Interleukin 34 Serves as a Novel Molecular Adjuvant against Nocardia Seriolae Infection in Largemouth Bass (Micropterus Salmoides)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish, Bacterial Strains, Plasmids, Reagents, and Growth Conditions
2.3. Sequence and Phylogenetic Analysis
2.4. Plasmid Construction and Cloning of pcIL-34 and pcHrp1
2.5. In vitro Expression Analysis of pcIL-34 and pcHrp1
2.6. Vaccination and Sampling
2.7. Challenge
2.8. Detection of pcIL-34 and pcHrp1 in the Muscle of Immnunized Fish by PCR
2.9. In Vivo Transcription Analysis of pcIL-34 and pcHrp1 by RT-PCR
2.10. In vivo Expression Analysis of pcIL-34 and pcHrp1 by Western Blotting
2.11. Western Blot Analysis
2.12. Serum Lysozyme Activity
2.13. Enzyme-Linked Immunosorbent Assay (ELISA)
2.14. qRT-PCR Analysis of the Expression of Immune-Related Genes
2.15. Statistical Analysis
3. Results
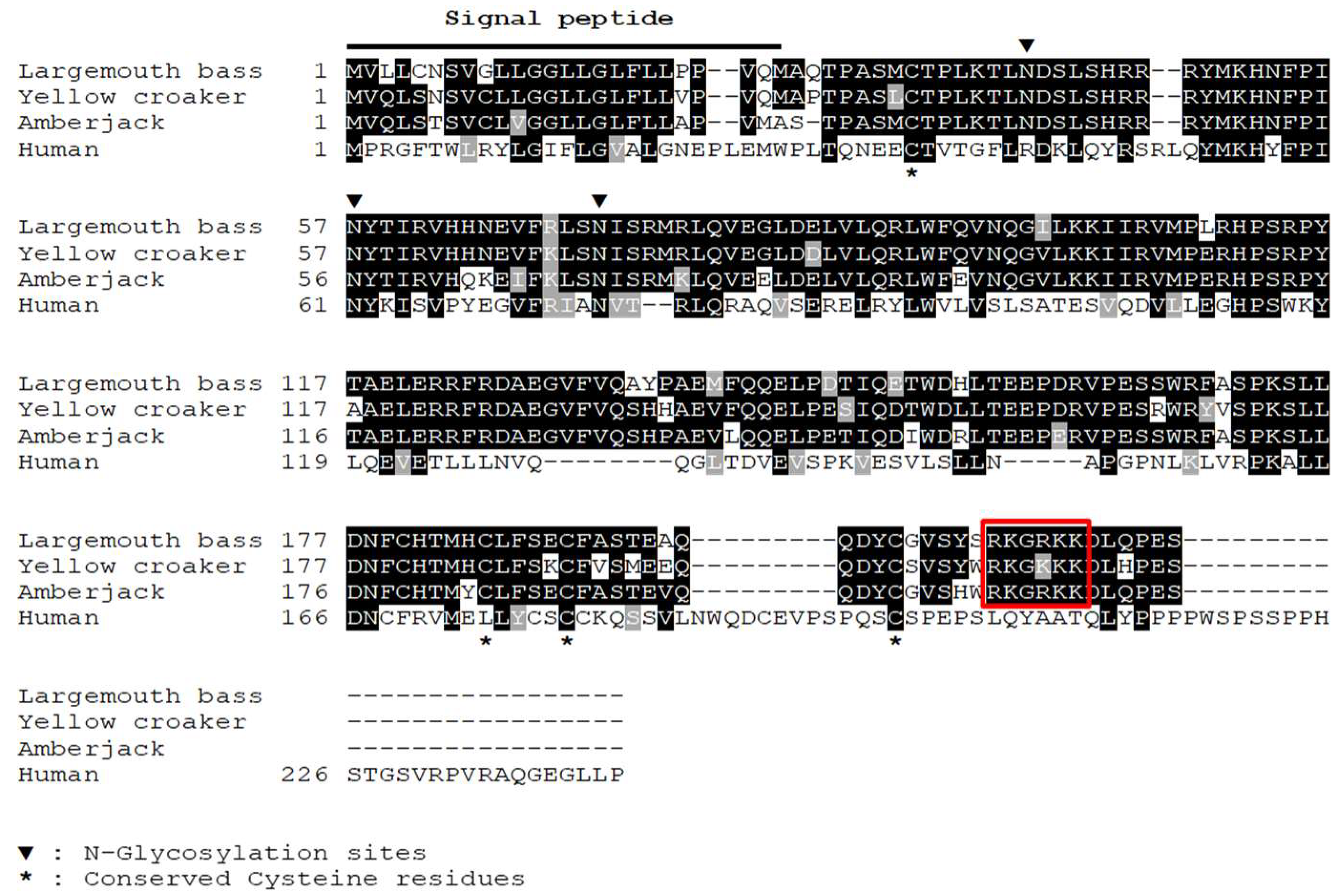
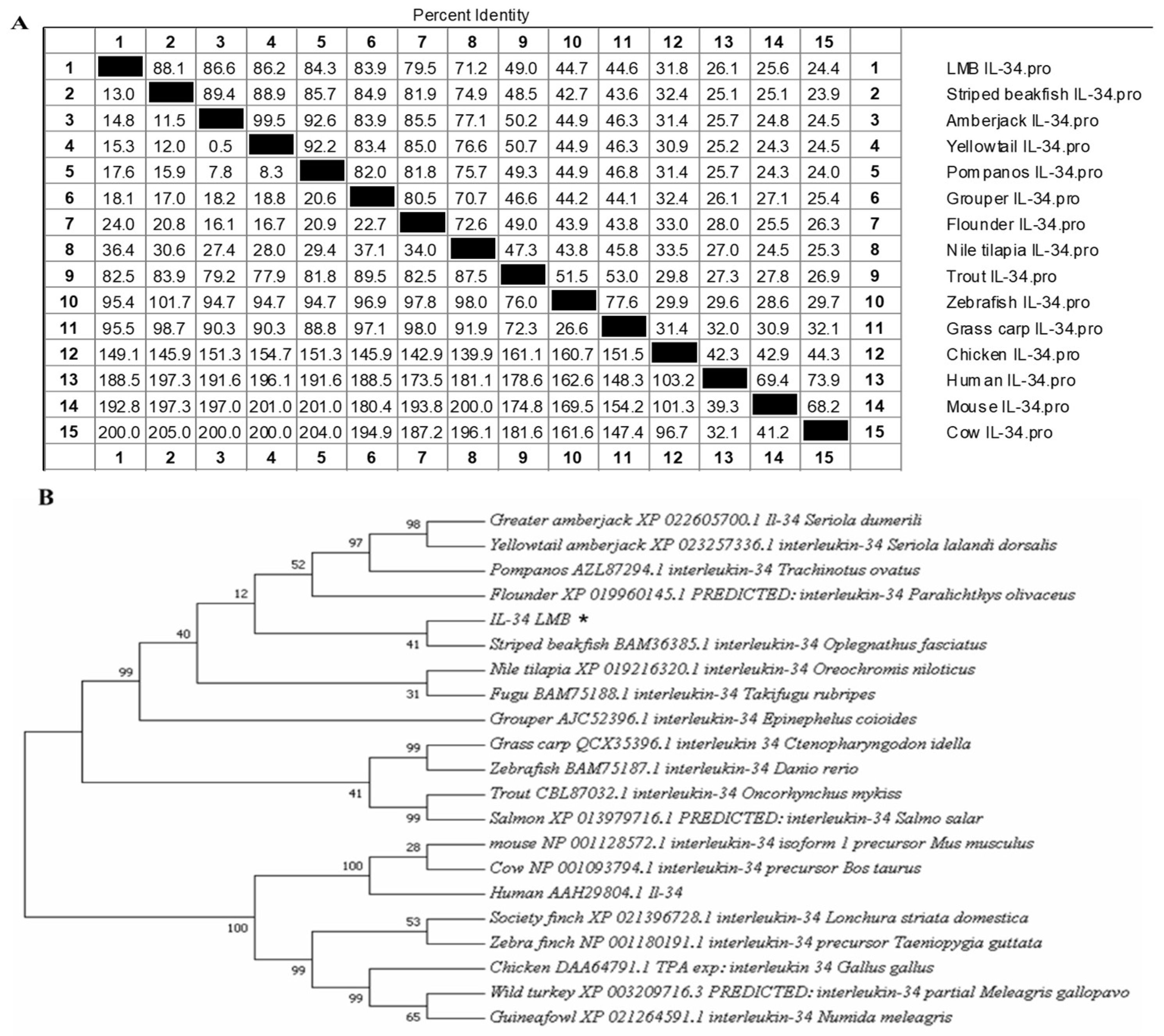
3.1. Sequence Analysis of IL-34
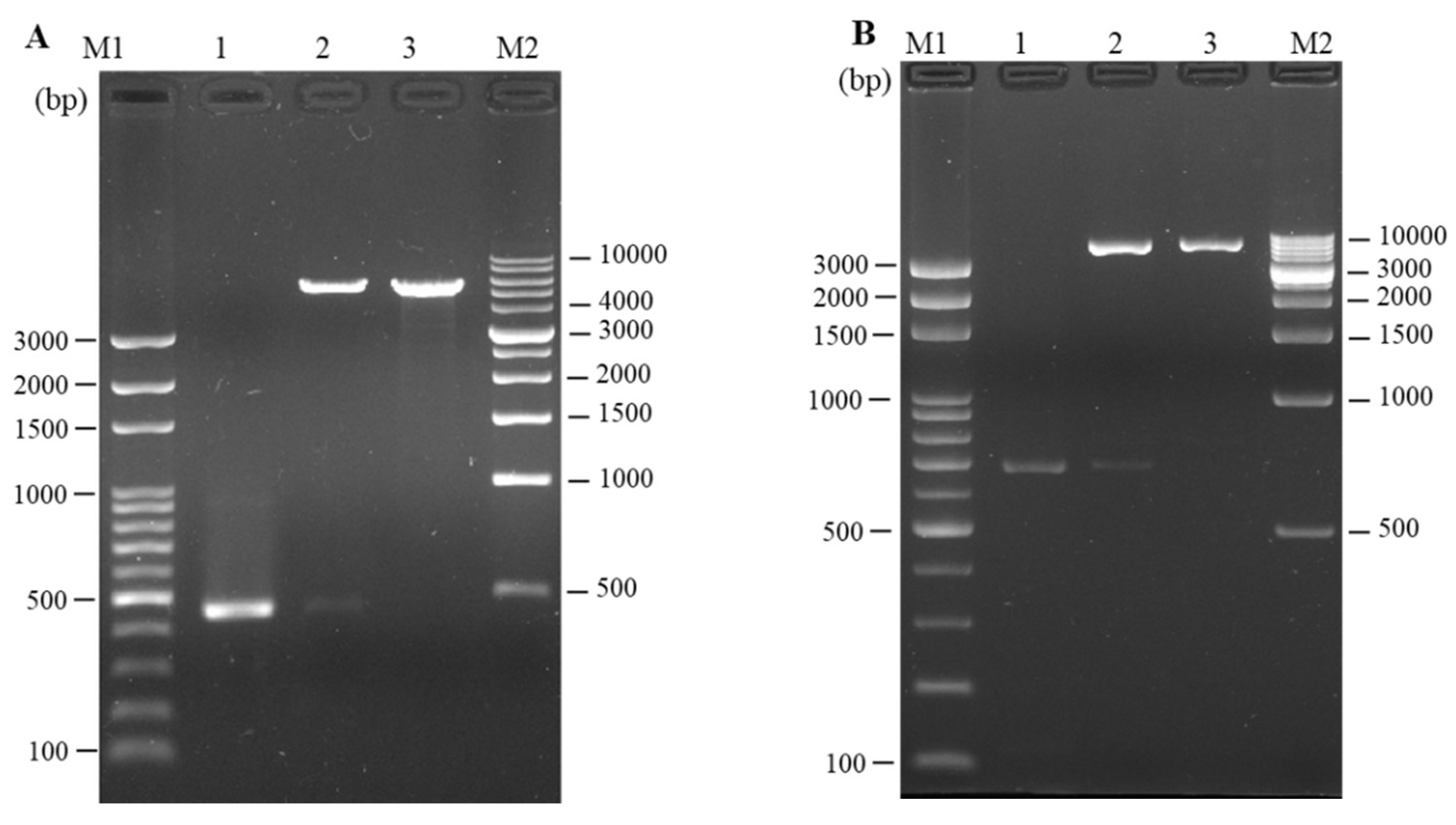
3.2. Cloning and In Vitro Expression of Plasmid pcIL-34 and pcHrp1
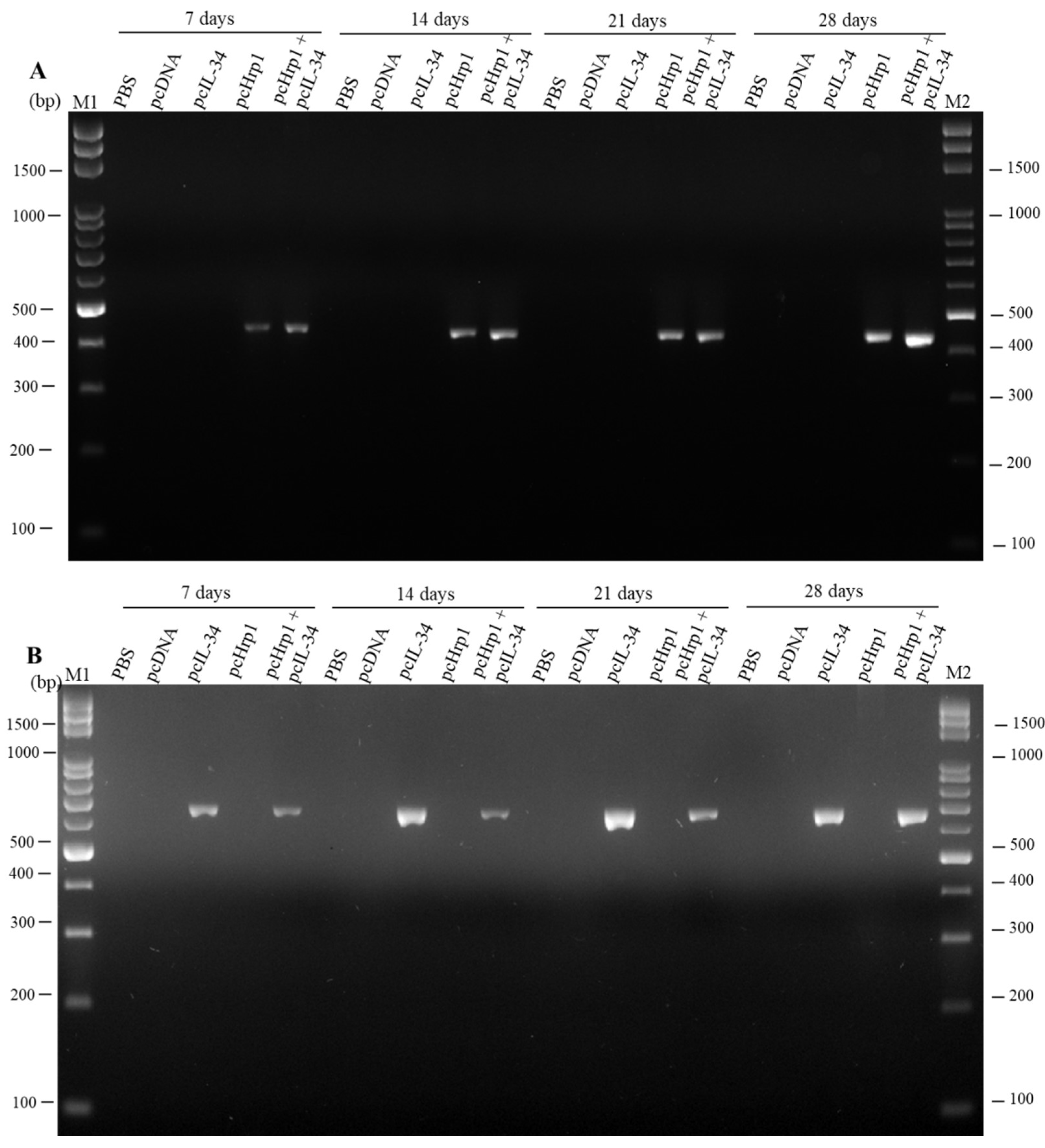
3.3. Detection of pcIL-34 and pcHrp1 in the Muscle of Immnunized Fish by PCR
3.4. In vivo Transcription Analysis of pcIL-34 and pcHrp1 by RT-PCR
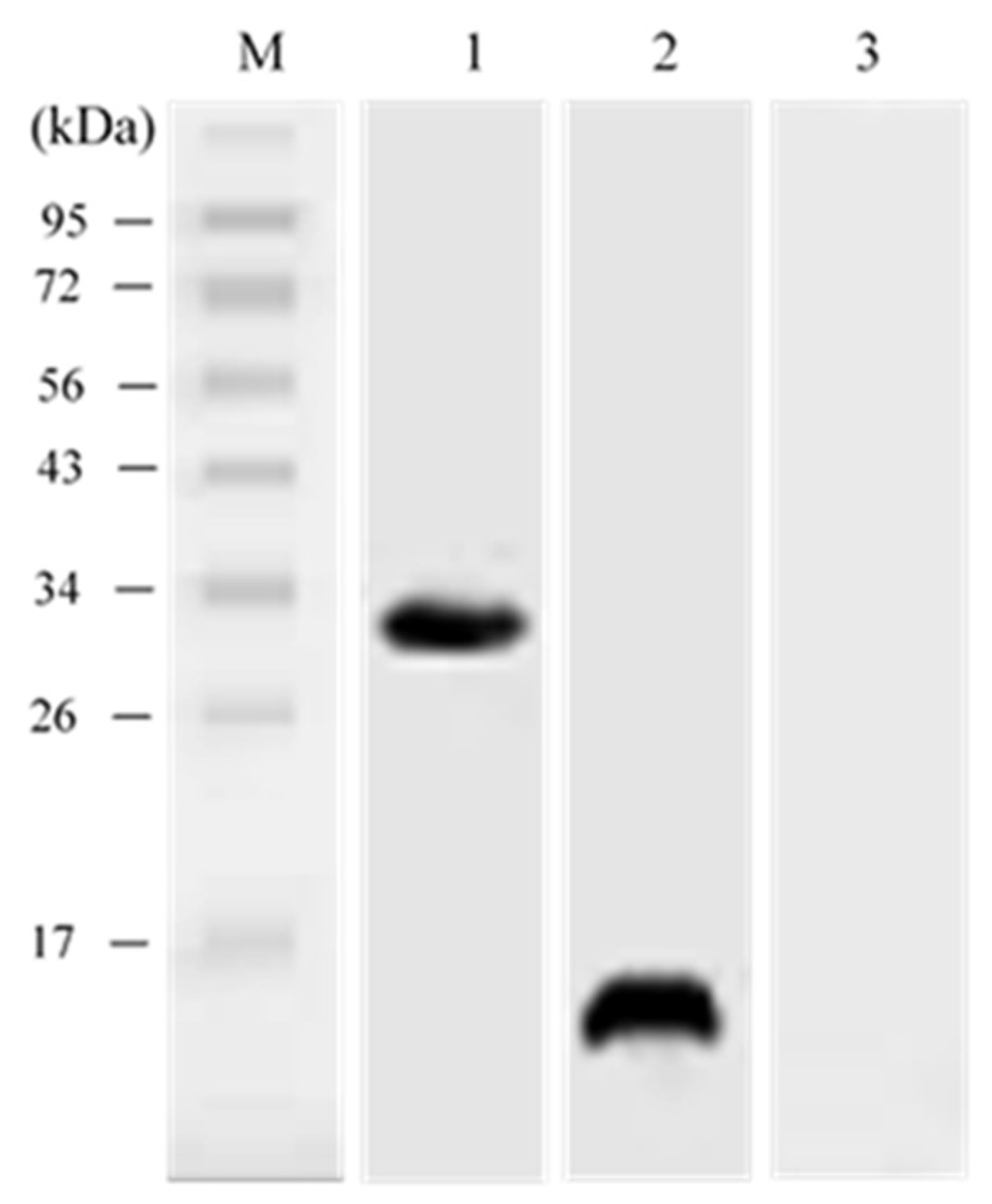
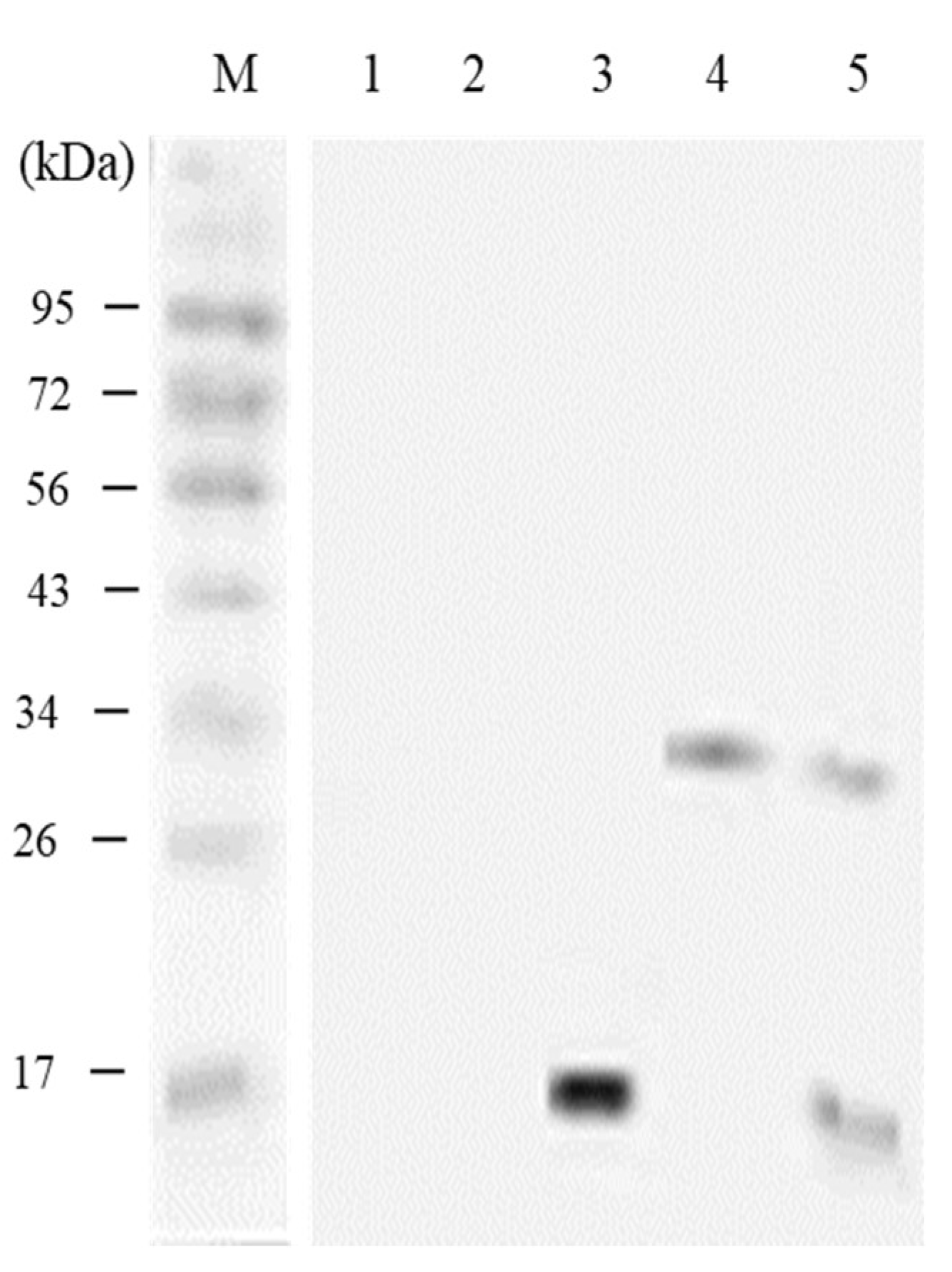
3.5. In vivo Expression Analysis of pcIL-34 and pcHrp1 by Western Blotting
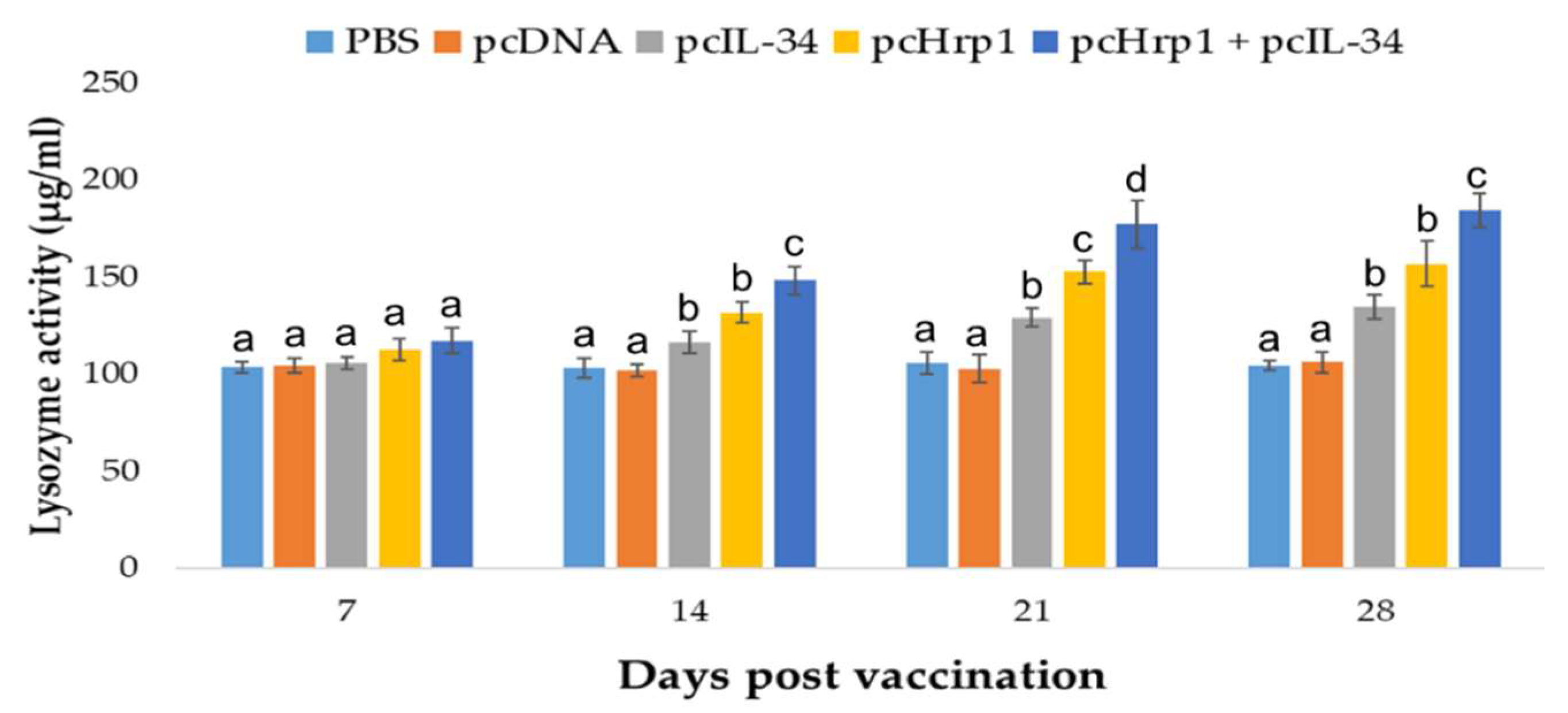
3.6. Serum Lysozyme Activity
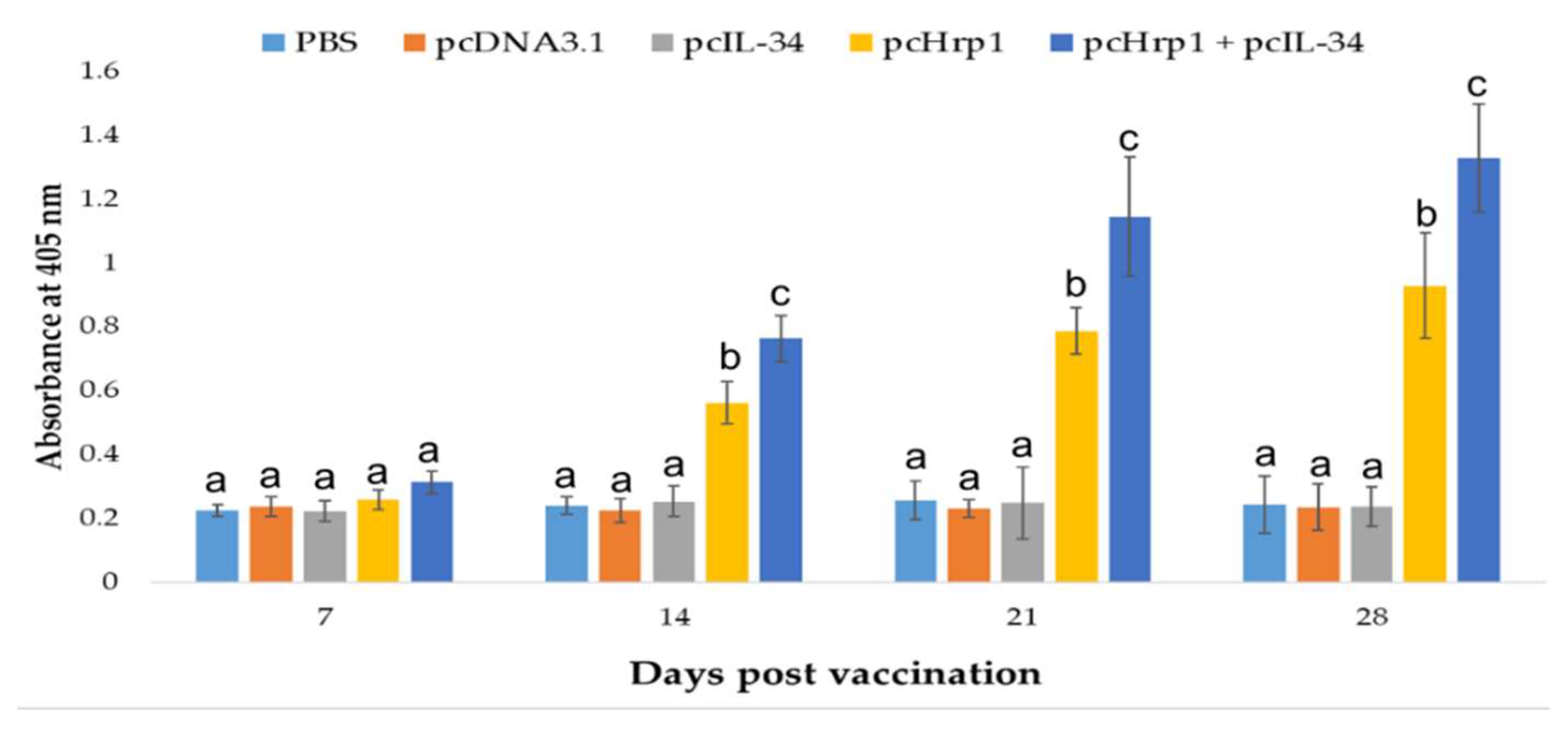
3.7. Specific Serum Antibody Production
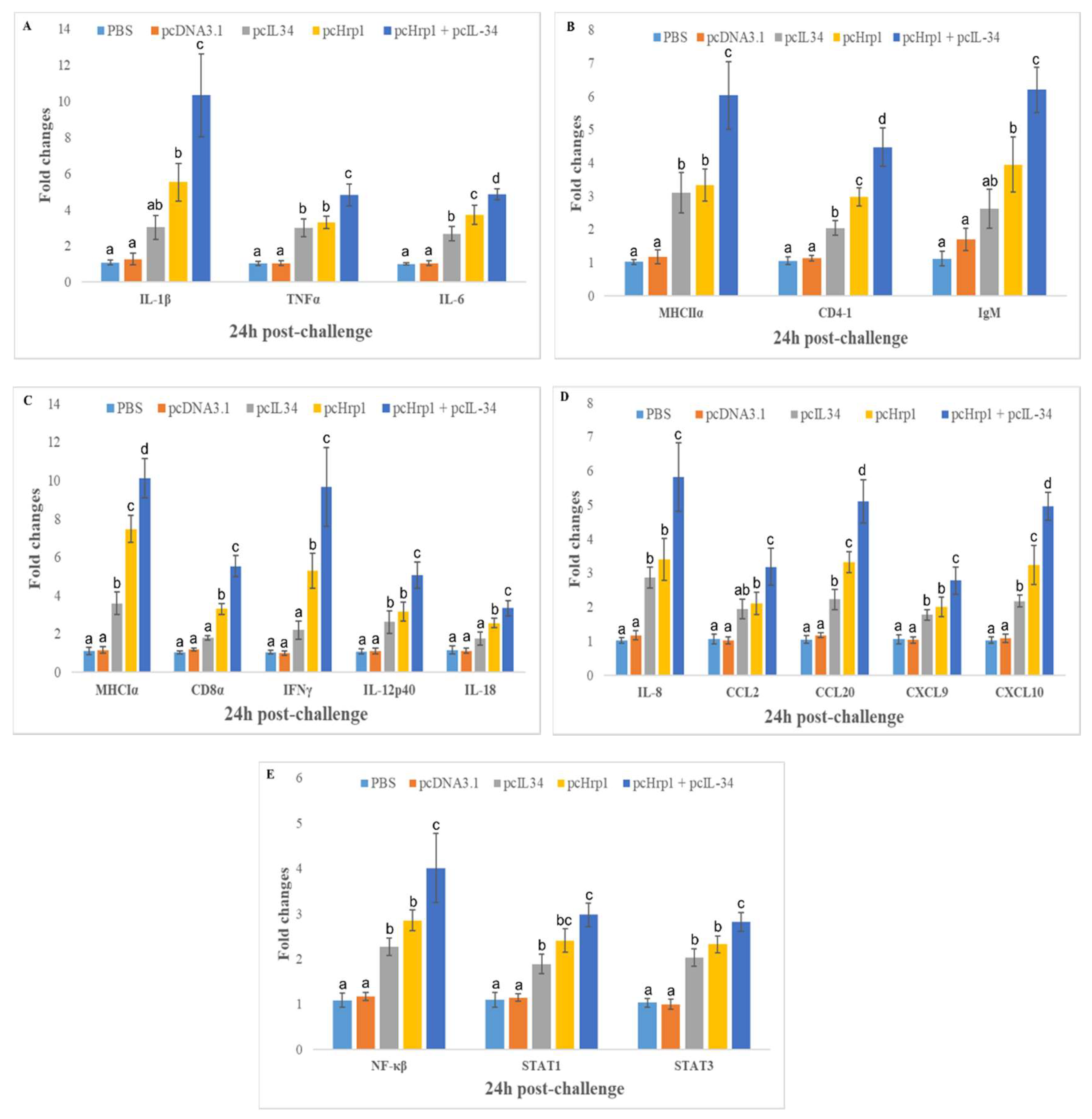
3.8. Expression of Immune-Related Genes
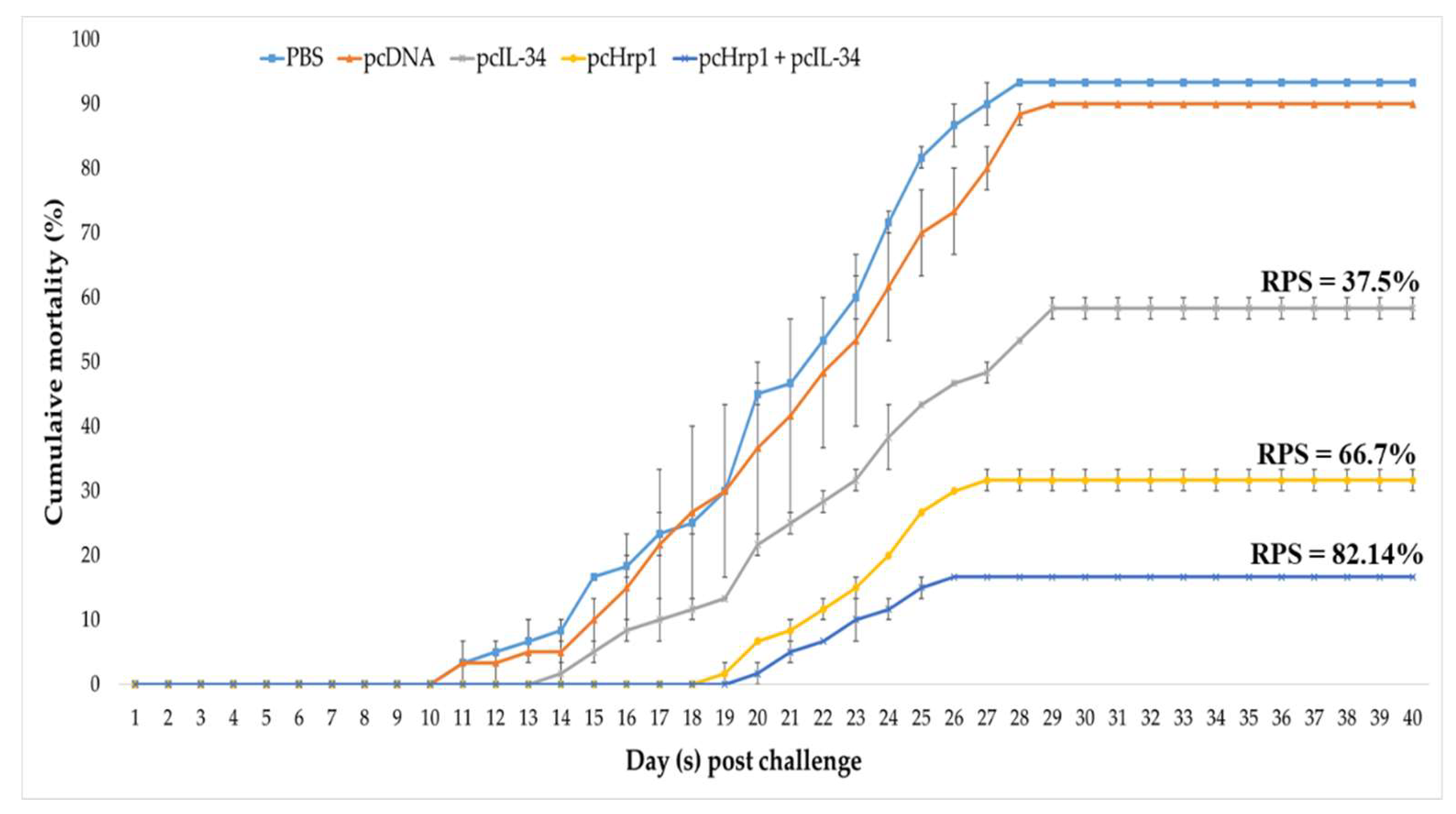
3.9. Immuno-Protective Efficacy against N. seriolae Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kariya, T.; Kubota, S.; Nakamura, Y.; Kira, K. Nocardia infection in cultured yellowtails (Seriola quinqueradiata and S. purpursacens) I. Bacteriological study. Fish Pathol. 1968, 3, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Lee, J.L.; Lai, C.C.; Gu, Y.W.; Wang, C.T.; Chang, H.Y. Nocardiosis in sea bass, Lateolabrax japonicus, in Taiwan. J. Fish Dis. 2000, 23, 299–307. [Google Scholar] [CrossRef]
- Huang, S.L.; Lai, K.C.; Su, S.C.; Shei, M.C.; Chen, S.N. Isolation and characterization of the pathogenic bacterium, Nocardia seriolae, from female broodstock of striped mullet (Mugil cephalus). J. Fish. Res. 2004, 12, 61–69. [Google Scholar]
- Labrie, L.; Ng, J.; Tan, Z.; Komar, C.; Ho, E.; Grisez, L. Nocardial infections in fish: An emerging problem in both freshwater and marine aquaculture systems in Asia. In Diseases in Asian Aquaculture VI; Reantaso, M.G.B., Mohan, V., Crumlish, M., Subasinghe, R.P., Eds.; Fish Health Section, Asian Fisheries Society: Manila, Philippines, 2008; pp. 297–312. [Google Scholar]
