
Prime-Boost Vaccination Based on Nanospheres and MVA Encoding the Nucleoprotein of Crimean-Congo Hemorrhagic Fever Virus Elicits Broad Immune Responses

 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus, Bacteria, and Cells
2.2. Construction of Recombinant Baculovirus to Express NP of CCHFV
2.3. Production and Purification of muNS-Mi Nanospheres Tagged to NP
2.4. Generation of Recombinant MVA-NP
2.5. Purification of the NP from MVA-NP by CsCl Gradient
2.6. Immunization of Mice
2.7. Enzyme-Linked Immunosorbent Assays
2.8. Selection of CCHFV NP Peptides
2.9. Flow Cytometric Analysis
2.10. Immunofluorescence Assays
2.11. Measurement of Cytokines by Luminex
2.12. Statistical Analyses
3. Results
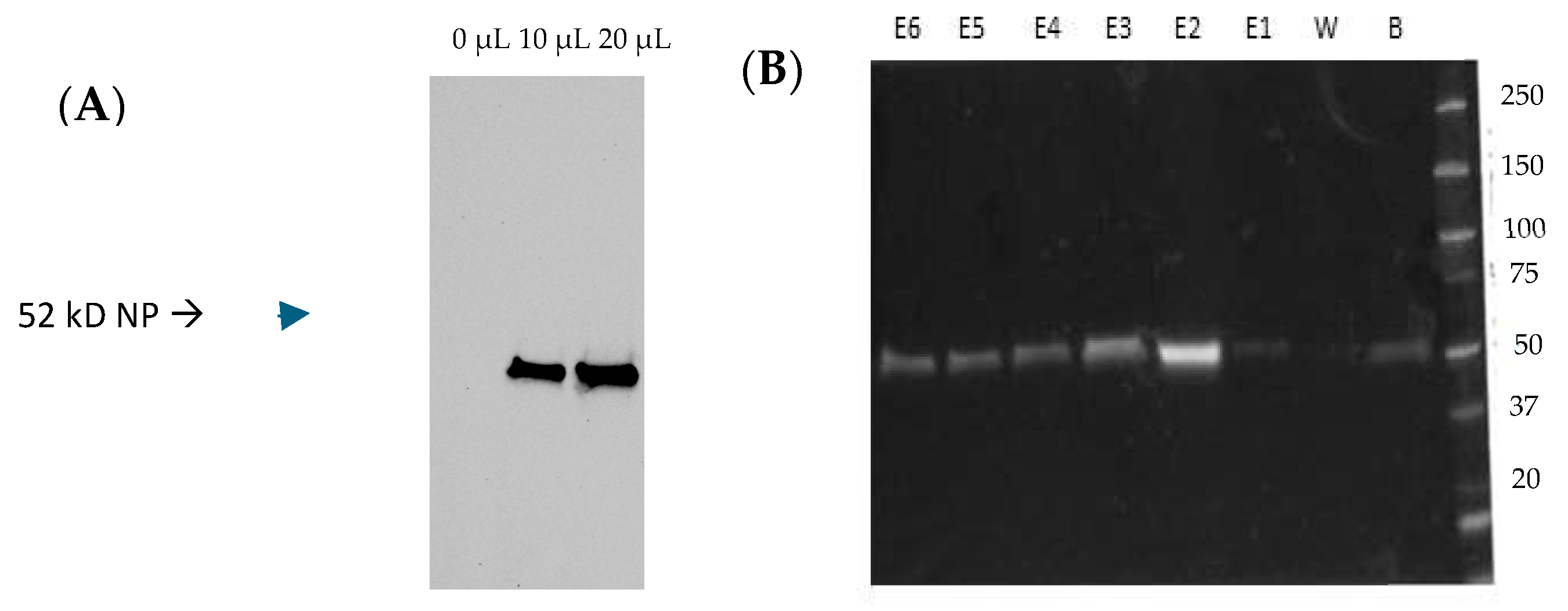
3.1. Expression of the IC-Tagged CCHFV NP and Association with muNS-Mi NS
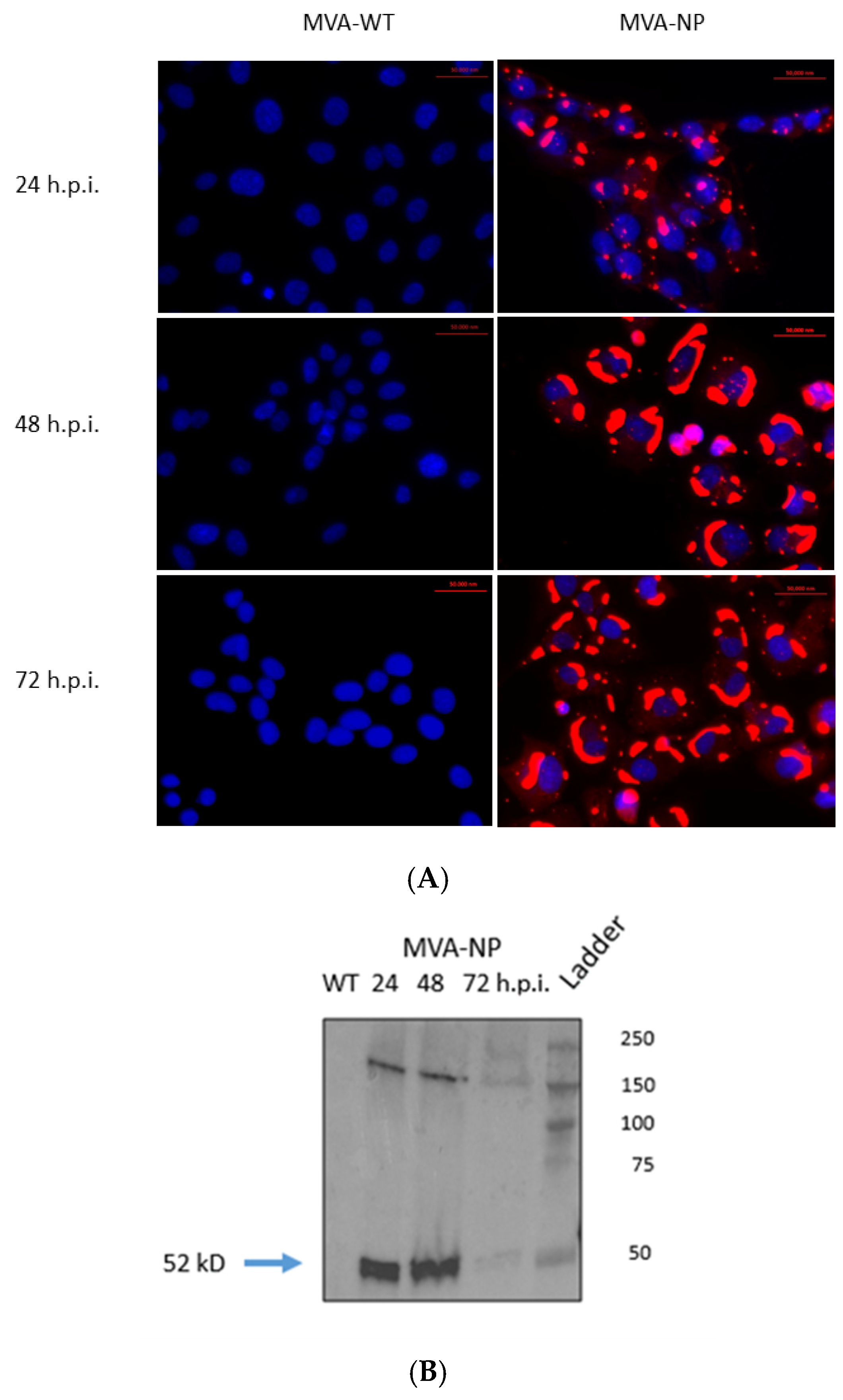
3.2. In Vitro Expression of CCHFV NP by Recombinant MVA-NP
3.3. Induction of Specific Immune Responses After Vaccination with NS-NP and MVA-NP Expressing CCHFV NP
3.3.1. Detection of Specific Humoral Responses After Vaccination
3.3.2. Cellular Immune Responses Induced After NP Vaccination
3.3.3. Up-Regulation of Cytokines in the Vaccinated Animals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swanepoel, R.; Shepherd, A.J.; Leman, P.A.; Shepherd, S.P.; McGillivray, G.M.; Erasmus, M.J.; Searle, L.A.; Gill, D.E. Epidemiologic and Clinical Features of Crimean-Congo Hemorrhagic Fever in Southern Africa. Am. J. Trop. Med. Hyg. 1987, 36, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Pigott, D.M.; Golding, N.; Duda, K.A.; Brownstein, J.S.; Weiss, D.J.; Gibson, H.; Robinson, T.P.; Gilbert, M.; William Wint, G.R.; et al. The Global Distribution of Crimean-Congo Hemorrhagic Fever. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 503–513. [Google Scholar] [CrossRef]
- Al-Abri, S.S.; Hewson, R.; Al-Kindi, H.; Al-Abaidani, I.; Al-Jardani, A.; Al-Maani, A.; Almahrouqi, S.; Atkinson, B.; Al-Wahaibi, A.; Al-Rawahi, B.; et al. Clinical and Molecular Epidemiology of Crimean-Congo Hemorrhagic Fever in Oman. PLoS Negl. Trop. Dis. 2019, 13, e0007100. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Palomar, A.M.; Santibáñez, P.; Sánchez, N.; Habela, M.A.; Portillo, A.; Romero, L.; Oteo, J.A. Crimean-Congo Hemorrhagic Fever Virus in Ticks, Southwestern Europe, 2010. Emerg. Infect. Dis. 2012, 18, 179–180. [Google Scholar] [CrossRef]
