz, R.K.; Kasirer-Friede, A.; Lin, L.; Berends, E.T.M.; Davies, M.R.; Dougan, G.; et al. The Classical Lancefield Antigen of Group a Streptococcus Is a Virulence Determinant with Implications for Vaccine Design. Cell Host Microbe 2014, 15, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.; Rebeyrotte, P.; Parlebas, J.; Halpern, B. Isolation from Heart Valves of Glycopeptides Which Share Immunological Properties with Streptococcus Haemolyticus Group A Polysaccharides. Nature 1968, 219, 866–868. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hernandez, T.; Carnathan, D.G.; Jones, S.; Cork, A.J.; Davies, M.R.; Moyle, P.M.; Toth, I.; Batzloff, M.R.; McCarthy, J.; Nizet, V.; et al. An Experimental Group A Streptococcus Vaccine That Reduces Pharyngitis and Tonsillitis in a Nonhuman Primate Model. MBio 2019, 10, e00693-19. [Google Scholar] [CrossRef] [PubMed]
- Kansal, R.G.; Nizet, V.; Jeng, A.; Chuang, W.-J.; Kotb, M. Selective Modulation of Superantigen-Induced Responses by Streptococcal Cysteine Protease. J. Infect. Dis. 2003, 187, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, T.; Petit Frère, C.; Satzger, M.; Raba, M.; Weisbach, M.; Döhn, K.; Popp, A.; Donzeau, M. Simultaneous Metal Chelate Affinity Purification and Endotoxin Clearance of Recombinant Antibody Fragments. J. Immunol. Methods 2006, 314, 67–73. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011; ISBN 9780309154000. [Google Scholar]
- Skinner, J.M.; Caro-Aguilar, I.C.; Payne, A.M.; Indrawati, L.; Fontenot, J.; Heinrichs, J.H. Comparison of Rhesus and Cynomolgus Macaques in a Streptococcus Pyogenes Infection Model for Vaccine Evaluation. Microb. Pathog. 2011, 50, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Roe, J.H. The Determination of Dextran in Blood and Urine with Anthrone Reagent. J. Biol. Chem. 1954, 208, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, L.; Ferreira, F.M.; Köhler, K.F.; Postol, E.; Kalil, J. A Vaccine against Streptococcus Pyogenes: The Potential to Prevent Rheumatic Fever and Rheumatic Heart Disease. Am. J. Cardiovasc. Drugs 2013, 13, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Postol, E.; Sá-Rocha, L.C.; Sampaio, R.O.; Demarchi, L.M.M.F.; Alencar, R.E.; Abduch, M.C.D.; Kalil, J.; Guilherme, L. Group A Streptococcus Adsorbed Vaccine: Repeated Intramuscular Dose Toxicity Test in Minipigs. Sci. Rep. 2019, 9, 9733. [Google Scholar] [CrossRef] [PubMed]
- Pastural, É.; McNeil, S.A.; MacKinnon-Cameron, D.; Ye, L.; Langley, J.M.; Stewart, R.; Martin, L.H.; Hurley, G.J.; Salehi, S.; Penfound, T.A.; et al. Safety and Immunogenicity of a 30-Valent M Protein-Based Group a Streptococcal Vaccine in Healthy Adult Volunteers: A Randomized, Controlled Phase I Study. Vaccine 2020, 38, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- de Sá-Rocha, L.C.; Demarchi, L.M.M.F.; Postol, E.; Sampaio, R.O.; de Alencar, R.E.; Kalil, J.; Guilherme, L. StreptInCor, a Group A Streptococcal Adsorbed Vaccine: Evaluation of Repeated Intramuscular Dose Toxicity Testing in Rats. Front. Cardiovasc. Med. 2021, 8, 643317. [Google Scholar] [CrossRef] [PubMed]
- Walkinshaw, D.R.; Wright, M.E.E.; Mullin, A.E.; Excler, J.-L.; Kim, J.H.; Steer, A.C. The Streptococcus Pyogenes Vaccine Landscape. NPJ Vaccines 2023, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.M.; Carapetis, J.R.; Van Beneden, C.A.; Cadarette, D.; Daw, J.N.; Moore, H.C.; Bloom, D.E.; Cannon, J.W. The Global Burden of Sore Throat and Group A Streptococcus Pharyngitis: A Systematic Review and Meta-Analysis. eClinicalMedicine 2022, 48, 101458. [Google Scholar] [CrossRef] [PubMed]