- Vu-Khac, H.; Duong, V.Q.B.; Chen, S.C.; Pham, T.H.; Nguyen, T.T.G.; Trinh, T.T.H. Isolation and genetic characterization of Nocardia seriolae from snubnose pompano Trachinotus blochii in Vietnam. Dis. Aquat. Org. 2016, 120, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Shimahara, Y.; Huang, Y.F.; Tsai, M.A.; Wang, P.C.; Yoshida, T.; Lee, J.L.; Chen, S.C. Genotypic and phenotypic analysis of fish pathogen, Nocardia seriolae, isolated in Taiwan. Aquaculture 2009, 294, 165–171. [Google Scholar] [CrossRef]
- Silveira, M.M.; Oliveira, L.; Schuch, R.A.; McBride, A.J.A.; Dellagostin, O.A.; Hartwig, D.D. DNA vaccines against leptospirosis: A literature review. Vaccine 2017, 35, 5559–5567. [Google Scholar] [CrossRef] [PubMed]
- McNeel, D.G.; Becker, J.T.; Johnson, L.E.; Olson, B.M. DNA Vaccines for Prostate Cancer. Curr. Cancer Ther. Rev. 2012, 8, 254–263. [Google Scholar] [CrossRef]
- Liu, C.; Hu, X.; Cao, Z.; Sun, Y.; Chen, X.; Zhang, Z. Construction and characterization of a DNA vaccine encoding the SagH against Streptococcus iniae. Fish Shellfish Immunol. 2019, 89, 71–75. [Google Scholar] [CrossRef]
- Xu, H.; Xing, J.; Tang, X.; Sheng, X.; Zhan, W. Intramuscular administration of a DNA vaccine encoding OmpK antigen induces humoral and cellular immune responses in flounder (Paralichthys olivaceus) and improves protection against Vibrio anguillarum. Fish Shellfish Immunol. 2019, 86, 618–626. [Google Scholar] [CrossRef]
- Xing, J.; Xu, H.; Tang, X.; Sheng, X.; Zhan, W. A DNA Vaccine Encoding the VAA Gene of Vibrio anguillarum Induces a Protective Immune Response in Flounder. Front. Immunol. 2019, 10, 499. [Google Scholar] [CrossRef]
- Jiao, X.D.; Zhang, M.; Hu, Y.; Sun, L. Construction and evaluation of DNA vaccines encoding Edwardsiella tarda antigens. Vaccine 2009, 27, 5195–5202. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, C.S.; Sun, L. Comparative study of the immune effect of an Edwardsiella tarda antigen in two forms: Subunit vaccine vs DNA vaccine. Vaccine 2011, 29, 2051–2057. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, J.; Zhang, H.; Liu, Q.; Xiao, J.; Zhang, Y. Design and evaluation of an Edwardsiella tarda DNA vaccine co-encoding antigenic and adjuvant peptide. Fish Shellfish Immunol. 2016, 59, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kato, G.; Kato, K.; Jirapongpairoj, W.; Kondo, H.; Hirono, I. Development of DNA vaccines against Nocardia seriolae infection in fish. Fish Pathol. 2014, 49, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, W.; Hou, S.; Fu, W.; Cai, J.; Xia, L.; Lu, Y. Comparison of protective efficacy between two DNA vaccines encoding DnaK and GroEL against fish nocardiosis. Fish Shellfish Immunol. 2019, 95, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Salonius, K.; Simard, N.; Harland, R.; Ulmer, J.B. The road to licensure of a DNA vaccine. Curr. Opin. Investig. Drugs 2007, 8, 635–641. [Google Scholar] [PubMed]
