
Evaluating the Compatibility of New Recombinant Protein Antigens (Trivalent NRRV) with a Mock Pentavalent Combination Vaccine Containing Whole-Cell Pertussis: Analytical and Formulation Challenges
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
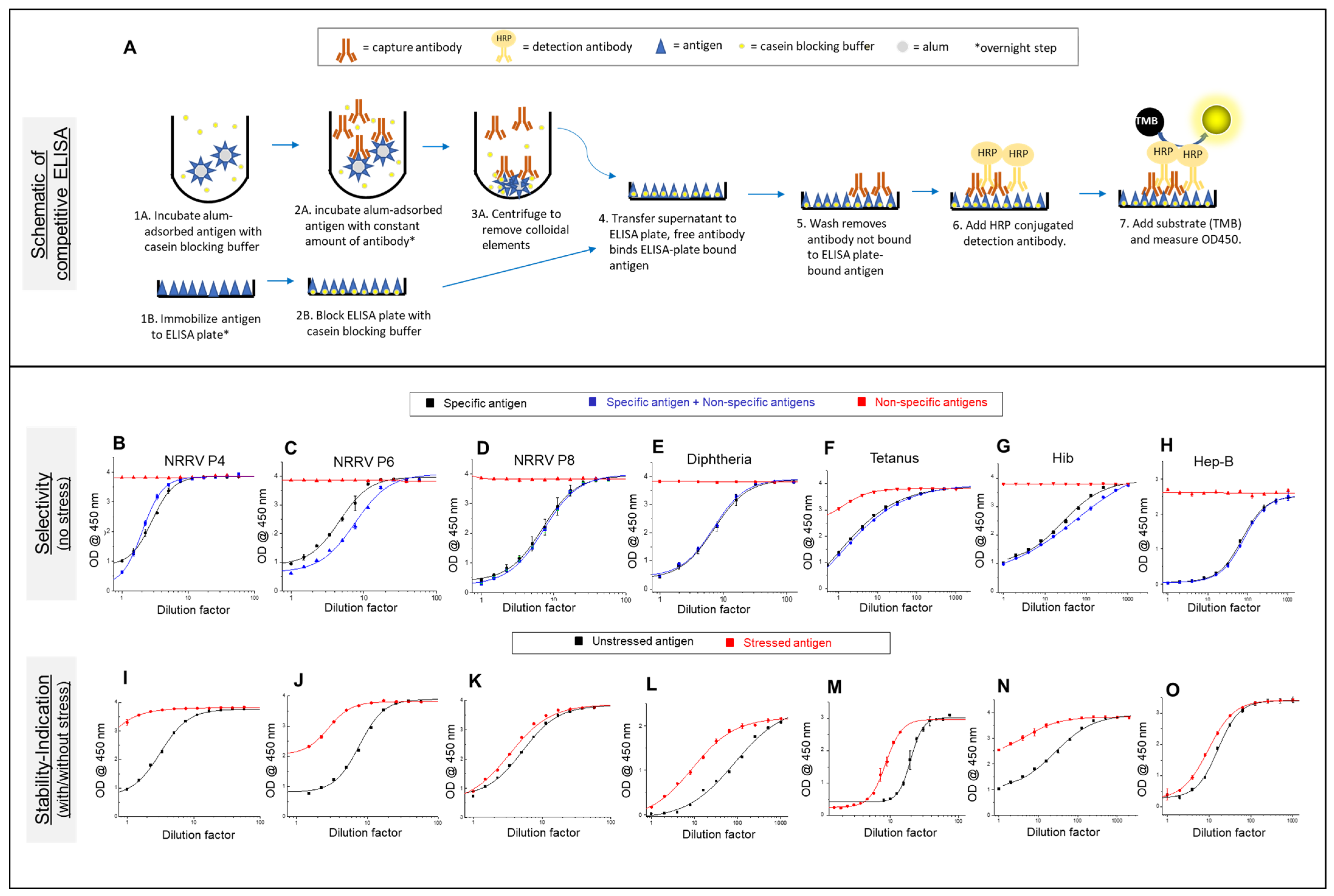
2.2. Competitive ELISA Development for t-NRRV and Four Antigens (D, T, Hib, and HepB) in the Mock Pentavalent Combination
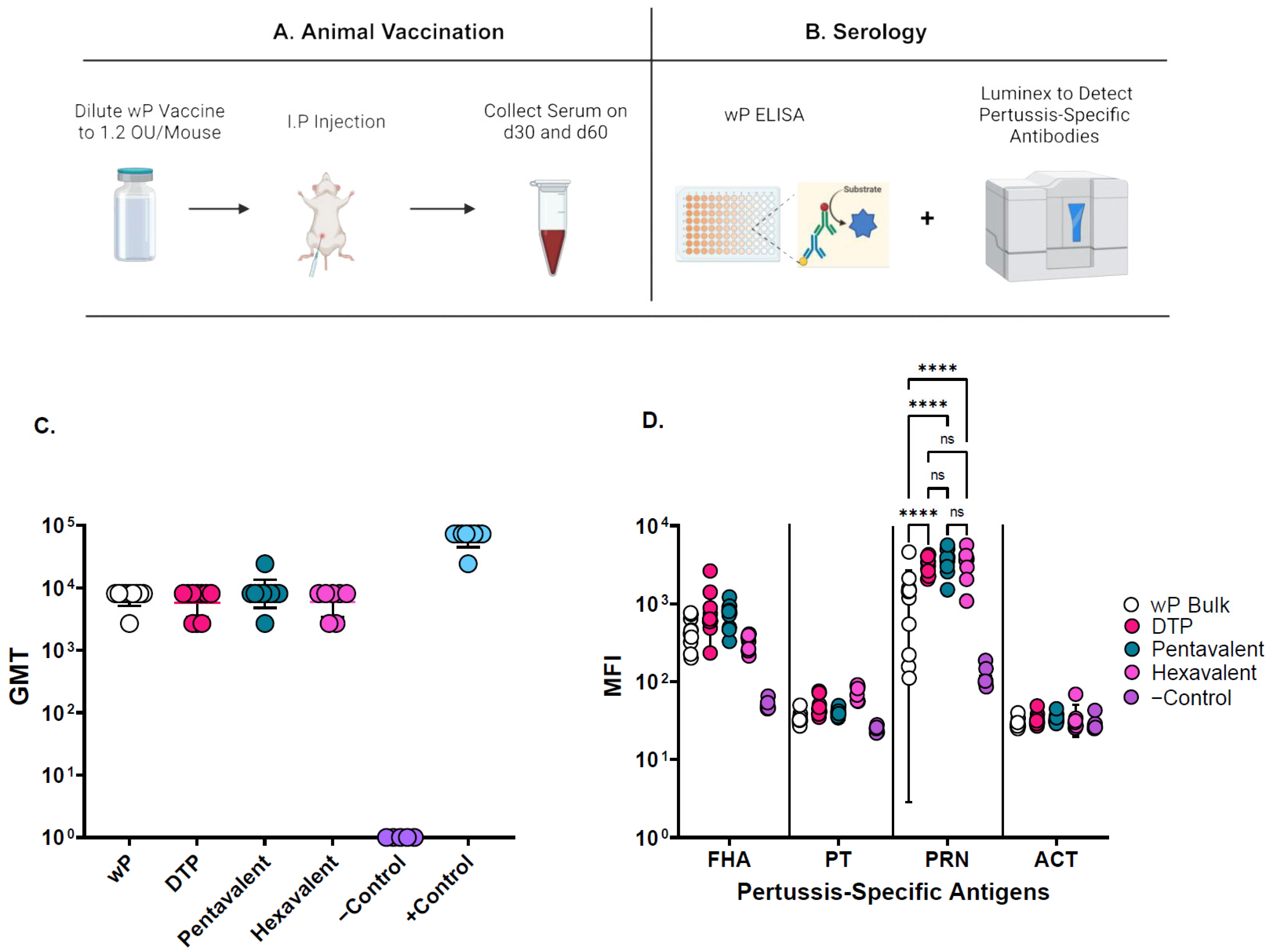
2.3. Mouse Immunogenicity Assay for wP
2.4. wP ELISA
2.5. Multiplex Immune Assay
2.6. Antigen–Aluminum Adjuvant Adsorption Assays
2.7. Compatibility Studies
2.8. Stability Studies
2.9. Stability Profile of AH-Adsorbed t-NRRV Antigens with Preservatives