- Lorenzo Juanes, H.M.; Carbonell, C.; Sendra, B.F.; López-Bernus, A.; Bahamonde, A.; Orfao, A.; Lista, C.V.; Ledesma, M.S.; Negredo, A.I.; Rodríguez-Alonso, B.; et al. Crimean-Congo Hemorrhagic Fever, Spain, 2013–2021. Emerg. Infect. Dis. 2023, 29, 252–259. [Google Scholar] [CrossRef]
- Negredo, A.; de la Calle-Prieto, F.; Palencia-Herrejón, E.; Mora-Rillo, M.; Astray-Mochales, J.; Sánchez-Seco, M.P.; Bermejo Lopez, E.; Menárguez, J.; Fernández-Cruz, A.; Sánchez-Artola, B.; et al. Autochthonous Crimean-Congo Hemorrhagic Fever in Spain. N. Engl. J. Med. 2017, 377, 154–161. [Google Scholar] [CrossRef]
- Rasikh, A.S.; Aram, M.M.; Noory, A.T. Clinical and Epidemiological Characteristics of 30 Fatal Cases of Crimean-Congo Hemorrhagic Fever in Kabul, Afghanistan: A Retrospective Observational Study. Infect. Drug Resist. 2023, 16, 3469–3476. [Google Scholar] [CrossRef] [PubMed]
- Atwan, Z.; Alhilfi, R.; Mousa, A.K.; Rawaf, S.; Torre, J.D.L.; Hashim, A.R.; Sharquie, I.K.; Khaleel, H.; Tabche, C. Alarming Update on Incidence of Crimean-Congo Hemorrhagic Fever in Iraq in 2023. IJID Reg. 2023, 10, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Lasecka, L.; Baron, M.D. The Molecular Biology of Nairoviruses, an Emerging Group of Tick-Borne Arboviruses. Arch. Virol. 2014, 159, 1249–1265. [Google Scholar] [CrossRef]
- Adam, I.A.; Mahmoud, M.A.M.; Aradaib, I.E. A Seroepidemiological Survey of Crimean Congo Hemorrhagic Fever among Cattle in North Kordufan State, Sudan. Virol. J. 2013, 10, 178. [Google Scholar] [CrossRef]
- Dzikwi-Emennaa, A.A.; Meseko, C.; Emennaa, P.; Adeyinka, A.J.; Adamu, A.M.; Adegboye, O.A. Detection of Crimean-Congo Hemorrhagic Fever Virus Antibodies in Cattle in Plateau State, Nigeria. Viruses 2022, 14, 2618. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Alkhovsky, S.V.; Avšič-Županc, T.; Bergeron, É.; Burt, F.; Ergünay, K.; Garrison, A.R.; Marklewitz, M.; Mirazimi, A.; Papa, A.; et al. ICTV Virus Taxonomy Profile: Nairoviridae 2024. J. Gen. Virol. 2024, 105, 001974. [Google Scholar] [CrossRef]
- Zivcec, M.; Scholte, F.E.M.; Spiropoulou, C.F.; Spengler, J.R.; Bergeron, É. Molecular Insights into Crimean-Congo Hemorrhagic Fever Virus. Viruses 2016, 8, 106. [Google Scholar] [CrossRef]