- Dalmo, R.A. DNA vaccines for fish: Review and perspectives on correlates of protection. J. Fish Dis. 2018, 41, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingolotti, M.; Kawalekar, O.; Shedlock, D.J.; Muthumani, K.; Weiner, D.B. DNA vaccines for targeting bacterial infections. Expert Rev. Vaccines 2010, 9, 747–763. [Google Scholar] [CrossRef] [Green Version]
- Reynes-Cerpa, S.; Maisey, K.; Reyes-López, F.; Toro-Ascuy, D.; Sandino, A.M.; Imarai, M. Fish cytokines and immune response. New Adv. Contrib. Fish Biol. 2012, 3–57. [Google Scholar] [CrossRef] [Green Version]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A. Adjuvants and immunostimulants in fish vaccines: Current knowledge and future perspectives. Fish Shellfish Immunol. 2013, 35, 1740–1750. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Tang, X.; Sheng, X.; Xing, J.; Zhan, W. The Immune Adjuvant Effects of Flounder (Paralichthys olivaceus) Interleukin-6 on E. tarda Subunit Vaccine OmpV. Int. J. Mol. Sci. 2017, 18, 1445. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Long, B.; Wang, K.; Wang, J.; He, Y.; Wang, X.; Yang, Q.; Liu, T.; Chen, D.; Geng, Y.; et al. Interleukin-8 holds promise to serve as a molecular adjuvant in DNA vaccination model against Streptococcus iniae infection in fish. Oncotarget 2016, 7, 83938–83950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Zhang, Q.; Xu, L.; Li, S.; Wang, D.; Zhao, J.; Liu, H.; Feng, J.; Lu, T. Effects of different cytokines on immune responses of rainbow trout in a virus DNA vaccination model. Oncotarget 2017, 8, 112222–112235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Tang, X.; Sheng, X.; Xing, J.; Zhan, W. The effects of IL-1β, IL-8, G-CSF and TNF-α as molecular adjuvant on the immune response to an E. tarda subunit vaccine in flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2018, 77, 374–384. [Google Scholar] [CrossRef]
- Xu, H.; Xing, J.; Tang, X.; Sheng, X.; Zhan, W. Generation and functional evaluation of a DNA vaccine co-expressing Vibrio anguillarum VAA protein and flounder interleukin-2. Fish Shellfish Immunol. 2019, 93, 1018–1027. [Google Scholar] [CrossRef]
- Wang, E.; Liu, T.; Wu, J.; Wang, K.; Chen, D.; Geng, Y.; Huang, X.; Ouang, P.; Lai, W.; Ai, X. Molecular characterization, phylogenetic analysis and adjuvant effect of channel catfish interleukin-1βs against Streptococcus iniae. Fish Shellfish Immunol. 2019, 87, 155–165. [Google Scholar] [CrossRef]