3. Results
3.1. Competitive ELISA Development for t-NRRV, D, T, Hib, and Hep B Antigens
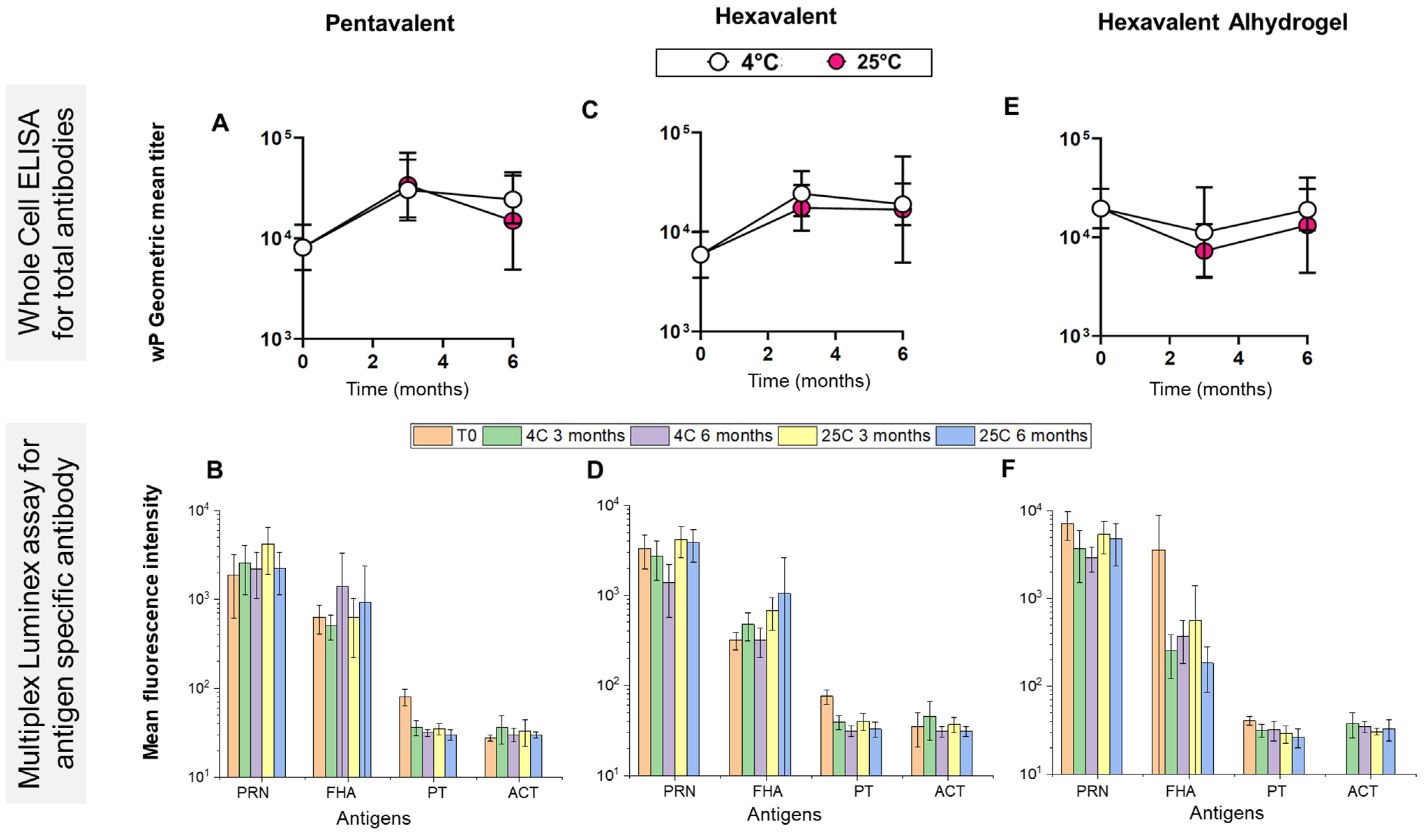
3.2. Mouse Immunogenicity Assays and Adjuvant-Adsorption Assay for wP Antigens
3.3. Compatibility of Aluminum-Salt-Adjuvanted Formulation of t-NRRV and DTwP-Hib-Hep B When Mixed Together
3.4. Mitigation of t-NRRV Desorption from AH Adjuvant upon Mixing with the Pentavalent Formulation
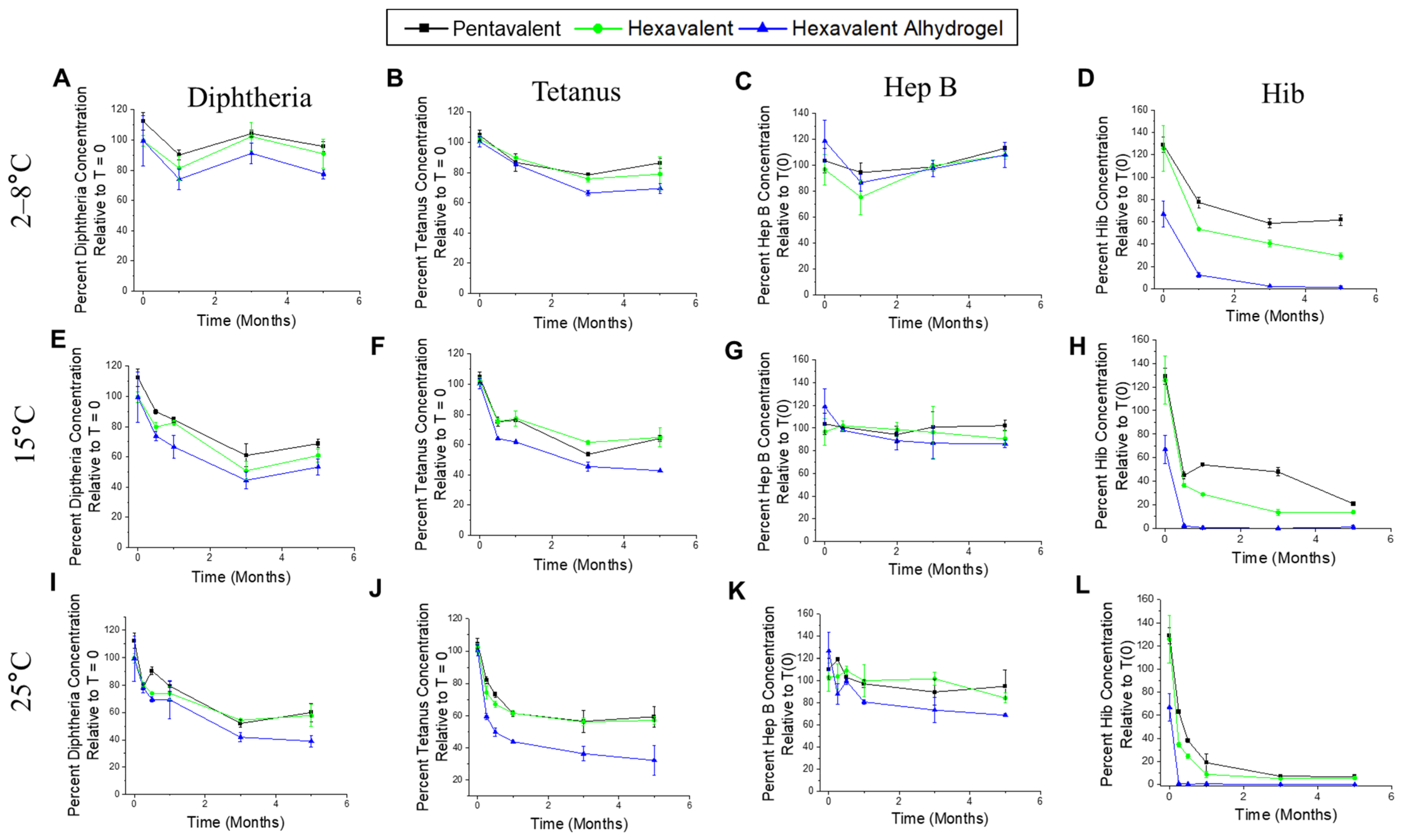
3.5. Real-Time and Accelerated Stability Studies with Various Combination Formulations (t-NRRV + Pentavalent Antigens in the Presence of Aluminum-Salt Adjuvants)
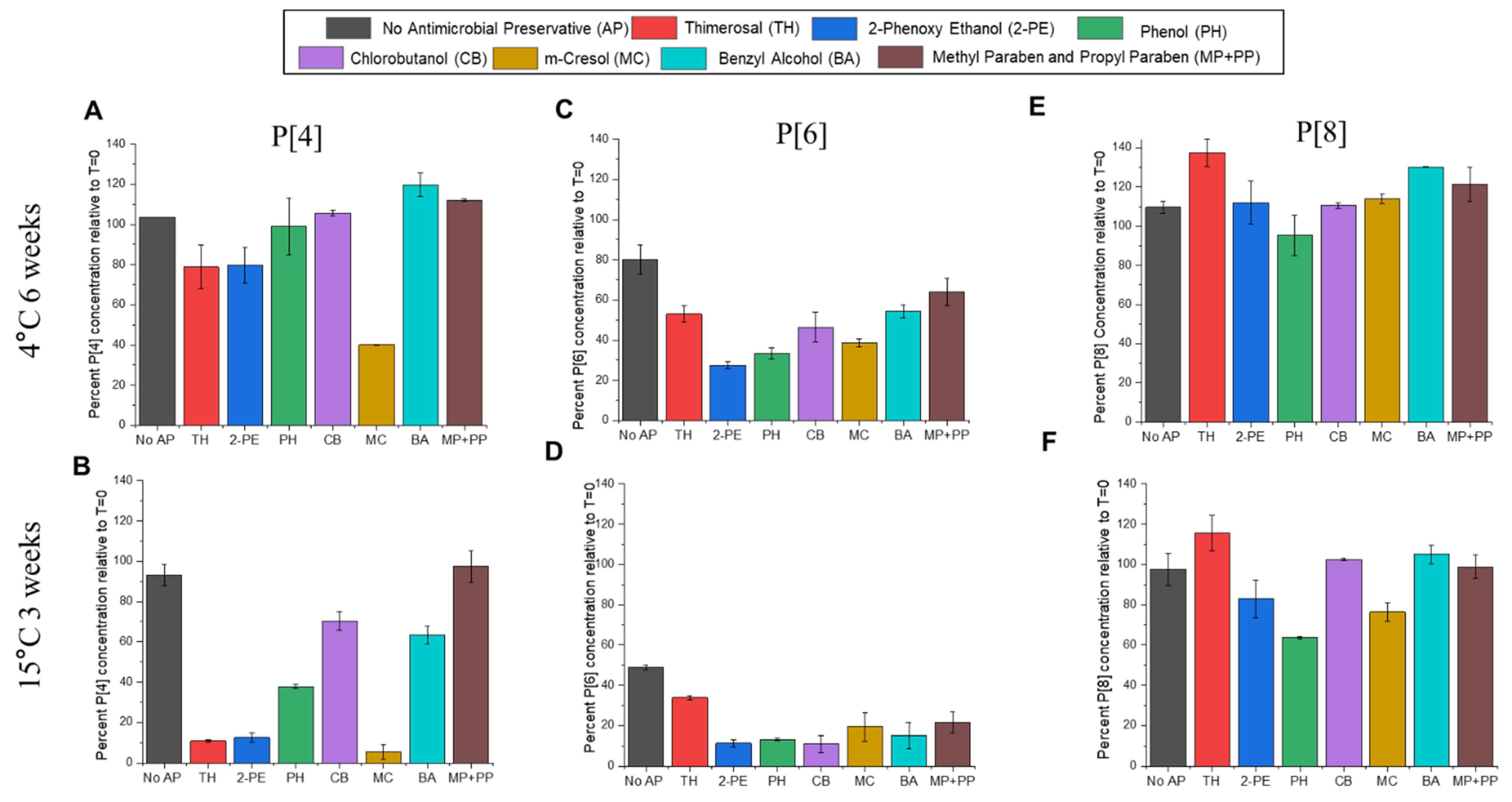