- Barnwal, B.; Karlberg, H.; Mirazimi, A.; Tan, Y.-J. The Non-Structural Protein of Crimean-Congo Hemorrhagic Fever Virus Disrupts the Mitochondrial Membrane Potential and Induces Apoptosis. J. Biol. Chem. 2016, 291, 582–592. [Google Scholar] [CrossRef]
- Hewson, R.; Gmyl, A.; Gmyl, L.; Smirnova, S.E.; Karganova, G.; Jamil, B.; Hasan, R.; Chamberlain, J.; Clegg, C. Evidence of Segment Reassortment in Crimean-Congo Haemorrhagic Fever Virus. J. Gen. Virol. 2004, 85, 3059–3070. [Google Scholar] [CrossRef] [PubMed]
- Fillâtre, P.; Revest, M.; Tattevin, P. Crimean-Congo Hemorrhagic Fever: An Update. Med. Mal. Infect. 2019, 49, 574–585. [Google Scholar] [CrossRef]
- Sana, M.; Javed, A.; Babar Jamal, S.; Junaid, M.; Faheem, M. Development of Multivalent Vaccine Targeting M Segment of Crimean Congo Hemorrhagic Fever Virus (CCHFV) Using Immunoinformatic Approaches. Saudi J. Biol. Sci. 2022, 29, 2372–2388. [Google Scholar] [CrossRef]
- Garrison, A.R.; Shoemaker, C.J.; Golden, J.W.; Fitzpatrick, C.J.; Suschak, J.J.; Richards, M.J.; Badger, C.V.; Six, C.M.; Martin, J.D.; Hannaman, D.; et al. A DNA Vaccine for Crimean-Congo Hemorrhagic Fever Protects against Disease and Death in Two Lethal Mouse Models. PLoS Negl. Trop. Dis. 2017, 11, e0005908. [Google Scholar] [CrossRef] [PubMed]
- Buttigieg, K.R.; Dowall, S.D.; Findlay-Wilson, S.; Miloszewska, A.; Rayner, E.; Hewson, R.; Carroll, M.W. A Novel Vaccine against Crimean-Congo Haemorrhagic Fever Protects 100% of Animals against Lethal Challenge in a Mouse Model. PLoS ONE 2014, 9, e91516. [Google Scholar] [CrossRef]
- Deyde, V.M.; Khristova, M.L.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Crimean-Congo Hemorrhagic Fever Virus Genomics and Global Diversity. J. Virol. 2006, 80, 8834–8842. [Google Scholar] [CrossRef]
- Moming, A.; Tuoken, D.; Yue, X.; Xu, W.; Guo, R.; Liu, D.; Li, Y.; Hu, Z.; Deng, F.; Zhang, Y.; et al. Mapping of B-Cell Epitopes on the N- Terminal and C-Terminal Segment of Nucleocapsid Protein from Crimean-Congo Hemorrhagic Fever Virus. PLoS ONE 2018, 13, e0204264. [Google Scholar] [CrossRef] [PubMed]
- Brandariz-Nuñez, A.; Menaya-Vargas, R.; Benavente, J.; Martinez-Costas, J. A Versatile Molecular Tagging Method for Targeting Proteins to Avian Reovirus muNS Inclusions. Use in Protein Immobilization and Purification. PLoS ONE 2010, 5, e13961. [Google Scholar] [CrossRef] [PubMed]
