1. Introduction
Poliomyelitis is an infectious viral disease caused by the three serotypes of poliovirus. It affects the central nervous system and can cause temporary or permanent paralysis. There are two excellent vaccines that protect against poliomyelitis. Inactivated poliovirus vaccine (IPV) was developed by Jonas Salk in 1955 [
1] and is administered by injection. The live attenuated poliovirus vaccine (OPV) developed by Albert Sabin in 1961 is administered orally [
2,
3]. Both vaccines are highly effective; however, recipients of OPV excrete mutant variants of Sabin strains [
4,
5] in stools that can become pathogenic vaccine-derived polioviruses and circulate in the population, occasionally causing disease in unvaccinated persons [
6,
7]. The limitation of IPV is that it elicits poor intestinal immunity and is not able to stop virus circulation [
8,
9]. Therefore, new improved poliovirus vaccines are under development [
10,
11,
12,
13]. The absence of significant poliovirus morbidity makes clinical evaluation of the new vaccines based on protection endpoint impossible. Therefore, their efficacy can only be assessed using immunogenicity endpoint. Serum neutralizing antibodies present at 1:8 or higher level represent a well-established biomarker of protection. In addition, the serum neutralization test is a crucial part of the rat potency test for IPV. Poliovirus neutralizing antibody titer is determined by a standard poliovirus neutralization test [
14,
15] using serial dilutions of serum samples that are incubated in 96-well microtiter plates with a fixed amount of poliovirus. After the incubation, a suspension of susceptible cells is added to the serum-poliovirus complex and incubated for 10 days [
14]. The standard poliovirus neutralization test requires personnel expertise to observe and interpret the cytopathic effect (CPE) in infected cells, takes 7 to 10 days to generate the results and is not suitable for large-scale analysis. Determination of neutralizing antibodies to each of the three serotypes of poliovirus must also be performed in a separate assay. Critically, the test produces variable results [
16,
17].
In the end-game of the polio eradication program, surveillance of seroprevalence against poliovirus in susceptible populations is fundamental for monitoring the exposure to poliovirus circulation in poliovirus-free countries to maintain the poliovirus-free status and the seroconversion rates in countries where poliovirus is endemic to assess the efficacy of vaccination programs. In these cases, it is important to generate data about the presence of anti-poliovirus antibodies in the shortest time possible and high-throughput manner to expedite the development of the improved poliovirus vaccines and to analyze the seroprevalence of population needed for the poliovirus eradication end-game. Therefore, a faster multiplex and high-throughput assay would be a desirable alternative to the conventional assay.
This communication describes the development of a multiplex PCR-based neutralization (MPBN) assay for anti-poliovirus antibodies titration that uses a quantitative multiplex one-step RT-PCR (qmosRT-PCR) [
18] as a read-out instead of CPE used in the conventional assay. Recently, we successfully used a similar approach to develop a multiplex PCR-based titration (MPBT) assay for the simultaneous titration of the three poliovirus serotypes [
19].
The MPBN assay generates similar results as the conventional neutralization assay while reducing the time to generate results and the variation that is due to the interpretation of CPE. It is reproducible, sensitive, robust and allows titration of anti-poliovirus sera against all three serotypes in the same reaction.
2. Materials and Methods
2.1. Hep-2C Cells, Viruses and Sera
HEp-2C cells (ATCC® CCL-23TM), derived from a human carcinoma, were cultured in 175-cm2 flasks at 37 °C ± 2 °C in Dulbecco’s modified Eagle’s medium (DMEM; ThermoFischer, Frederick, MD, USA) supplemented with 5% fetal calf serum (FCS; ThermoFischer, Frederick, MD, USA) and penicillin-streptomycin (100 U/mL and 100 µg/ mL, respectively; ThermoFischer, Frederick, MD, USA). Viable cells were sampled, stained with Trypan Blue (ThermoFischer, Frederick, MD, USA) and counted with a hemocytometer (CountessTM; ThermoFischer, Frederick, MD, USA). One hundred microliters of cell suspension containing 4 × 104 cells for MPBN assays and 1–2 × 104 cells for conventional neutralization assay was added to each well of a 96-well microplate.
Lots of US neurovirulence poliovirus reference vaccines (Sabin type 1, 2 and 3 OPV strains having GenBank accession numbers AY184219, AY184220 and AY184221, respectively) were used as challenges to compare results of the conventional neutralization and MPBN assays. Poliovirus sera were obtained from our Laboratory of Method Development stock via Dr. Diana Kouiavskaia (CBER, FDA).