3.6. Effect of Preservatives on AH-Adsorbed t-NRRV as Measured by Competitive ELISA
4. Discussion
4.1. Analytical Challenges and Future Work
4.2. Formulation Challenges and Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skibinski, D.A.; Baudner, B.C.; Singh, M.; O’Hagan, D.T. Combination Vaccines. J. Glob. Infect. Dis. 2011, 3, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Obando-Pacheco, P.; Rivero-Calle, I.; Gómez-Rial, J.; Sánchez, C.R.T.; Martinón-Torres, F. New Perspectives for Hexavalent Vaccines. Vaccine 2018, 36, 5485–5494. [Google Scholar] [CrossRef]
- Ma, J.; Li, Z.; Sun, Y.; Liu, Z.; Dang, Y.; Huang, Y. Improving Innovation and Access to Combination Vaccines for Childhood Immunization in China. Int. J. Environ. Res. Public Health 2022, 19, 15557. [Google Scholar] [CrossRef]
- Shashidhar, A. Hexavalent Vaccinations: The Future of Routine Immunization? Indian. Pediatr. 2017, 54, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; Pelkowski, S.; Atherly, D.; Sitrin, R.; Donnelly, J.J. Hexavalent Ipv-Based Combination Vaccines for Public-Sector Markets of Low-Resource Countries. Hum. Vaccin. Immunother. 2013, 9, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- WHO. Thiomersal. Available online: https://www.who.int/teams/health-product-policy-and-standards/standards-and-specifications/vaccines-quality/thiomersal (accessed on 8 November 2023).