- Initiative for Vaccine Research (IVR) of the Department of Immunization, Vaccines and Biologicals. WHO Preferred Product Characteristics for Group a Streptococcus Vaccines; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Schödel, F.; Moreland, N.J.; Wittes, J.T.; Mulholland, K.; Frazer, I.; Steer, A.C.; Fraser, J.D.; Carapetis, J. Clinical Development Strategy for a Candidate Group A Streptococcal Vaccine. Vaccine 2017, 35, 2007–2014. [Google Scholar] [CrossRef] [PubMed]
- Mohan, C.; Ganguly, N.K.; Chakravarti, R.N.; Chitkan, N.L. Prevalence of Haemolytic Streptococcal Infection in Rhesus Monkeys. J. Hyg. Epidemiol. Microbiol. Immunol. 1977, 21, 203–208. [Google Scholar] [PubMed]
- Osowicki, J.; Azzopardi, K.I.; Fabri, L.; Frost, H.R.; Rivera-Hernandez, T.; Neeland, M.R.; Whitcombe, A.L.; Grobler, A.; Gutman, S.J.; Baker, C.; et al. A Controlled Human Infection Model of Streptococcus Pyogenes Pharyngitis (CHIVAS-M75): An Observational, Dose-Finding Study. Lancet Microbe 2021, 2, e291–e299. [Google Scholar] [CrossRef] [PubMed]
- Nanno, Y.; Sterner, E.; Gildersleeve, J.C.; Hering, B.J.; Burlak, C. Profiling Natural Serum Antibodies of Non-Human Primates with a Carbohydrate Antigen Microarray. Xenotransplantation 2020, 27, e12567. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, N.; Uchiyama, S.; Pill, L.; Bautista, L.; Sedra, A.; Yin, L.; Regan, M.; Chu, E.; Rabara, T.; Wong, M.; et al. Non-Native Amino Acid Click Chemistry-Based Technology for Site-Specific Polysaccharide Conjugation to a Bacterial Protein Serving as Both Carrier and Vaccine Antigen. ACS Omega 2022, 7, 24111–24120. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hernandez, T.; Rhyme, M.S.; Cork, A.J.; Jones, S.; Segui-Perez, C.; Brunner, L.; Richter, J.; Petrovsky, N.; Lawrenz, M.; Goldblatt, D.; et al. Vaccine-Induced Th1-Type Response Protects against Invasive Group A Streptococcus Infection in the Absence of Opsonizing Antibodies. MBio 2020, 11, e00122-20. [Google Scholar] [CrossRef] [PubMed]
- Dunne, E.M.; Marshall, J.L.; Baker, C.A.; Manning, J.; Gonis, G.; Danchin, M.H.; Smeesters, P.R.; Satzke, C. Detection of group a streptococcal pharyngitis by quantitative PCR. BMC Infect. Dis. 2013, 13, 312. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Hernandez, T.; Carnathan, D.G.; Richter, J.; Marchant, P.; Cork, A.J.; Elangovan, G.; Henningham, A.; Cole, J.N.; Choudhury, B.; Moyle, P.M.; et al. Efficacy of Alum-Adjuvanted Peptide and Carbohydrate Conjugate Vaccine Candidates against Group A Streptococcus Pharyngeal Infection in a Non-Human Primate Model. Vaccines 2024, 12, 382. https://doi.org/10.3390/vaccines12040382
Rivera-Hernandez T, Carnathan DG, Richter J, Marchant P, Cork AJ, Elangovan G, Henningham A, Cole JN, Choudhury B, Moyle PM, et al. Efficacy of Alum-Adjuvanted Peptide and Carbohydrate Conjugate Vaccine Candidates against Group A Streptococcus Pharyngeal Infection in a Non-Human Primate Model. Vaccines. 2024; 12(4):382. https://doi.org/10.3390/vaccines12040382
Chicago/Turabian StyleRivera-Hernandez, Tania, Diane G. Carnathan, Johanna Richter, Patrick Marchant, Amanda J. Cork, Gayathiri Elangovan, Anna Henningham, Jason N. Cole, Biswa Choudhury, Peter M. Moyle, and et al. 2024. "Efficacy of Alum-Adjuvanted Peptide and Carbohydrate Conjugate Vaccine Candidates against Group A Streptococcus Pharyngeal Infection in a Non-Human Primate Model" Vaccines 12, no. 4: 382. https://doi.org/10.3390/vaccines12040382
APA StyleRivera-Hernandez, T., Carnathan, D. G., Richter, J., Marchant, P., Cork, A. J., Elangovan, G., Henningham, A., Cole, J. N., Choudhury, B., Moyle, P. M., Toth, I., Batzloff, M. R., Good, M. F., Agarwal, P., Kapoor, N., Nizet, V., Silvestri, G., & Walker, M. J. (2024). Efficacy of Alum-Adjuvanted Peptide and Carbohydrate Conjugate Vaccine Candidates against Group A Streptococcus Pharyngeal Infection in a Non-Human Primate Model. Vaccines, 12(4), 382. https://doi.org/10.3390/vaccines12040382