2.2. Quantitative Multiplex One-Step RT-PCR
The quantitative multiplex one-step RT-PCR (qmosRT-PCR) was described previously [
18]. Briefly, the qmosRT-PCR reactions were prepared in 96-well optical plates in a final volume of 25 μL using 2 μL of 1/20 diluted cell lysate and QuantiFast Multiplex RT-PCR Kit (QIAGEN, Valencia, CA, USA). The diluted lysates of cells infected with Sabin 1, 2 and 3 were used as positive control, and water and diluted lysates of noninfected cells were used as negative controls. All control samples were run in duplicates. The specific primer pairs and probes used for each Sabin strain were: for Sabin 1, forward primer—2771Sab1F, 5′CAGCTTCCACCAAGAATAA3′, reverse primer—3036Sab1R, 5′GATTGATGGATTTGATGAG3′ and probe—Sab1-FAM, FAM-5′ACAGTGTGGAAGATC3′-NFQ; for Sabin 2, forward primer—2682TqS2F, 5′CCAGAGACGAACGCGA3′, reverse primer—2803TqS2R, 5′CAAACCGAAAACAATCTGC3′ and probe—Sab2-VIC, VIC-5′CACGGTTGAGTCATTC3′-NFQ, and, for Sabin 3, forward primer—1411TqS3F 5′GGGAAAATTTTACTCCCAA3′, reverse primer—1629TqS3R 5′TGAATCAATGGCCAAAGCA3′ and probe—Sab3-NED, NED-5′AACGCAGTAACATCC3′-NFQ.
The primers were used at a concentration of 0.4 μM for each primer of poliovirus type 2 and 0.8 μM for each primer of poliovirus type 1 and 3. The TaqMan probes were used at concentration 25 nM for each probe of each poliovirus serotype. The qmosRT-PCR procedure was performed using real-time PCR System ViiA7 (Applied Biosystems, Foster City, CA, USA) at the following thermal cycling conditions: one cycle incubation for 20 min at 50 °C and 5 min at 95 °C, followed by 45 cycles, each consisting of 15 s at 95 °C, 15 s at 50 °C and 50 s at 60 °C.
2.3. Conventional Neutralization Assay
The conventional neutralization assay was performed using Sabin 1, 2 and 3 strains and a HEp-2C cell line according to the World Health Organization (WHO) manual [
15]. Briefly, the titers of neutralizing antibodies against poliovirus serotypes were determined using 96-well microplates. The sera were inactivated at 56 °C for 30 min and subsequently diluted from 1:8 to 1:2048 in two-fold serial dilutions; this dilution was performed in 25 μL. To each well, 25 μL of poliovirus (100 CCID
50/25 μL) was added and incubated for 3 h at 36 °C to allow the antibodies to bind to poliovirus. Next, one-hundred-µL aliquots of cell suspension containing 1–2 × 10
4 HEp-2C cells in DMEM with 2% FCS were added to each well of diluted sera-virus complex in triplicate wells of 96-well plates for each sample. Virus-infected plates were incubated for ten days at 36 °C in a 5% CO
2 humid atmosphere; wells were periodically inspected for the presence of cytopathic effect (CPE). Wells showing CPE were counted on day ten after infection and sera titers calculated using the Spearman-Karber formula [
20].