- Kumar, P.; Bird, C.; Holland, D.; Joshi, S.B.; Volkin, D.B. Current and Next-Generation Formulation Strategies for Inactivated Polio Vaccines to Lower Costs, Increase Coverage, and Facilitate Polio Eradication. Hum. Vaccines Immunother. 2022, 18, 2154100. [Google Scholar] [CrossRef]
- WHO. List of Prequalified Vaccines. Available online: https://extranet.who.int/pqweb/vaccines/list-prequalified-vaccines (accessed on 12 September 2023).
- US-FDA. Vaccines. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines (accessed on 12 September 2023).
- Burnett, E.; Parashar, U.; Tate, J. Rotavirus Vaccines: Effectiveness, Safety, and Future Directions. Paediatr. Drugs 2018, 20, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, C.D.; Ma, L.-F.; Carey, M.E.; Steele, A.D. The Rotavirus Vaccine Development Pipeline. Vaccine 2019, 37, 7328–7335. [Google Scholar] [CrossRef]
- Kumar, P.; Pullagurla, S.R.; Patel, A.; Shukla, R.S.; Bird, C.; Kumru, O.S.; Hamidi, A.; Hoeksema, F.; Yallop, C.; Bines, J.E.; et al. Effect of Formulation Variables on the Stability of a Live, Rotavirus (Rv3-Bb) Vaccine Candidate Using in Vitro Gastric Digestion Models to Mimic Oral Delivery. J. Pharm. Sci. 2021, 110, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Magwira, C.A.; Taylor, M.B. Composition of Gut Microbiota and Its Influence on the Immunogenicity of Oral Rotavirus Vaccines. Vaccine 2018, 36, 3427–3433. [Google Scholar] [CrossRef]
- Harris, V.C.; Armah, S.G.; Fuentes, K.E.; Korpela, U.; Parashar, J.C.; Victor, J.; Tate, C.; de Weerth, C.; Giaquinto, W.J.; Wiersinga, K.D.; et al. Significant Correlation between the Infant Gut Microbiome and Rotavirus Vaccine Response in Rural Ghana. J. Infect. Dis. 2017, 215, 34–41. [Google Scholar] [CrossRef]
- Parker, E.P.; Praharaj, I.; Zekavati, A.; Lazarus, R.P.; Giri, S.; Operario, D.J.; Liu, J.; Houpt, E.; Iturriza-Gómara, M.; Kampmann, B.; et al. Influence of the Intestinal Microbiota on the Immunogenicity of Oral Rotavirus Vaccine Given to Infants in South India. Vaccine 2018, 36, 264–272. [Google Scholar] [CrossRef]
- Lazarus, R.P.; John, J.; Shanmugasundaram, E.; Rajan, A.K.; Thiagarajan, S.; Giri, S.; Babji, S.; Sarkar, R.; Kaliappan, P.S.; Venugopal, S.; et al. The Effect of Probiotics and Zinc Supplementation on the Immune Response to Oral Rotavirus Vaccine: A Randomized, Factorial Design, Placebo-Controlled Study among Indian Infants. Vaccine 2018, 36, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Gentsch, J.R.; Glass, R.I. Inactivated Rotavirus Vaccines: A Priority for Accelerated Vaccine Development. Vaccine 2018, 26, 6754–6758. [Google Scholar] [CrossRef]
- O’Ryan, M.; Lopman, B.A. Lopman. Parenteral Protein-Based Rotavirus Vaccine. Lancet Infect. Dis. 2017, 17, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Debellut, F.; Pecenka, C.; Hausdorff, W.P.; Clark, A. Potential Impact and Cost-Effectiveness of Injectable Next-Generation Rotavirus Vaccines in 137 Lmics: A Modelling Study. Hum. Vaccin. Immunother. 2022, 18, 2040329. [Google Scholar] [CrossRef]
- Wen, X.; Cao, D.; Jones, R.W.; Hoshino, Y.; Yuan, L. Tandem Truncated Rotavirus Vp8* Subunit Protein with T Cell Epitope as Non-Replicating Parenteral Vaccine Is Highly Immunogenic. Hum. Vaccin. Immunother. 2015, 11, 2483–2489. [Google Scholar] [CrossRef]
- Wen, X.; Cao, D.; Jones, R.W.; Li, J.; Szu, S.; Hoshino, Y. Construction and Characterization of Human Rotavirus Recombinant Vp8* Subunit Parenteral Vaccine Candidates. Vaccine 2012, 30, 6121–6126. [Google Scholar] [CrossRef] [PubMed]
- PATH. Path Announces Early Closure of Pivotal Phase 3 Study of an Injectable Rotavirus Vaccine Candidate. Available online: https://www.path.org/media-center/path-announces-early-closure-of-pivotal-phase-3-study-of-an-injectable-rotavirus-vaccine-candidate/ (accessed on 12 December 2023).