- Matsumoto, M.; Araki, K.; Hayashi, K.; Takeuchi, Y.; Shiozaki, K.; Suetake, H.; Yamamoto, A. Adjuvant effect of recombinant interleukin-12 in the Nocardiosis formalin-killed vaccine of the amberjack Seriola dumerili. Fish Shellfish Immunol. 2017, 67, 263–269. [Google Scholar] [CrossRef]
- Xu, H.; Xing, J.; Tang, X.; Sheng, X.; Zhan, W. The effects of CCL3, CCL4, CCL19 and CCL21 as molecular adjuvants on the immune response to VAA DNA vaccine in flounder (Paralichthys olivaceus). Dev. Comp. Immunol. 2019, 103, 103492. [Google Scholar] [CrossRef]
- Kumari, R.; Kole, S.; Soman, P.; Rathore, G.; Triphathi, G.; Makesh, M.; Rajendran, K.V.; Bedekar, M.K. Bicistronic DNA vaccine against Edwardsiella tarda infection in Labeo rohita: Construction and comparative evaluation of its protective efficacy against monocistronic DNA vaccine. Aquaculture 2018, 485, 201–209. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y.M. Immunomodulation of interleukin-34 and its potential significance as a disease Biomarker and Therapeutic target. Int. J. Biol. Sci. 2019, 15, 1835–1845. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Kono, T.; Monte, M.; Kuse, H.; Costa, M.; Korenaga, H.; Maehr, T.; Husain, M.; Sakai, M.; Secombes, C.J. Identification of IL-34 in teleost fish: Differential expression of rainbow trout IL-34, MCSF1 and MCSF2, ligands of the MCSF receptor. Mol. Immunol. 2013, 53, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, Z.Q.; Li, Y.W.; Zhou, L.; Li, A.X.; Luo, X.C.; Dan, X.M. Grouper (Epinephelus coioides) IL-34/MCSF2 and MCSFR1/MCSFR2 were involved in mononuclear phagocytes activation against Cryptocaryon irritans infection. Fish Shellfish Immunol. 2015, 67, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, L.; Wu, C.; Lou, B. Molecular characterization and expression analysis of large yellow croaker (Larimichthys crocea) interleukin-12A, 16 and 34 after poly I:C and Vibrio anguillarum challenge. Fish Shellfish Immunol. 2018, 74, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Jiang, X.; Gao, J.; Li, X.; Xu, J.; Wang, J.; Gao, Q.; Zou, J. Functional characterisation of interleukin 34 in grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2019, 92, 91–100. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Suzuki, S. A PCR method to detect Nocardia seriolae in fish samples. Fish Pathol. 2003, 38, 93–97. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Heijne von, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Pitti, T.; Chen, C.T.; Lin, H.N.; Choong, W.K.; Hsu, W.L.; Sung, T.Y. N-GlyDE: A two-stage N-linked glycosylation site prediction incorporating gapped dipeptides and pattern-based encoding. Sci. Rep. 2019, 9, 15975. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; McGinnis, S.; Madden, T.L. BLAST: Improvements for better sequence analysis. Nucleic Acids Res. 2006, 34, W6–W9. [Google Scholar] [CrossRef] [Green Version]
