
FlagT4G Vaccine Prevents Transplacental Transmission of Highly Virulent Classical Swine Fever Virus after Single Vaccination in Pregnant Sows

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Experimental Design
2.3. Detection of Antibody Responses by ELISA and Virus Neutralization Tests
2.4. CSFV RNA Detection
2.5. Determination of IFN-α Levels in Serum by ELISA Test
2.6. CSFV Isolation in Tonsils Samples
3. Results
3.1. FlagT4G Vaccine Protects Pregnant Sows from Clinical Signs after Challenge with a Highly Virulent CSFV
3.2. FlagT4G Elicited Strong CSFV-Specific Antibody in Vaccinated Pregnant Sows
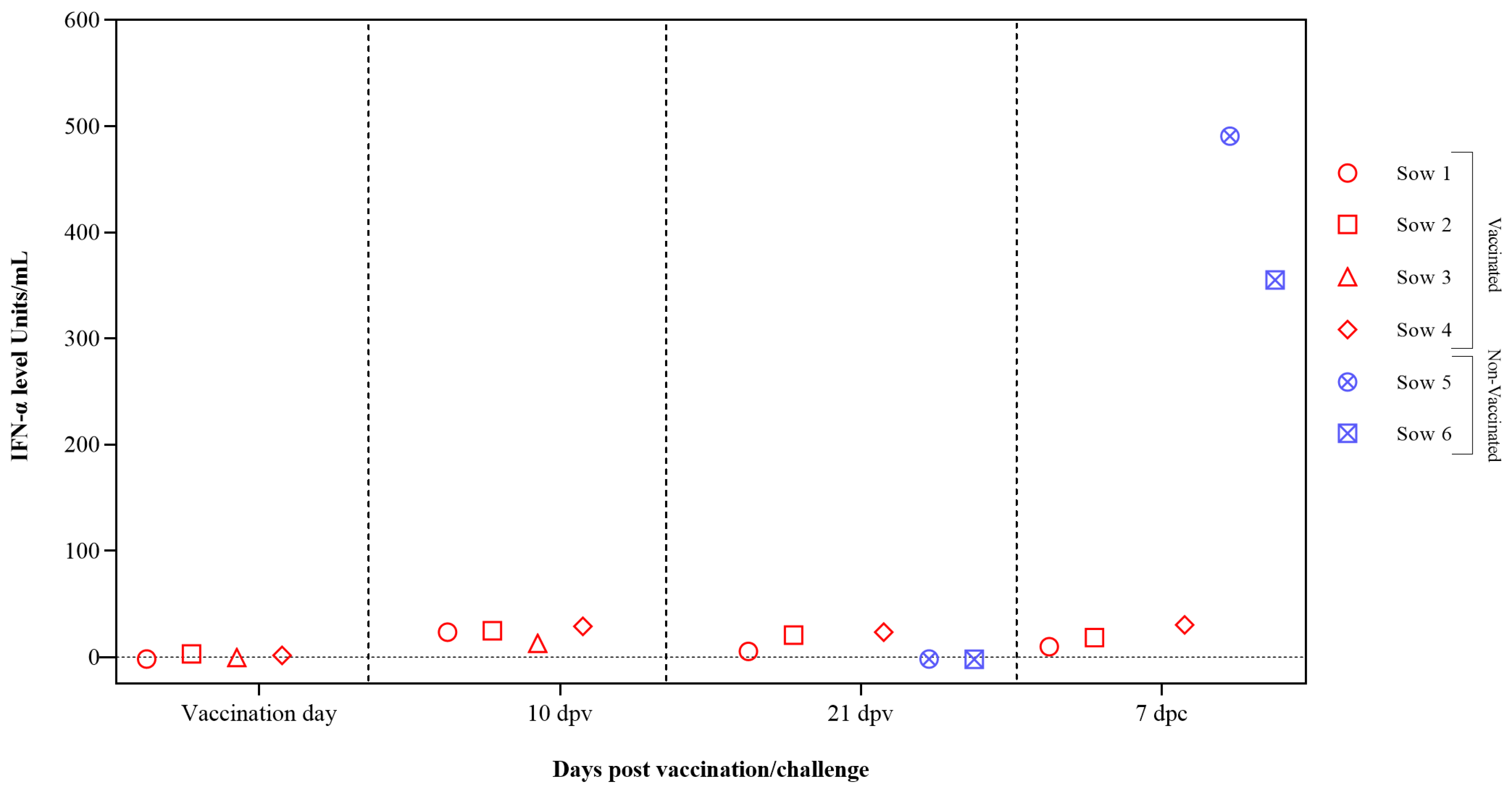
3.3. The CSFV-Induced Interferon Alpha-Exacerbated Response Is Prevented by FlagT4G Vaccine
3.4. FlagT4G Vaccine Avoids Systemic and Tissue Replication of the Highly Virulent Challenge Virus
3.5. FlagT4G Vaccine Afforded Total Protection from CSFV Highly Virulent Challenge in the Foetuses from Vaccinated Sows
3.6. FlagT4G Vaccine Colonizes the Foetuses from Vaccinated Sows with a High CSFV RNA Load at Systemic and Tissue Levels
3.7. CSFV Antibody and IFN-α Responses Were Not Found in the Foetuses from Vaccinated Sows
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Tautz, N.; Tews, B.A.; Meyers, G. The Molecular Biology of Pestiviruses. Adv. Virus Res. 2015, 93, 47–160. [Google Scholar] [PubMed]
- Rümenapf, T.; Unger, G.; Strauss, J.H.; Thiel, H.J. Processing of the envelope glycoproteins of pestiviruses. J. Virol. 1993, 67, 3288–3294. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical Signs and Epidemiology of Classical Swine Fever: A Re-view of New Knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Bohórquez, J.A.; Muñoz-González, S.; Pérez-Simó, M.; Muñoz, I.; Rosell, R.; Coronado, L.; Domingo, M.; Ganges, L. Foetal immune response activation and high replication rate during generation of classical swine fever congenital infection. Pathogens 2020, 9, 285. [Google Scholar] [CrossRef]
- Van Oirschot, J.; Terpstra, C. A congenital persistent swine fever infection. I. Clinical and virological observations. Vet. Microbiol. 1977, 2, 121–132. [Google Scholar] [CrossRef]
- Van Oirschot, J. Experimental production of congenital persistent swine fever infections: II. Effect on functions of the immune system. Vet. Microbiol. 1979, 4, 133–147. [Google Scholar] [CrossRef]
- Coronado, L.; Bohórquez, J.A.; Muñoz-González, S.; Perez, L.J.; Rosell, R.; Fonseca, O.; Delgado, L.; Perera, C.L.; Frías, M.T.; Ganges, L. Investigation of chronic and persistent classical swine fever infections under field conditions and their impact on vaccine efficacy. BMC Vet. Res. 2019, 15, 247. [Google Scholar] [CrossRef] [PubMed]