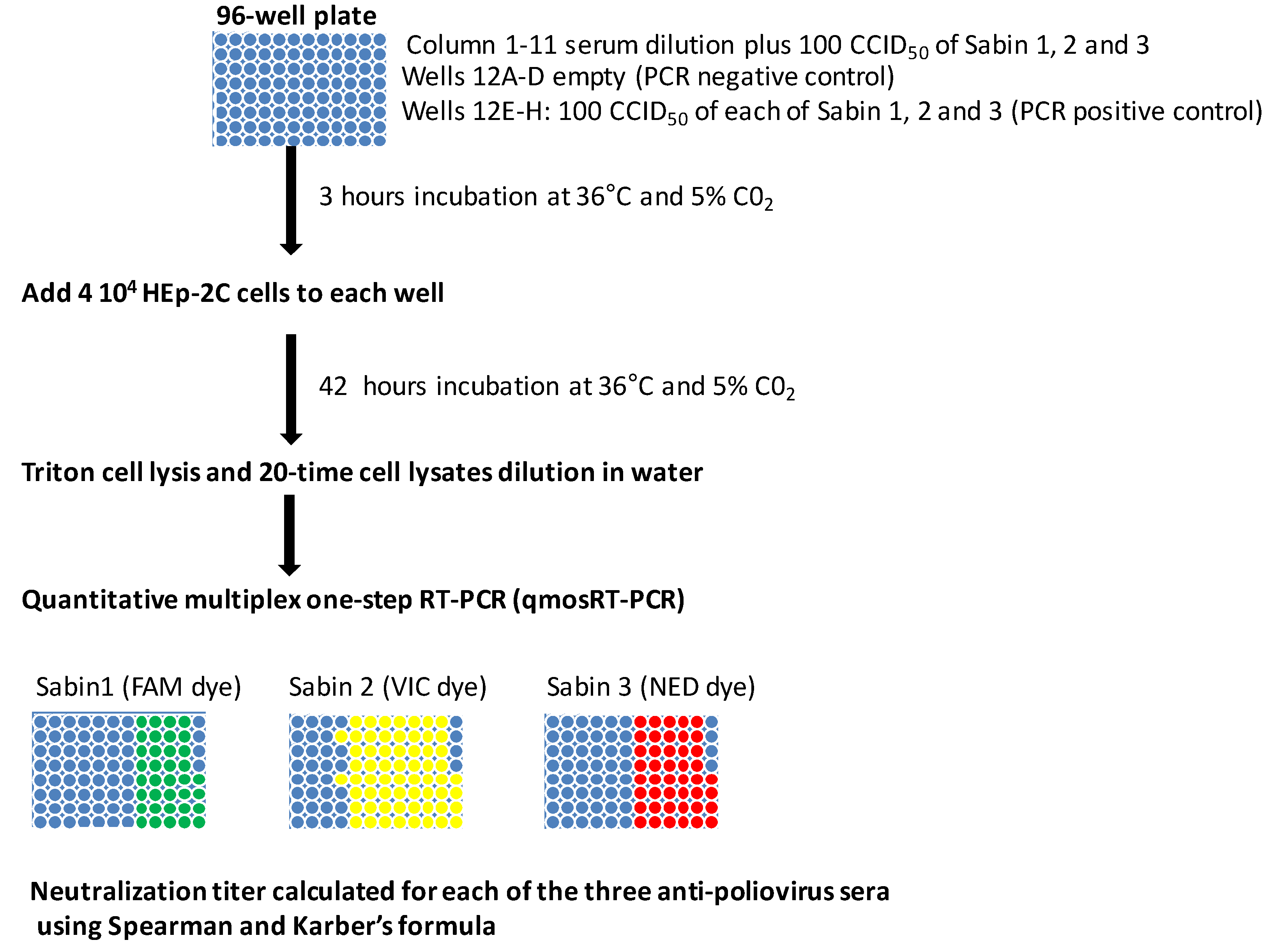
2.4. MPBN Assay and Data Analysis
Like the conventional neutralization assay, the MPBN assay was performed using Sabin 1, 2 and 3 strains and a HEp-2C cell line according to the WHO manual [
15]. The titers of neutralizing antibodies against poliovirus serotypes were determined using the 96-well microplates. The sera were inactivated at 56 °C for 30 min and subsequently diluted from 1:8 to 1:2048 in two-fold serial dilutions; this dilution was performed in 25 μL per well of a 96-well plate. To each well, 25 μL of the three Sabin strains (100 CCID
50 of each) was added and incubated for 3 h at 36 °C to allow the antibodies to bind to poliovirus. Next, one-hundred-µL aliquots of cell suspension containing 4 × 10
4 HEp-2C cells in DMEM with 2% FCS were added to each well of viruses-sera mixture. The plates were then incubated at 36 °C in a 5% CO
2 humid atmosphere for 42 h. Next, the viral medium was aspirated (this allows us to detect the replication of only those viruses that enter the cell) from the wells, and 50 μL of 0.9% Triton X-100 in DMEM was added to each well. The plates were sealed with a foil and stored at −80 °C prior to qmosRT-PCR analysis.
The plates with the triton-lysed cells were thawed for 30 min at room temperature and briefly centrifuged in a 5810R centrifuge (Eppendorf, San Diego, CA, USA) at 1000 rpm for 1 min to collect the liquid at the bottom of the wells. The 1:20 dilutions were prepared by adding 10 μL of the cell lysates to 190 μL of molecular biology grade water (5-PRIME, Gaithersburg, MD, USA).