- Kozak, M. Structural features in eukaryotic mRNAs that modulate the initiation of translation. Biol. Chem. 1991, 266, 19867–19870. [Google Scholar]
- Hoang, H.H.; Wang, P.C.; Chen, S.C. The protective efficacy of recombinant hypoxic response protein 1 of Nocardia seriolae in largemouth bass (Micropterus salmoides). Vaccine 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.T.; Nguyen, H.T.; Wang, P.C.; Chen, S.C. Identification and expression analysis of two pro-inflammatory cytokines, TNF-alpha and IL-8, in cobia (Rachycentron canadum L.) in response to Streptococcus dysgalactiae infection. Fish Shellfish Immunol. 2017, 67, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Byadgi, O.; Chen, C.W.; Wang, P.C.; Tsai, M.A.; Chen, S.C. De Novo Transcriptome Analysis of Differential Functional Gene Expression in Largemouth Bass (Micropterus salmoides) after Challenge with Nocardia seriolae. Int. J. Mol. Sci. 2016, 17, 1315. [Google Scholar] [CrossRef] [Green Version]
- Panzarin, V.; Toffan, A.; Abbadi, M.; Buratin, A.; Mancin, M.; Braaen, S.; Olsen, C.M.; Bargelloni, L.; Rimstad, E.; Cattoli, G. Molecular Basis for Antigenic Diversity of Genus Betanodavirus. PLoS ONE 2016, 11, e0158814. [Google Scholar] [CrossRef]
- Huang, Y.; Cai, S.; Pang, H.; Jian, J.; Wu, Z. Immunogenicity and efficacy of DNA vaccine encoding antigenic AcfA via addition of the molecular adjuvant Myd88 against Vibrio alginolyticus in Epinephelus coioides. Fish Shellfish Immunol. 2017, 66, 71–77. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, J.; Liu, R.; Jia, J. Identification and evaluation of an outer membrane protein OmpU from a pathogenic Vibrio harveyi isolate as vaccine candidate in turbot (Scophthalmus maximus). Lett. Appl. Microbiol. 2011, 53, 22–29. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, J.; Liu, M.; Ren, G.; Jian, F.; Yin, J.; Feng, J.; Liu, H.; Lu, T. Bivalent DNA vaccine induces significant immune responses against infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus in rainbow trout. Sci. Rep. 2017, 7, 5700. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Nguyen, T.T.T.; Chen, Y.C.; Vu-Khac, H.; Wang, P.C.; Chen, S.C. Enhanced immune responses and effectiveness of refined outer membrane protein vaccines against Vibrio harveyi in orange-spotted grouper (Epinephelus coioides). J. Fish Dis. 2018, 41, 1349–1358. [Google Scholar] [CrossRef]
- Ellis, A.E. Lysozyme Assays. InTech. Fish Immunol. 1990, 1, 101–103. [Google Scholar]