- Prashant, K.; Atsushi, H.; Christopher, B.; Brandy, D.; Soraia, S.; Sangeeta, B.J.; David, B.V. Evaluating the Compatibility of three Aluminum-Salt Adjuvanted Recombinant Protein Antigens (Trivalent NRRV) Combined with a Mock Trivalent Sabin-IPV: Analytical and Formulation Challenges. Vaccines 2024. manuscript in preparation. [Google Scholar]
- van der Ark, A.A.; Hozbor, D.F.; Boog, C.J.; Metz, B.; Dobbelsteen, G.P.v.D.; van Els, C.A. Resurgence of Pertussis Calls for Re-Evaluation of Pertussis Animal Models. Expert Rev. Vaccines 2012, 11, 1121–1137. [Google Scholar] [CrossRef]
- Jerajani, K.; Wan, Y.; Kumru, O.S.; Pullagurla, S.R.; Kumar, P.; Sharma, N.; Ogun, O.; Mapari, S.; Brendle, S.; Christensen, N.D.; et al. Multi-Dose Formulation Development for a Quadrivalent Human Papillomavirus Virus-like Particle-Based Vaccine: Part I—Screening of Preservative Combinations. J. Pharm. Sci. 2023, 112, 446–457. [Google Scholar] [CrossRef]
- McAdams, D.; Estrada, M.; Holland, D.; Singh, J.; Sawant, N.; Hickey, J.M.; Kumar, P.; Plikaytis, B.; Joshi, S.B.; Volkin, D.B.; et al. Concordance of in Vitro and in Vivo Measures of Non-Replicating Rotavirus Vaccine Potency. Vaccine 2022, 40, 5069–5078. [Google Scholar] [CrossRef] [PubMed]
- McAdams, D.; Lakatos, K.; Estrada, M.; Chen, D.; Plikaytis, B.; Sitrin, R.; White, J.A. Quantification of Trivalent Non-Replicating Rotavirus Vaccine Antigens in the Presence of Aluminum Adjuvant. J. Immunol. Methods 2021, 494, 113056. [Google Scholar] [CrossRef] [PubMed]
- Coombes, L.; Stickings, P.; Tierney, R.; Rigsby, P.; Sesardic, D. Development and Use of a Novel in Vitro Assay for Testing of Diphtheria Toxoid in Combination Vaccines. J. Immunol. Methods 2009, 350, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Coombes, L.; Tierney, R.; Rigsby, P.; Sesardic, D.; Stickings, P. In Vitro Antigen Elisa for Quality Control of Tetanus Vaccines. Biologicals 2012, 40, 466–472. [Google Scholar] [CrossRef] [PubMed]
- AlphaDiagnostics. Anti-Haemophilus Influenzae, Type B (Heat Killed, Whole Bacteria) Antiserum. Available online: https://www.4adi.com/4adi/anti-haemophilus-influenzae-type-b-heat-killed-whole-bacteria-antiserum-24301-p.html (accessed on 13 September 2023).
- BioRad. Hepatitis B Surface Antigen Ad/Ay Antibody. Available online: https://www.bio-rad-antibodies.com/results/go?w=4940-1404 (accessed on 13 September 2023).
- HogenEsch, H.; Dunham, A.; Hansen, B.; Anderson, K.; Maisonneuve, J.-F.; Hem, S.L. Formulation of a Killed Whole Cell Pneumococcus Vaccine—Effect of Aluminum Adjuvants on the Antibody and Il-17 Response. J. Immune Based Ther. Vaccines 2011, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, K.; McAdams, D.; White, J.A.; Chen, D. Formulation and Preclinical Studies with a Trivalent Rotavirus P2-Vp8 Subunit Vaccine. Hum. Vaccin. Immunother. 2020, 16, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Khatuntseva, E.A.; Nifantiev, N.E. Glycoconjugate Vaccines for Prevention of Haemophilus Influenzae Type B Diseases. Russ. J. Bioorg Chem. 2021, 47, 26–52. [Google Scholar] [CrossRef]
- Sturgess, A.W.; Rush, K.; Charbonneau, R.J.; I Lee, J.; West, D.J.; Sitrin, R.D.; Hennessey, J.P. Haemophilus Influenzae Type B Conjugate Vaccine Stability: Catalytic Depolymerization of Prp in the Presence of Aluminum Hydroxide. Vaccine 1999, 17, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Hickey, J.M.; McAdams, D.; White, J.A.; Sitrin, R.; Khandke, L.; Cryz, S.; Joshi, S.B.; Volkin, D.B. Effect of Aluminum Adjuvant and Preservatives on Structural Integrity and Physicochemical Stability Profiles of Three Recombinant Subunit Rotavirus Vaccine Antigens. J. Pharm. Sci. 2020, 109, 476–487. [Google Scholar] [CrossRef]
- Kaur, K.; Xiong, J.; Sawant, N.; Agarwal, S.; Hickey, J.M.; Holland, D.A.; Mukhopadhyay, T.K.; Brady, J.R.; Dalvie, N.C.; Tracey, M.K.; et al. Mechanism of Thimerosal-Induced Structural Destabilization of a Recombinant Rotavirus P[4] Protein Antigen Formulated as a Multi-Dose Vaccine. J. Pharm. Sci. 2021, 110, 1054–1066. [Google Scholar] [CrossRef]
- Maman, K.; Zöllner, Y.; Greco, D.; Duru, G.; Sendyona, S.; Remy, V. The Value of Childhood Combination Vaccines: From Beliefs to Evidence. Hum. Vaccin. Immunother. 2015, 11, 2132–2141. [Google Scholar] [CrossRef] [PubMed]
- Agnolon, V.; Bruno, C.; Galletti, B.; Mori, E.; Ugozzoli, M.; Pergola, C.; O’hagan, D.T.; Baudner, B.C. Multiplex Immunoassay for in Vitro Characterization of Acellular Pertussis Antigens in Combination Vaccines. Vaccine 2016, 34, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Kraan, H.; Have, R.T.; van der Maas, L.; Kersten, G.; Amorij, J.-P. Incompatibility of Lyophilized Inactivated Polio Vaccine with Liquid Pentavalent Whole-Cell-Pertussis-Containing Vaccine. Vaccine 2016, 34, 4572–4578. [Google Scholar] [CrossRef] [PubMed]
- WHO. Thimerosal. Available online: https://www.who.int/teams/health-product-policy-and-standards/standards-and-specifications/vaccines-quality/thiomersal#:~:text=In%20some%20production%20processes%20thiomersal,of%20whole%20cell%20pertussis%20vaccine (accessed on 13 September 2023).