- Brandariz-Nuñez, A.; Menaya-Vargas, R.; Benavente, J.; Martinez-Costas, J. Avian Reovirus microNS Protein Forms Homo-Oligomeric Inclusions in a Microtubule-Independent Fashion, Which Involves Specific Regions of Its C-Terminal Domain. J. Virol. 2010, 84, 4289–4301. [Google Scholar] [CrossRef]
- Marín-López, A.; Barreiro-Piñeiro, N.; Utrilla-Trigo, S.; Barriales, D.; Benavente, J.; Nogales, A.; Martínez-Costas, J.; Ortego, J.; Calvo-Pinilla, E. Cross-Protective Immune Responses against African Horse Sickness Virus after Vaccination with Protein NS1 Delivered by Avian Reovirus muNS Microspheres and Modified Vaccinia Virus Ankara. Vaccine 2020, 38, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, G.; López-Gil, E.; Ortego, J.; Brun, A. Efficacy of Different DNA and MVA Prime-Boost Vaccination Regimens against a Rift Valley Fever Virus (RVFV) Challenge in Sheep 12 Weeks Following Vaccination. Vet. Res. 2018, 49, 21. [Google Scholar] [CrossRef]
- Lorenzo, M.M.; Marín-López, A.; Chiem, K.; Jimenez-Cabello, L.; Ullah, I.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Lorenzo, G.; Moreno, S.; Ye, C.; et al. Vaccinia Virus Strain MVA Expressing a Prefusion-Stabilized SARS-CoV-2 Spike Glycoprotein Induces Robust Protection and Prevents Brain Infection in Mouse and Hamster Models. Vaccines 2023, 11, 1006. [Google Scholar] [CrossRef]
- Pérez, P.; Astorgano, D.; Albericio, G.; Flores, S.; Sánchez-Corzo, C.; Noriega, M.A.; Sánchez-Cordón, P.J.; Labiod, N.; Delgado, R.; Casasnovas, J.M.; et al. MVA-Based Vaccine Candidates Expressing SARS-CoV-2 Prefusion-Stabilized Spike Proteins of the Wuhan, Beta or Omicron BA.1 Variants Protect Transgenic K18-hACE2 Mice against Omicron Infection and Elicit Robust and Broad Specific Humoral and Cellular Immune Responses. Front. Immunol. 2024, 15, 1420304. [Google Scholar] [CrossRef]
- Barreiro-Piñeiro, N.; Lostalé-Seijo, I.; Varela-Calviño, R.; Benavente, J.; Martínez-Costas, J.M. IC-Tagging Methodology Applied to the Expression of Viral Glycoproteins and the Difficult-to-Express Membrane-Bound IGRP Autoantigen. Sci. Rep. 2018, 8, 16286. [Google Scholar] [CrossRef]
- Tipih, T.; Burt, F.J. Crimean–Congo Hemorrhagic Fever Virus: Advances in Vaccine Development. Bioresearch Open Access 2020, 9, 137–150. [Google Scholar] [CrossRef]