- Martyniuk, C.J.; Doperalski, N.J.; Prucha, M.S.; Zhang, J.L.; Kroll, K.J.; Conrow, R.; Barber, D.S.; Denslow, N.D. High contaminant loads in Lake Apopka’s riparian wetland disrupt gene networks involved in reproduction and immune function in largemouth bass. Comp. Biochem. Physiol. D Genom. Proteom. 2016, 19, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Baghdadi, M.; Umeyama, Y.; Hama, N.; Kobayashi, T.; Han, N.; Wada, H.; Seino, K.I. Interleukin-34, a comprehensive review. J. Leukoc. Biol. 2018, 104, 931–951. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Leo, C.; Chen, X.; Wong, B.R.; Williams, L.T.; Lin, H.; He, X. The mechanism of shared but distinct CSF-1R signaling by the non-homologous cytokines IL-34 and CSF-1. Biochim. Biophys. Acta 2012, 1824, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Hølvold, L.B.; Myhr, A.I.; Dalmo, R.A. Strategies and hurdles using DNA vaccines to fish. Vet. Res. 2014, 45, 21. [Google Scholar] [CrossRef] [Green Version]
- Roupie, V.; Romano, M.; Zhang, L.; Korf, H.; Lin, M.Y.; Franken, K.L.M.C. Immunogenicity of eight dormancy regulon-encoded proteins of Mycobacterium tuberculosis in DNA-vaccinated and tuberculosis-infected mice. Infect. Immun. 2007, 75, 941–949. [Google Scholar] [CrossRef] [Green Version]
- Bashir, N.; Kounsar, F.; Mukhopadhyay, S.; Hasnain, S.E. Mycobacterium tuberculosis conserved hypothetical protein rRv2626c modulates macrophage effector functions. Immunology 2010, 130, 34–45. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, M.; Chaudhry, A.; Sharma, S. Rv2626c and Rv2032 activate TH1 response and downregulate regulatory T cells in peripheral blood mononuclear cells of tuberculosis patients. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 46–53. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Falk, K.; Weli, S.C.; Koppang, E.O.; Kvellestad, A. A sequential study of incomplete Freund’s adjuvant-induced peritonitis in Atlantic cod. Fish Shellfish Immunol. 2012, 32, 141–150. [Google Scholar] [CrossRef]
- Veenstra, K.A.; Wang, T.; Alnabulsi, A.; Douglas, A.; Russell, K.S.; Tubbs, L.; Arous, J.B.; Secombes, C.J. Analysis of adipose tissue immune gene expression after vaccination of rainbow trout with adjuvanted bacterins reveals an association with side effects. Mol. Immunol. 2017, 88, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Jiao, C.; Bao, P.; Liu, Q.; Wang, P.; Zhang, R.; Liu, X.; Zhang, Y. Efficacy of MontanideTM ISA 763 A VG as aquatic adjuvant administrated with an inactivated Vibrio harveyi vaccine in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2019, 84, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jin, P.; Liu, Q.; Wang, Q.; Zhang, Y.; Liu, X. A CpG-riched plasmids as vaccine adjuvant reduce antigen dose of an inactivated Vibrio anguillarum vaccine in turbot (Scophthalamus maximus L.). Fish Shellfish Immunol. 2020, 98, 312–317. [Google Scholar] [CrossRef]