- Liess, B. Persistent infections of hog cholera: A review. Prev. Vet. Med. 1984, 2, 109–113. [Google Scholar] [CrossRef]
- Henke, J.; Carlson, J.; Zani, L.; Leidenberger, S.; Schwaiger, T.; Schlottau, K.; Teifke, J.P.; Schröder, C.; Beer, M.; Blome, S. Protection against transplacental transmission of moderately virulent classical swine fever virus using live marker vaccine “CP7_E2alf”. Vaccine 2018, 36, 4181–4187. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (WOAH). Chapter 3.9.3: Classical Swine Fever Virus (infection with Classical Swine Fever Virus). In WOAH Terrestrial Manual; WOAH: Paris, France, 2022. [Google Scholar]
- Meyer, D.; Loeffen, W.; Postel, A.; Fritsche, S.; Becher, P. Reduced specificity of E rns antibody ELISAs for samples from piglets with maternally derived antibodies induced by vaccination of sows with classical swine fever marker vaccine CP7_E2alf. Transbound. Emerg. Dis. 2018, 65, e505–e508. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Pedroso, M.; Sordo-Puga, Y.; Sosa-Teste, I.; Rodriguez-Molto, M.P.; Naranjo-Valdes, P.; Sardina-Gonzalez, T.; Santana-Rodriguez, E.; Montero-Espinosa, C.; Frias-Laporeaux, M.T.; Fuentes-Rodriguez, Y.; et al. Novel chimeric E2CD154 subunit vaccine is safe and confers long lasting protection against classi-cal swine fever virus. Vet. Immunol. Immunopathol. 2021, 234, 110222. [Google Scholar] [CrossRef]
- Muñoz-González, S.; Sordo, Y.; Pérez-Simó, M.; Suarez, M.; Canturri, A.; Rodriguez, M.P.; Frías-Lepoureau, M.T.; Domingo, M.; Estrada, M.P.; Ganges, L. Corrigendum to “Efficacy of E2 glycoprotein fused to porcine CD154 as a novel chimeric subunit vaccine to prevent classical swine fever virus vertical transmission in pregnant sows”. Vet. Microbiol. 2018, 213, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Holinka, L.; Fernandez-Sainz, I.; Sanford, B.; O’Donnell, V.; Gladue, D.; Carlson, J.; Lu, Z.; Risatti, G.; Borca, M. Development of an improved live attenuated antigenic marker CSF vaccine strain candidate with an increased genetic stability. Virology 2014, 471–473, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Bohórquez, J.A.; Defaus, S.; Rosell, R.; Pérez-Simó, M.; Alberch, M.; Gladue, D.P.; Borca, M.V.; Andreu, D.; Ganges, L. Development of a dendrimeric peptide-based approach for the differentiation of animals vaccinated with flagt4g against classical swine fever from infected pigs. Viruses 2021, 13, 1980. [Google Scholar] [CrossRef] [PubMed]
- Bohórquez, J.A.; Wang, M.; Díaz, I.; Alberch, M.; Pérez-Simó, M.; Rosell, R.; Gladue, D.P.; Borca, M.V.; Ganges, L. The FlagT4G Vaccine Confers a Strong and Regulated Immunity and Early Virological Protection against Classical Swine Fever. Viruses 2022, 14, 1954. [Google Scholar] [CrossRef] [PubMed]