- Muzammil, K.; Rayyani, S.; Abbas Sahib, A.; Gholizadeh, O.; Naji Sameer, H.; Jwad Kazem, T.; Badran Mohammed, H.; Ghafouri Kalajahi, H.; Zainul, R.; Yasamineh, S. Recent Advances in Crimean-Congo Hemorrhagic Fever Virus Detection, Treatment, and Vaccination: Overview of Current Status and Challenges. Biol. Proced. Online 2024, 26, 20. [Google Scholar] [CrossRef]
- Di Bella, S.; Babich, S.; Luzzati, R.; Cavasio, R.A.; Massa, B.; Braccialarghe, N.; Zerbato, V.; Iannetta, M. Crimean-Congo Haemorrhagic Fever (CCHF): Present and Future Therapeutic Armamentarium. Infez. Med. 2024, 32, 421–433. [Google Scholar] [CrossRef]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Hunter, L.; Watson, R.; Taylor, I.; Rule, A.; Carroll, M.W.; Hewson, R. Protective Effects of a Modified Vaccinia Ankara-Based Vaccine Candidate against Crimean-Congo Haemorrhagic Fever Virus Require Both Cellular and Humoral Responses. PLoS ONE 2016, 11, e0156637. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ding, Z.; Lan, J.; Wong, G. Advances and Perspectives in the Development of Vaccines against Highly Pathogenic Bunyaviruses. Front. Cell. Infect. Microbiol. 2023, 13, 1174030. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Weaver, G.; Raabe, V.; State of the Clinical Science Working Group of the National Emerging Pathogens Training and Education Center’s Special Pathogens Research Network. Crimean-Congo Hemorrhagic Fever Virus for Clinicians-Diagnosis, Clinical Management, and Therapeutics. Emerg. Infect. Dis. 2024, 30, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Shin, O.S.; Monticelli, S.R.; Hjorth, C.K.; Hornet, V.; Doyle, M.; Abelson, D.; Kuehne, A.I.; Wang, A.; Bakken, R.R.; Mishra, A.K.; et al. Crimean-Congo Hemorrhagic Fever Survivors Elicit Protective Non-Neutralizing Antibodies That Target 11 Overlapping Regions on Glycoprotein GP38. Cell Rep. 2024, 43, 114502. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime-Boost Vaccine Strategy against Viral Infections: Mechanisms and Benefits. Vaccine 2016, 34, 413–423. [Google Scholar] [CrossRef]
- Dowall, S.D.; Carroll, M.W.; Hewson, R. Development of Vaccines against Crimean-Congo Haemorrhagic Fever Virus. Vaccine 2017, 35, 6015–6023. [Google Scholar] [CrossRef]
- Wampande, E.M.; Waiswa, P.; Allen, D.J.; Hewson, R.; Frost, S.D.W.; Stubbs, S.C.B. Phylogenetic Characterization of Crimean-Congo Hemorrhagic Fever Virus Detected in African Blue Ticks Feeding on Cattle in a Ugandan Abattoir. Microorganisms 2021, 9, 438. [Google Scholar] [CrossRef]