The three poliovirus serotypes were quantified in the same reaction of qmosRT-PCR as described previously [
18] and summarized above. The positive and negative results for viral replication were recorded for each well and used for sera titers calculation for each poliovirus serotype in accordance with the Spearman-Karber formula [
20].
The samples that had Ct less or equal than 40 were considered positive, and samples with Ct higher than 40 were considered negative. The results of the PCR were considered nonvalid if both reactions of the positive control gave negatives results and/or both reactions of the negative controls gave positive results.
4. Discussion
In 1988, WHO launched the Global Polio Eradication Initiative (GPEI) designed to eradicate poliomyelitis [
22]. At that time, the annual global cases of paralytic poliomyelitis were estimated to be about 350,000 cases, with wild poliovirus (WPV) circulating in most countries. Extensive use of live, attenuated OPV in mass vaccination campaigns improved routine immunization programs and resulted in a gradual decrease of poliomyelitis cases. Presently, only Afghanistan and Pakistan are endemic with wild poliovirus type 1 [
23], but outbreaks caused by virulent vaccine-derived polioviruses of all three types occur regularly. Wild poliovirus type 2 was last detected in 1999 and was declared eradicated in September 2015, while wild poliovirus type 3 has been eliminated since 2012 and declared eradicated in October 2019 [
24]. Inactivated polio vaccine (IPV) is the only polio vaccine that has been given in the United States since the turn of the century. Many other countries still use OPV in their childhood immunization schedule, while others have gradually switched to IPV consistent with the WHO strategy to reduce the emergence of vaccine-derived polioviruses (VDPV) [
25].
Epidemiological and seroprevalence surveillance are an essential part of GPEI [
26]. It is crucial for monitoring the quality of vaccination campaigns and to identify regions that may require supplemental immunizations.
The level of population immunity required for poliovirus elimination must be about above 80% and depends on the level of hygiene [
27]. A neutralizing-antibody titer higher than 1:8 is considered to be protective against paralytic poliomyelitis [
28]. In laboratories, the neutralizing-antibody titer has been determined by a conventional poliovirus neutralization test using a susceptible cell culture system and infectious challenge virus [
14].
The conventional poliovirus neutralization test is used to measure the ability of anti-poliovirus antibodies to inhibit virus cytopathic effects (CPE) in HEp-2c cells. Neutralizing antibody titers are expressed as the reciprocal of the highest serum dilution protecting half of the cell-wells from virus infection [
29]. Virus titration and neutralization assays using CPE endpoint are simple but have some limitations. They are usually time-consuming (7–10 days for poliovirus), and their results are variable because of difficulties in evaluating CPE [
16]. They cannot be used for viruses that do not cause CPE, and they cannot be used in multiplex format. Recently, for many viruses, these assays were modified to include a quantitative non-multiplex PCR for assessing virus replication [
30,
31,
32,
33,
34]. Therefore, to overcome these limitations of the conventional assay, we developed the MPBN assay.
Similar to conventional neutralization assays, the MPBN assay tests serially diluted sera samples mixed with a predetermined amount of a mixture of the three Sabin strains and HEp-2c cells in replicate wells of 96-well plates, except that, instead of waiting ten days for the development of CPE, medium supernatant is discarded after 36–42 h of incubation, cells lysed and viral nucleic acids in cell lysates are quantified by qmosRT-PCR assay; results are used to determine sera titers expressed as the reciprocal of the highest serum dilution protecting cells from virus infection per mL. This way, serum titer is not determined based on proportionality to Ct (cycle threshold) values obtained by qmosRT-PCR, but rather, qmosRT-PCR is used as a readout to determine the presence or the absence of viral replication. A Ct of 40 is used as threshold; Ct less or equal 40 are considered positive for viral replication in the cells. Therefore, qmosRT-PCR results are expressed in yes or no format, allowing a simple Karber formula to be used similar to the conventional neutralization assay.
The MPBN assay proved to be very sensitive, accurately titrating an equivalent of anti-poliovirus sera of 1.55–0.77 per mL for type 1, 1.52–0.76 per mL for type 2 and 1.64–0.82 per mL for type 3 in mixtures (sera against the three poliovirus serotypes together). In addition, the MPBN method generated titers similar (within the margin of a single dilution) to those of conventional assays for all combinations of Sabin challenge strains.