- Sawant, N.; Joshi, S.B.; Weis, D.D.; Volkin, D.B. Interaction of Aluminum-Adjuvanted Recombinant P[4] Protein Antigen with Preservatives: Storage Stability and Backbone Flexibility Studies. J. Pharm. Sci. 2022, 111, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Dounighi, N.M.; Razzaghi-Abyane, M.; Nofeli, M.; Zolfagharian, H.; Shahcheraghi, F. Study on Toxicity Reduction and Potency Induction in Whole-Cell Pertussis Vaccine by Developing a New Optimal Inactivation Condition Processed on Bordetella Pertussis. Jundishapur J. Microbiol. 2016, 9, e34153. [Google Scholar]
- Biotech Panacea. Easysix Package Insert. Available online: https://media.panaceabiotec.com/documents/2019/7/24/Easysix-PMPIS05903.pdf (accessed on 12 September 2023).
- Hexasiil. Serum Institute of India. Available online: https://www.seruminstitute.com/product_hexasiil.php (accessed on 14 December 2023).
- Shan6 Package Insert. Sanofi Healthcare. Available online: https://cdsco.gov.in/opencms/resources/UploadCDSCOWeb/2018/UploadSmPC/SmPC%20of%20%20Shan6%20Multi-dose%20of%20%20by%20Ms%20Sanofi%20Healthcare%20India%20Pvt%20Ltd%20%20%20.pdf (accessed on 14 December 2023).
- Sharma, H.; Lalwani, S.; Parekh, S.; Pujari, P.; Shewale, S.; Palkar, S.; Hanumante, N.; Gokhale, S.; Ks, J.; Kumar, R.; et al. A Phase I, Open Label, Clinical Study to Assess the Safety and Immunogenicity of Indigenously Developed Liquid (Dtwp-Hepb-Ipv-Hib) Hexavalent Combination Vaccine in Healthy Toddlers Aged 16–24 Months. Hum. Vaccines Immunother. 2022, 18, 2146435. [Google Scholar] [CrossRef]
- Rakesh, K.; Sharma, I.J.; Shitole, A.V.; Doddapaneni, M.; Sharma, H.J. An Immunogenic Composition Having Improved Stability, Enhanced Immunogenicity and Reduced Reactogenicity and Process for Preparation Thereof. Patent WO2019016654, 24 January 2019. [Google Scholar]
- Young, C.S.; Ae, N.Y.; Ji, K.E. Combination Vaccine Composition for Multiple-Dosage. Patent WO2017048038A1, 23 March 2017. [Google Scholar]
- ClinicalTrials.gov. Study of Dtwp-Hepb-Hib-Ipv (Shan6™) Vaccine Administered Concomitantly with Routine Pediatric Vaccines to Healthy Infants and Toddlers in Thailand. Available online: https://classic.clinicaltrials.gov/ct2/show/study/NCT04429295 (accessed on 12 September 2023).
- Hausdorff, W.P.; Price, J.; Debellut, F.; Mooney, J.; Torkelson, A.A.; Giorgadze, K.; Pecenka, C. Does Anybody Want an Injectable Rotavirus Vaccine, and Why? Understanding the Public Health Value Proposition of Next-Generation Rotavirus Vaccines. Vaccines 2022, 10, 149. [Google Scholar] [CrossRef]
- Orsi, A.; Azzari, C.; Bozzola, E.; Chiamenti, G.; Chirico, G.; Esposito, S.; Francia, F.; Lopalco, P.; Prato, R.; Russo, R.; et al. Hexavalent Vaccines: Characteristics of Available Products and Practical Considerations from a Panel of Italian Experts. J. Prev. Med. Hyg. 2018, 59, E107–E119. [Google Scholar]
- WHO. Requirements for Diphtheria, Tetanus, Pertussis and Combined Vaccines. Available online: https://cdn.who.int/media/docs/default-source/biologicals/vaccine-quality/requirements-for-diphteria-tetanus-pertussis-and-combined-vaccinesd5900f3c-b63e-4f7e-b7df-fcef6a149b39.pdf?sfvrsn=6da899bc_1&download=true (accessed on 14 December 2023).