- Suschak, J.J.; Golden, J.W.; Fitzpatrick, C.J.; Shoemaker, C.J.; Badger, C.V.; Schmaljohn, C.S.; Garrison, A.R. A CCHFV DNA Vaccine Protects against Heterologous Challenge and Establishes GP38 as Immunorelevant in Mice. NPJ Vaccines 2021, 6, 31. [Google Scholar] [CrossRef]
- Leventhal, S.S.; Bisom, T.; Clift, D.; Rao, D.; Meade-White, K.; Shaia, C.; Murray, J.; Mihalakakos, E.A.; Hinkley, T.; Reynolds, S.J.; et al. Antibodies Targeting the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein Protect via TRIM21. Nat. Commun. 2024, 15, 9236. [Google Scholar] [CrossRef]
- Karaaslan, E.; Çetin, N.S.; Kalkan-Yazıcı, M.; Hasanoğlu, S.; Karakeçili, F.; Özdarendeli, A.; Kalkan, A.; Kılıç, A.O.; Doymaz, M.Z. Immune Responses in Multiple Hosts to Nucleocapsid Protein (NP) of Crimean-Congo Hemorrhagic Fever Virus (CCHFV). PLoS Negl. Trop. Dis. 2021, 15, e0009973. [Google Scholar] [CrossRef] [PubMed]
- Goedhals, D.; Paweska, J.T.; Burt, F.J. Long-Lived CD8+ T Cell Responses Following Crimean-Congo Haemorrhagic Fever Virus Infection. PLoS Negl. Trop. Dis. 2017, 11, e0006149. [Google Scholar] [CrossRef]
- Hawman, D.W.; Ahlén, G.; Appelberg, K.S.; Meade-White, K.; Hanley, P.W.; Scott, D.; Monteil, V.; Devignot, S.; Okumura, A.; Weber, F.; et al. A DNA-Based Vaccine Protects against Crimean-Congo Haemorrhagic Fever Virus Disease in a Cynomolgus Macaque Model. Nat. Microbiol. 2021, 6, 187–195. [Google Scholar] [CrossRef]
- Dowall, S.D.; Buttigieg, K.R.; Findlay-Wilson, S.J.D.; Rayner, E.; Pearson, G.; Miloszewska, A.; Graham, V.A.; Carroll, M.W.; Hewson, R. A Crimean-Congo Hemorrhagic Fever (CCHF) Viral Vaccine Expressing Nucleoprotein Is Immunogenic but Fails to Confer Protection against Lethal Disease. Hum. Vaccin. Immunother. 2016, 12, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-L.; Zhang, L.-Q.; Liu, X.-Q.; Ye, W.; Zhao, Y.-X.; Zhang, L.; Qiang, Z.-X.; Zhang, L.-X.; Lei, Y.-F.; Jiang, D.-B.; et al. Construction and Evaluation of DNA Vaccine Encoding Crimean Congo Hemorrhagic Fever Virus Nucleocapsid Protein, Glycoprotein N-Terminal and C-Terminal Fused with LAMP1. Front. Cell. Infect. Microbiol. 2023, 13, 1121163. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.E.; Gilbride, C.; Dowall, S.; Morris, S.; Ulaszewska, M.; Spencer, A.J.; Rayner, E.; Graham, V.A.; Kennedy, E.; Thomas, K.; et al. Adenoviral Vectored Vaccination Protects against Crimean-Congo Haemorrhagic Fever Disease in a Lethal Challenge Model. EBioMedicine 2023, 90, 104523. [Google Scholar] [CrossRef]