- Zou, J.; Carrington, A.; Collet, B.; Dijkstra, J.M.; Yoshiura, Y.; Bols, N.; Secombes, C. Identification and bioactivities of IFN-γ in rainbow trout Oncorhynchus mykiss: The first Th1-type cytokine characterized functionally in fish. J. Immunol. 2005, 175, 2484–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonheim, T.C.; Bøgwald, J.; Dalmo, R.A. What happens to the DNA vaccine in fish? A review of current knowledge. Fish Shellfish Immunol. 2008, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Z.; Wang, W.; Hou, S.; Cai, J.; Xia, L.; Lu, Y. Development of DNA vaccines encoding ribosomal proteins (RplL and RpsA) against Nocardia seriolae infection in fish. Fish Shellfish Immunol. 2020, 96, 201–212. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, Y.H.; Liu, C.S.; Sun, L. Construction and comparative study of monovalent and multivalent DNA vaccines against Streptococcus iniae. Fish Shellfish Immunol. 2012, 33, 1303–1310. [Google Scholar] [CrossRef]
- Lee, L.Y.Y.; Izzard, L.; Hurt, A.C. A Review of DNA Vaccines Against Influenza. Front. Immunol. 2018, 9, 1568. [Google Scholar] [CrossRef] [Green Version]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights to the function of lysozyme. PLoS Pathog. 2017, 21, e1006512. [Google Scholar] [CrossRef]
- Brott, A.S.; Clarke, A.J. Peptidoglycan O-Acetylation as a Virulence Factor: Its Effect on Lysozyme in the Innate Immune System. Antibiotics (Basel) 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Tang, X.; Liu, F.; Sheng, X.; Xing, J.; Zhan, W. Recombinant NADP-dependent isocitrate dehydrogenase of Edwardsiella tarda induces both Th1 and Th2 type immune responses and evokes protective efficacy against edwardsiellosis. Vaccine 2018, 36, 2337–2345. [Google Scholar] [CrossRef] [PubMed]
- Prados-Rosales, R.; Carreño, L.; Cheng, T.; Blanc, C.; Weinrick, B.; Malek, A.; Lowary, T.; Baena, A.; Joe, M.; Bai, Y.; et al. Enhanced control of Mycobacterium tuberculosis extrapulmonary dissermination in mice by an arabinomannan-protein conjugate vaccine. PLoS Pathog. 2017, 13, e1006250. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.E.; Burtnick, M.N.; Stokes, M.G.; Whelan, A.O.; Williamson, E.D.; Atkins, T.P.; Prior, J.L.; Brett, P.J. Burkholderia pseudomallei capsular polysaccharide conjugates provide protection against acute melioidosis. Infect. Immun. 2014, 82, 3206–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawool, D.B.; Bitsaktsis, C.; Li, Y.; Gosselin, D.R.; Lin, Y.; Kurkure, N.V.; Metzger, D.W.; Gosselin, E.J. Utilization of Fc receptors as a mucosal vaccine strategy against an intracellular bacterium, Francisella tularensis. J. Immunol. 2008, 180, 5548–5557. [Google Scholar] [CrossRef]