- Xing, D.; Markey, K.; Das, R.G.; Feavers, I. Whole-Cell Pertussis Vaccine Potency Assays: The Kendrick Test and Alternative Assays. Expert. Rev. Vaccines 2014, 13, 1175–1182. [Google Scholar] [CrossRef]
- Suresh, K.; Mahendra, P.S.; Vijay, K.B.; Ramendra, P.P. Quality Control of Vaccines-a Journey from Classical Approach to 3rs. Microbiol. Curr. Res. 2018, 2, 45–61. [Google Scholar]
- Iwaki, M.; Kenri, T.; Senoh, M. An Elisa System for Tetanus Toxoid Potency Tests: An Alternative to Lethal Challenge. Biologicals 2023, 82, 101681. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Fujii, Y.; Someya, Y. Effects of the Thermal Denaturation of Sabin-Derived Inactivated Polio Vaccines on the D-Antigenicity and the Immunogenicity in Rats. Vaccine 2020, 38, 3295–3299. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Hickey, J.M.; Sahni, N.; Toth, R.T.; Robertson, G.A.; Sitrin, R.; Cryz, S.; Joshi, S.B.; Volkin, D.B. Recombinant Subunit Rotavirus Trivalent Vaccine Candidate: Physicochemical Comparisons and Stability Evaluations of Three Protein Antigens. J. Pharm. Sci. 2020, 109, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Duprez, J.; Kalbfleisch, K.; Deshmukh, S.; Payne, J.; Haer, M.; Williams, W.; Durowoju, I.; Kirkitadze, M. Structure and Compositional Analysis of Aluminum Oxyhydroxide Adsorbed Pertussis Vaccine. Comput. Struct. Biotechnol. J. 2021, 19, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Kalbfleisch, K.; Deshmukh, S.; Mei, C.; Ore, M.; Williams, W.; Durowoju, I.; Duprez, J.; Morin, S.; Carpick, B.; Kirkitadze, M. Identity, Structure and Compositional Analysis of Aluminum Phosphate Adsorbed Pediatric Quadrivalent and Pentavalent Vaccines. Comput. Struct. Biotechnol. J. 2019, 17, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Crisel, R.M.; Baker, R.S.; Dorman, D.E. Capsular Polymer of Haemophilus Influenzae, Type B. J. Biol. Chem. 1975, 250, 4926–4930. [Google Scholar] [CrossRef] [PubMed]
- HogenEsch, H.; O’Hagan, D.T.; Fox, C.B. Optimizing the Utilization of Aluminum Adjuvants in Vaccines: You Might Just Get What You Want. NPJ Vaccines 2018, 51, 51. [Google Scholar] [CrossRef]
- US-FDA. Pentacel Packahe Insert. Available online: https://www.fda.gov/media/74385/download (accessed on 13 September 2023).
- WHO. Eupenta Package Insert. Available online: https://extranet.who.int/pqweb/content/eupenta (accessed on 13 September 2023).
- Chem, L.G. A Phase Iv Study to Assess the Safety of Eupentatm Inj. Available online: https://clinicaltrials.gov/study/NCT04056728 (accessed on 14 December 2023).
- Meyer, B.K.; Ni, A.; Hu, B.; Shi, L. Antimicrobial Preservative Use in Parenteral Products: Past and Present. J. Pharm. Sci. 2007, 96, 3155–3167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aluminum-Salt-Adjuvanted Antigen | Assay | Capture Antibody | Capture Ab Binds | Reference | |
|---|---|---|---|---|---|
| NRRV antigens | NRRV P[8] | Competition ELISA | mAb 7H7 (Precision Antibody) | Linear epitope of NRRV P[8] | [26,27] |
| NRRV P[6] | mAb 3G11 (Precision Antibody) | Linear epitope of NRRV P[6] | |||
| NRRV P[4] | mAb 13A1 (Precision Antibody) | Conformational epitope NRRV P[4] | |||
| Pentavalent antigens | Diphtheria (D) | mAb 10/130 (NIBSC) | Conformational epitopes on diphtheria toxoid | [28,29] | |
| Tetanus (T) | mAb NIBSC 10/134 (NIBSC) | Conformational epitopes on tetanus toxin | |||
| PRP-T conjugate (Hib) | HIB12-S (Alpha Diagnostics) | Hib PRP region | [30] | ||
| Hepatitis B surface antigen (HepB) | mAb 4940-1404 surface sntigen (Bio-Rad) | Conformational epitopes on Hep B surface antigen | [31] | ||
| Whole-cell inactivated pertussis (wP) | Mouse Immunogenicity | N/A | N/A | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, P.; Holland, D.A.; Secrist, K.; Taskar, P.; Dotson, B.; Saleh-Birdjandi, S.; Adewunmi, Y.; Doering, J.; Mantis, N.J.; Volkin, D.B.; et al. Evaluating the Compatibility of New Recombinant Protein Antigens (Trivalent NRRV) with a Mock Pentavalent Combination Vaccine Containing Whole-Cell Pertussis: Analytical and Formulation Challenges. Vaccines 2024, 12, 609. https://doi.org/10.3390/vaccines12060609
Kumar P, Holland DA, Secrist K, Taskar P, Dotson B, Saleh-Birdjandi S, Adewunmi Y, Doering J, Mantis NJ, Volkin DB, et al. Evaluating the Compatibility of New Recombinant Protein Antigens (Trivalent NRRV) with a Mock Pentavalent Combination Vaccine Containing Whole-Cell Pertussis: Analytical and Formulation Challenges. Vaccines. 2024; 12(6):609. https://doi.org/10.3390/vaccines12060609
Chicago/Turabian StyleKumar, Prashant, David A. Holland, Kathryn Secrist, Poorva Taskar, Brandy Dotson, Soraia Saleh-Birdjandi, Yetunde Adewunmi, Jennifer Doering, Nicholas J. Mantis, David B. Volkin, and et al. 2024. "Evaluating the Compatibility of New Recombinant Protein Antigens (Trivalent NRRV) with a Mock Pentavalent Combination Vaccine Containing Whole-Cell Pertussis: Analytical and Formulation Challenges" Vaccines 12, no. 6: 609. https://doi.org/10.3390/vaccines12060609