- Scholte, F.E.M.; Karaaslan, E.; O’Neal, T.J.; Sorvillo, T.E.; Genzer, S.C.; Welch, S.R.; Coleman-McCray, J.D.; Spengler, J.R.; Kainulainen, M.H.; Montgomery, J.M.; et al. Vaccination with the Crimean-Congo Hemorrhagic Fever Virus Viral Replicon Vaccine Induces NP-Based T-Cell Activation and Antibodies Possessing Fc-Mediated Effector Functions. Front. Cell. Infect. Microbiol. 2023, 13, 1233148. [Google Scholar] [CrossRef]
- Verheust, C.; Goossens, M.; Pauwels, K.; Breyer, D. Biosafety Aspects of Modified Vaccinia Virus Ankara (MVA)-Based Vectors Used for Gene Therapy or Vaccination. Vaccine 2012, 30, 2623–2632. [Google Scholar] [CrossRef]
- Marín-López, A.; Otero-Romero, I.; de la Poza, F.; Menaya-Vargas, R.; Calvo-Pinilla, E.; Benavente, J.; Martínez-Costas, J.M.; Ortego, J. VP2, VP7, and NS1 Proteins of Bluetongue Virus Targeted in Avian Reovirus muNS-Mi Microspheres Elicit a Protective Immune Response in IFNAR(−/−) Mice. Antivir. Res. 2014, 110, 42–51. [Google Scholar] [CrossRef]
- Marín-López, A.; Calvo-Pinilla, E.; Barriales, D.; Lorenzo, G.; Benavente, J.; Brun, A.; Martínez-Costas, J.M.; Ortego, J. Microspheres-Prime/rMVA-Boost Vaccination Enhances Humoral and Cellular Immune Response in IFNAR(−/−) Mice Conferring Protection against Serotypes 1 and 4 of Bluetongue Virus. Antivir. Res. 2017, 142, 55–62. [Google Scholar] [CrossRef]
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Barreiro-Piñeiro, N.; Pose-Boirazian, T.; Martínez-Costas, J.; Marín-López, A.; Ortego, J. Nanoparticle- and Microparticle-Based Vaccines against Orbiviruses of Veterinary Importance. Vaccines 2022, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.; Kim, S.; Anandasabapathy, N. Vaccinia Virus: Mechanisms Supporting Immune Evasion and Successful Long-Term Protective Immunity. Viruses 2024, 16, 870. [Google Scholar] [CrossRef] [PubMed]