- Holinka, L.G.; O’donnell, V.; Risatti, G.R.; Azzinaro, P.; Arzt, J.; Stenfeldt, C.; Velazquez-Salinas, L.; Carlson, J.; Gladue, D.P.; Borca, M.V. Early protection events in swine immunized with an experimental live attenuated classical swine fever marker vaccine, FlagT4G. PLoS ONE 2017, 12, e0177433. [Google Scholar] [CrossRef] [PubMed]
- Vannier, P.; Plateau, E.; Tillon, J.P. Congenital tremor in pigs farrowed from sows given hog cholera virus during pregnancy. Am. J. Vet. Res. 1981, 42, 135–137. [Google Scholar] [PubMed]
- Kaden, V.; Lange, E.; Steyer, H.; Lange, B.; Klopfleisch, R.; Teifke, J.; Bruer, W. Classical swine fever virus strain “C” protects the offspring by oral immunisation of pregnant sows. Vet. Microbiol. 2008, 130, 20–27. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Boonstra, J.; Bloemraad, M.; Van Zaane, D. Production of monoclonal antibodies against swine fever virus and their use in laboratory diagnosis. Vet. Microbiol. 1986, 12, 101–108. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Terpstra, C.; Bloemraad, M.; Gielkens, A.L.J. The neutralizing peroxidase-linked assay for detection of antibody against swine fever virus. Vet. Microbiol. 1984, 9, 113–120. [Google Scholar] [CrossRef]
- Terpstra, C.; Wensvoort, G. The protective value of vaccine-induced neutralising antibody titres in swine fever. Vet. Microbiol. 1988, 16, 123–128. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Muñoz-González, S.; Pérez-Simó, M.; Colom-Cadena, A.; Cabezón, O.; Bohórquez, J.A.; Rosell, R.; Pérez, L.J.; Marco, I.; Lavín, S.; Domingo, M.; et al. Classical swine fever virus vs. Classical swine fever virus: The superinfection exclusion phenome-non in experimentally infected wild boar. PLoS ONE 2016, 11, e0149469. [Google Scholar] [CrossRef]
- Wang, M.; Bohórquez, J.A.; Muñoz-González, S.; Gerber, M.; Alberch, M.; Pérez-Simó, M.; Abad, X.; Liniger, M.; Ruggli, N.; Ganges, L. Removal of the E rns RNase Activity and of the 3′ Untranslated Region Polyuridine Insertion in a Low-Virulence Classical Swine Fever Virus Triggers a Cytokine Storm and Lethal Disease. J. Virol. 2022, 96, e0043822. [Google Scholar] [CrossRef]
- Lowings, P.; Ibata, G.; Needham, J.; Paton, D. Classical swine fever virus diversity and evolution. J. Gen. Virol. 1996, 77, 1311–1321. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids 1999, 41, 95–98. [Google Scholar]
- de Arce, H.D.; Artursson, K.; L’Haridon, R.; Perers, A.; La Bonnardiere, C.; Alm, G. A sensitive immunoassay for porcine interferon-α. Vet. Immunol. Immunopathol. 1992, 30, 319–327. [Google Scholar] [CrossRef]