We found excellent correlations between the conventional and MPBN assays for anti-poliovirus sera of all three serotypes with an R
2 value of 0.98 for type 1, 0.99 for Sabin 2 and 0.96 for type 3. This correlation was observed for sera with both high and low titers. The LLOT was within the linear range, indicating that the assay can accurately titrate samples with low titers. The MPBN assay generated consistent and reproducible results for all three sera types and proved to be very robust. Even a one-hour delay in the addition of cells following incubation of the sera-viruses complex, or a five-fold variation in the number of cells, had no effect on neutralization titers (
Table 4 and
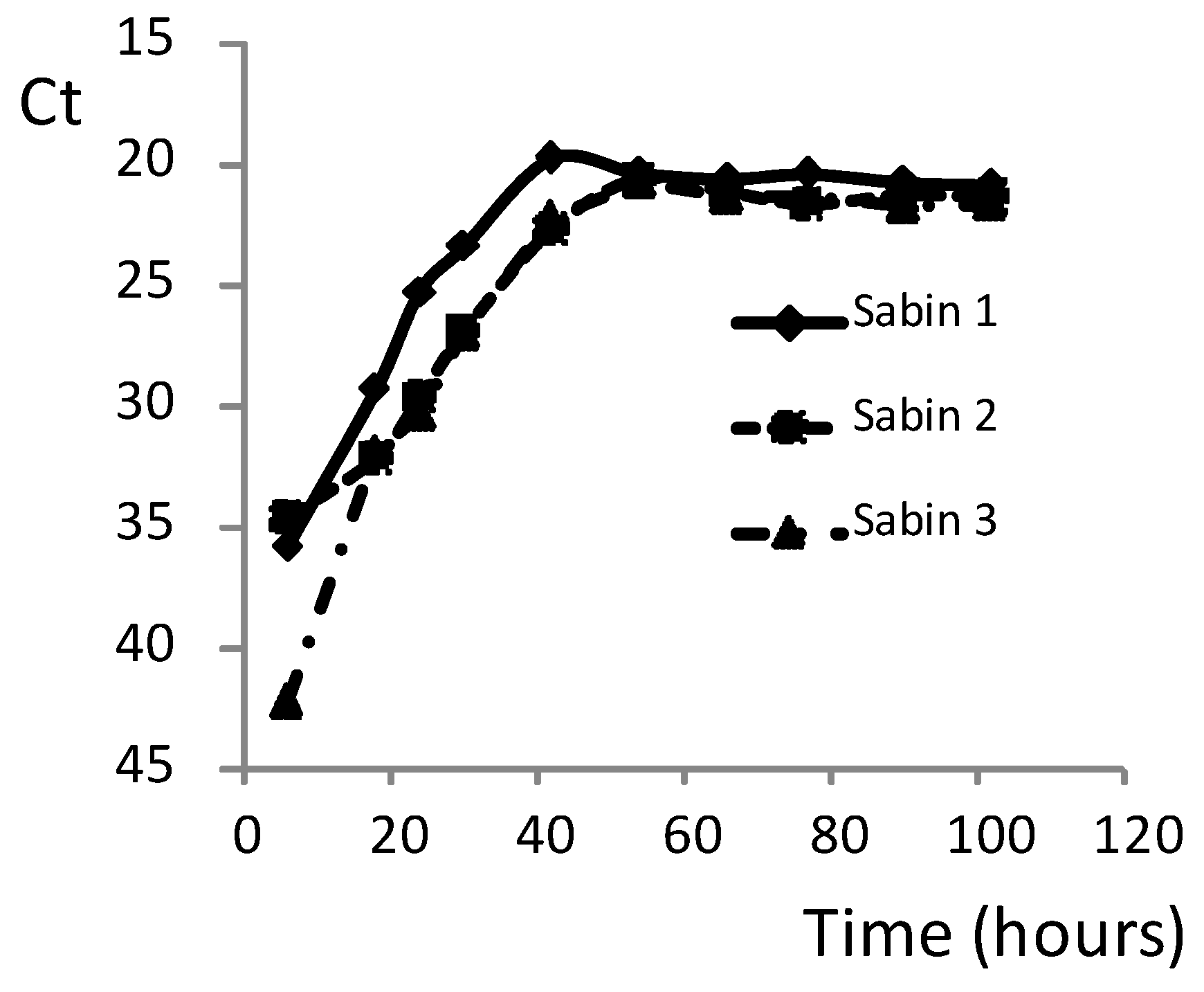
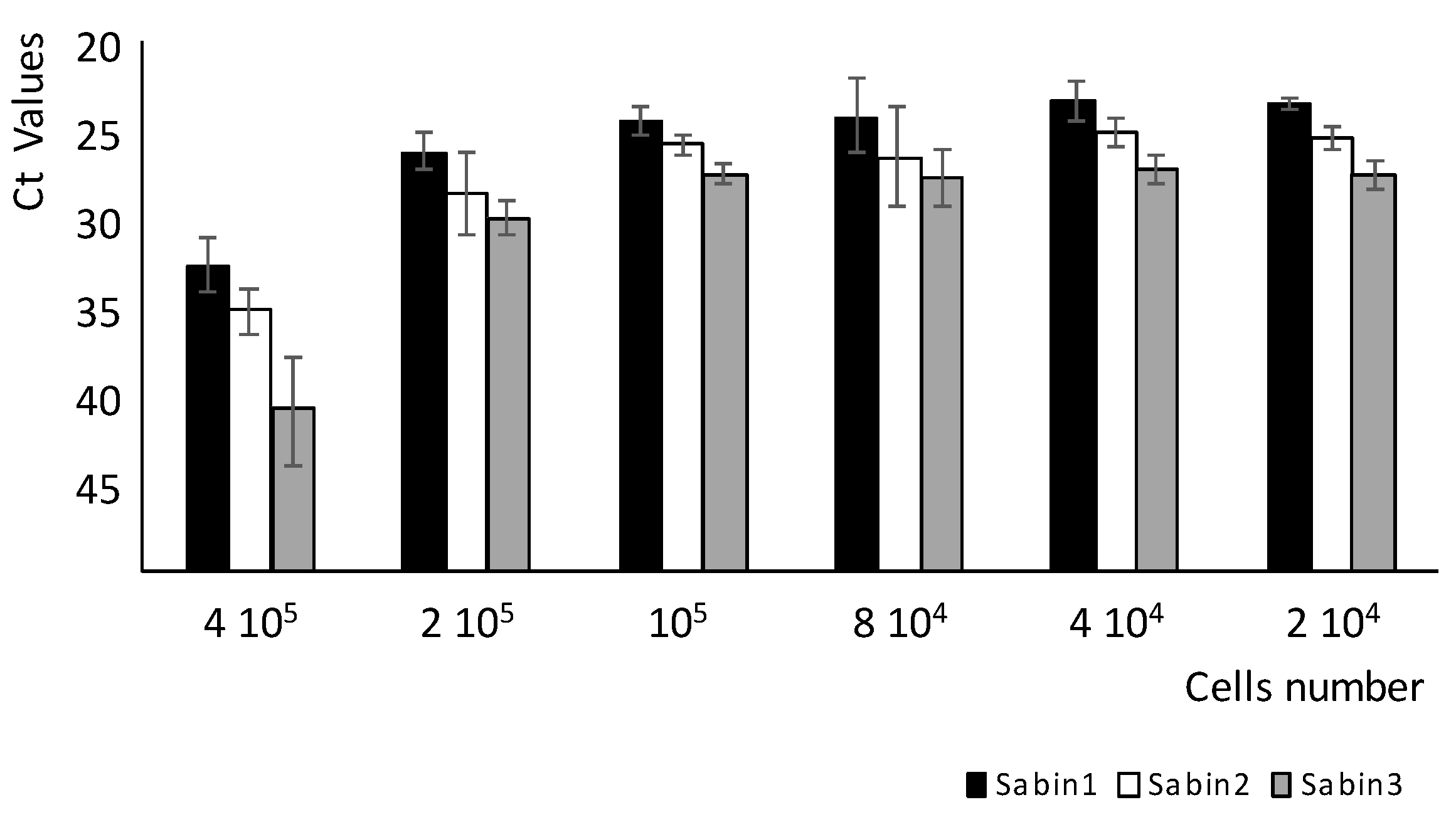
Table 5). This method reduced the time needed to titrate sera against polioviruses from the seven to ten days for conventional neutralization assay to only two to three days. Importantly, the MPBN assay accurately titrated the sera of all three poliovirus serotypes simultaneously in the same reaction-run.
The MPBN assay can be used to analyze seroprevalence of anti-poliovirus antibodies and seroconversion rates in populations. It can also be used for analysis of sera collected from clinical studies of a new generation of polio vaccines and for analysis of animal immunogenicity studies, such as the rat IPV potency assay. The ability to obtain results in just 2–3 days and its multiplex design can make MPBN attractive for time-critical and high-throughput applications.






{kind=link}
{kind=link}
{kind=link}
{kind=link}