- Lázaro-Frías, A.; Pérez, P.; Zamora, C.; Sánchez-Cordón, P.J.; Guzmán, M.; Luczkowiak, J.; Delgado, R.; Casasnovas, J.M.; Esteban, M.; García-Arriaza, J. Full Efficacy and Long-Term Immunogenicity Induced by the SARS-CoV-2 Vaccine Candidate MVA-CoV2-S in Mice. NPJ Vaccines 2022, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guo, Y.; Deng, Y.; Gao, W.; Huang, B.; Yao, W.; Zhao, Y.; Zhang, Q.; Huang, M.; Liu, M.; et al. Long-Lasting Humoral and Cellular Memory Immunity to Vaccinia Virus Tiantan Provides Pre-Existing Immunity against Mpox Virus in Chinese Population. Cell Rep. 2024, 43, 113609. [Google Scholar] [CrossRef]
- Marín-Hernández, D.; Nixon, D.F.; Hupert, N. Heterologous Vaccine Interventions: Boosting Immunity Against Future Pandemics. Mol. Med. 2021, 27, 54. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Position/Size | Sequence |
|---|---|---|
| p1 | 140–148/9 | RVNANTAAL |
| p2 | 146–154/9 | AALSNKVLA |
| p3 | 272–280/9 | SADSMITNL |
| p4 | 275–284/10 | SMITNLLKHI |
| p5 | 306–314/9 | TAFSSYYWL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvo-Pinilla, E.; Moreno, S.; Barreiro-Piñeiro, N.; Sánchez-Puig, J.M.; Blasco, R.; Martínez-Costas, J.; Brun, A.; Lorenzo, G. Prime-Boost Vaccination Based on Nanospheres and MVA Encoding the Nucleoprotein of Crimean-Congo Hemorrhagic Fever Virus Elicits Broad Immune Responses. Vaccines 2025, 13, 291. https://doi.org/10.3390/vaccines13030291
Calvo-Pinilla E, Moreno S, Barreiro-Piñeiro N, Sánchez-Puig JM, Blasco R, Martínez-Costas J, Brun A, Lorenzo G. Prime-Boost Vaccination Based on Nanospheres and MVA Encoding the Nucleoprotein of Crimean-Congo Hemorrhagic Fever Virus Elicits Broad Immune Responses. Vaccines. 2025; 13(3):291. https://doi.org/10.3390/vaccines13030291
Chicago/Turabian StyleCalvo-Pinilla, Eva, Sandra Moreno, Natalia Barreiro-Piñeiro, Juana M. Sánchez-Puig, Rafael Blasco, José Martínez-Costas, Alejandro Brun, and Gema Lorenzo. 2025. "Prime-Boost Vaccination Based on Nanospheres and MVA Encoding the Nucleoprotein of Crimean-Congo Hemorrhagic Fever Virus Elicits Broad Immune Responses" Vaccines 13, no. 3: 291. https://doi.org/10.3390/vaccines13030291
APA StyleCalvo-Pinilla, E., Moreno, S., Barreiro-Piñeiro, N., Sánchez-Puig, J. M., Blasco, R., Martínez-Costas, J., Brun, A., & Lorenzo, G. (2025). Prime-Boost Vaccination Based on Nanospheres and MVA Encoding the Nucleoprotein of Crimean-Congo Hemorrhagic Fever Virus Elicits Broad Immune Responses. Vaccines, 13(3), 291. https://doi.org/10.3390/vaccines13030291