- Guzylack-Piriou, L.; Balmelli, C.; McCullough, K.C.; Summerfield, A. Type-A CpG oligonucleotides activate exclusively porcine natural interferon-producing cells to secrete interferon-α, tumour necrosis factor-α and interleukin-12. Immunology 2004, 112, 28–37. [Google Scholar] [CrossRef]
- Kaden, V.; Lange, E.; Riebe, R.; Lange, B. Classical Swine Fever Virus Strain ‘C’. How Long is it Detectable After Oral Vac-cination? J. Vet. Med. Ser. B 2004, 51, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, A.; Ruggli, N. Immune Responses Against Classical Swine Fever Virus: Between Ignorance and Lunacy. Front. Vet. Sci. 2015, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Kaden, V.; Lange, E.; Polster, U.; Klopfleisch, R.; Teifke, J.P. Studies on the Virulence of Two Field Isolates of the Classical Swine Fever Virus Genotype 2.3 Rostock. in Wild Boars of Different Age Groups. J. Vet. Med. Ser. B 2004, 51, 202–208. [Google Scholar] [CrossRef]
- Maciag, S.; Volpato, F.; Bombassaro, G.; Forner, R.; Oliveira, K.P.; Bovolato, A.L.C.; Lopes, L.; Bastos, A.P. Effects of freezing storage on the stability of maternal cellular and humoral immune components in porcine colostrum. Vet. Immunol. Immunopathol. 2022, 254, 110520. [Google Scholar] [CrossRef]
- Folimonova, S.Y. Superinfection Exclusion Is an Active Virus-Controlled Function That Requires a Specific Viral Protein. J. Virol. 2012, 86, 5554–5561. [Google Scholar] [CrossRef] [PubMed]
- Soller, A.; Epstein, H. Biochemical and immunological aspects of the exclusion of lambda by superinfection with T4. Virology 1965, 26, 715–726. [Google Scholar] [CrossRef]
- Ramírez, S.; Pérez-Del-Pulgar, S.; Carrión, J.A.; Coto-Llerena, M.; Mensa, L.; Dragun, J.; García-Valdecasas, J.C.; Navasa, M.; Forns, X. Hepatitis C virus superinfection of liver grafts: A detailed analysis of early exclusion of non-dominant virus strains. J. Gen. Virol. 2010, 91, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Rao, J.; Zhang, J.; Ye, J.; Cao, S.; Chen, H.; Song, Y. Japanese encephalitis virus inhibits superinfection of Zika virus in cells by the NS2B protein. J. Virol. 2024, 98, e0185923. [Google Scholar] [CrossRef]
- Cwick, J.P.; Owen, J.E.; Kochetkova, I.; Hain, K.S.; Van Horssen, N.; Taylor, M.P. Superinfection Exclusion of Alphaherpesviruses Interferes with Virion Trafficking. Microbiol. Spectr. 2022, 10, e0068422. [Google Scholar] [CrossRef]
- Burkard, T.; Proske, N.; Resner, K.; Collignon, L.; Knegendorf, L.; Friesland, M.; Verhoye, L.; Sayed, I.M.; Brüggemann, Y.; Nocke, M.K.; et al. Viral Interference of Hepatitis C and E Virus Replication in Novel Experimental Co-Infection Systems. Cells 2022, 11, 927. [Google Scholar] [CrossRef]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex Flavivirus and West Nile Virus Mosquito Coinfection and Positive Ecological Association in Chicago, United States. Vector-Borne Zoonotic Dis. 2011, 11, 1099–1105. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Tscherne, D.M.; Yun, S.-I.; Frolov, I.; Rice, C.M. Dual Mechanisms of Pestiviral Superinfection Exclusion at Entry and RNA Replication. J. Virol. 2005, 79, 3231–3242. [Google Scholar] [CrossRef]
- Walters, K.-A.; Joyce, M.A.; Addison, W.R.; Fischer, K.P.; Tyrrell, D.L.J. Superinfection Exclusion in Duck Hepatitis B Virus Infection Is Mediated by the Large Surface Antigen. J. Virol. 2004, 78, 7925–7937. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FlagT4G Vaccine (Genotype 1.2) | CSFV Margarita Strain (Genotype 1.4) | CSFV Diepholz Strain (Genotype 2.3) | CSFV Alfort/187 Strain (Genotype 1.1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sow ID | 21 dpv | 7 dpc | 14 dpc | 21 dpv | 7 dpc | 14 dpc | 21 dpv | 7 dpc | 14 dpc | 21 dpv | 7 dpc | 14 dpc | |
| FlagT4G vaccinated | 1 | 1280 | 2560 | 1260 | 40 | 640 | 640 | 320 | 640 | 160 | 160 | 320 | 640 |
| 2 | 320 | 1280 | 1280 | 40 | 320 | 640 | 80 | 1280 | 5120 | 20 | 80 | 160 | |
| 3 † |  | ||||||||||||
| 4 | 640 | 320 | 1260 | 40 | 160 | 160 | 160 | 320 | 128 | 40 | 80 | 160 | |
| Non-vaccinated | 5 | (−) | (−) | 80 * | (−) | (−) | 160 * | (−) | (−) | 160 * | (−) | (−) | 10 * |
| 6 | (−) | (−) | (−) | (−) | (−) | (−) | (−) | (−) | (−) | (−) | (−) | (−) | |
 = no sample; (−) = negative sample; † = sow euthanized at 9 dpv; * = sample taken at 12 dpc.
= no sample; (−) = negative sample; † = sow euthanized at 9 dpv; * = sample taken at 12 dpc.Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coronado, L.; Muñoz-Aguilera, A.; Cantero, G.; Martínez, P.; Alberch, M.; Rosell, R.; Gladue, D.P.; Borca, M.V.; Ganges, L. FlagT4G Vaccine Prevents Transplacental Transmission of Highly Virulent Classical Swine Fever Virus after Single Vaccination in Pregnant Sows. Vaccines 2024, 12, 832. https://doi.org/10.3390/vaccines12080832
Coronado L, Muñoz-Aguilera A, Cantero G, Martínez P, Alberch M, Rosell R, Gladue DP, Borca MV, Ganges L. FlagT4G Vaccine Prevents Transplacental Transmission of Highly Virulent Classical Swine Fever Virus after Single Vaccination in Pregnant Sows. Vaccines. 2024; 12(8):832. https://doi.org/10.3390/vaccines12080832
Chicago/Turabian StyleCoronado, Liani, Adriana Muñoz-Aguilera, Guillermo Cantero, Patricia Martínez, Mònica Alberch, Rosa Rosell, Douglas P. Gladue, Manuel V. Borca, and Llilianne Ganges. 2024. "FlagT4G Vaccine Prevents Transplacental Transmission of Highly Virulent Classical Swine Fever Virus after Single Vaccination in Pregnant Sows" Vaccines 12, no. 8: 832. https://doi.org/10.3390/vaccines12080832