- Singh, A.K.; Kingston, J.J.; Murali, H.S.; Batra, H.V. A recombinant bivalent fusion protein rVE confers active and passive protection against Yersinia enterocolitica infection in mice. Vaccine 2014, 32, 1233–1239. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.; Olano, J.P.; Feng, H.M.; Walker, D.H. Current status of immune mechanism of killing of intracellular microorganisms. FEMS Microbiol. Lett. 2002, 207, 111–120. [Google Scholar] [CrossRef]
- Yung, S.C.; Murphy, P.M. Antimicrobial chemokines. Front. Immunol. 2012, 3, 276. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.N.; Teixeira-Cunha, M.G.; Figueiredo, R.T.; Almeida, P.E.; Alves, S.C.; Bozza, P.T.; Bozza, F.A.; Bozza, M.T.; Zimmerman, G.A.; Castro-Faria-Neto, H.C. Bacterial clearance in septic mice is modulated by MCP-1/CCL2 and nitric oxide. Shock 2013, 39, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Depaolo, R.W.; Lathan, R.; Rollins, B.J.; Karpus, W.J. The chemokine CCL2 is required for control of murine gastric Salmonella enterica infection. Infect. Immun. 2005, 73, 6514–6522. [Google Scholar] [CrossRef] [Green Version]
- Collin, M.; Linge, H.M.; Bjartell, A.; Giwercman, A.; Malm, J.; Egesten, A. Constitutive expression of the antibacterial CXC chemokine GCP-2/CXCL6 by epithelial cells of male reproductive tract. J. Reprod. Immunol. 2008, 79, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Valdivia-Silva, J.; Medina-Tayamo, J.; Gracia-Zepeda, E.A. Chemokine-derived peptides: Novel antimicrobial and antineoplastic agents. Int. J. Mol. Sci. 2015, 16, 12958–12985. [Google Scholar] [CrossRef] [PubMed]
- Söbirk, S.K.; Mörgelin, M.; Egesten, A.; Bates, P.; Shannon, O.; Collin, M. Human chemokines as antimicrobial peptides with direct parasiticidal effect on Leishmania mexicana in vitro. PLoS ONE 2013, 8, e58129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannidis, L.J.; Nie, C.Q.; Hansen, D.S. The role of chemokines in severe malaria: More than meets the eye. Parasitology 2014, 141, 602–613. [Google Scholar] [CrossRef]
- Domingo-Gonzalez, R.; Prince, O.; Cooper, A.; Khader, S.A. Cytokines and Chemokines in Mycobacterium tuberculosis Infection. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.P.; Wu, X.S.; Xie, Y.Y.; Dai, B.B.; Hu, W.; Ge, J.F.; Chen, F.H. Functions of interleukin-34 and its emerging association with rheumatoid arthritis. Immunology 2016, 149, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Truong, A.D.; Hong, Y.; Lee, J.; Lee, K.; Kil, D.Y.; Lillehoj, H.S.; Hong, Y.H. Interleukin-34 Regulates Th1 and Th17 Cytokine Production by Activating Multiple Signaling Pathways through CSF-1R in Chicken Cell Lines. Int. J. Mol. Sci. 2018, 19, 1665. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Luo, H.; Yan, X.; Song, Z.; Gao, X.; Xia, Y.; Zhang, L.; Yin, Y.; Cao, J. Interleukin-34 ameliorates survival and bacterial clearance in polymicrobial sepsis. Crit. Care Med. 2018, 46, e584–e590. [Google Scholar] [CrossRef]
- Mikalsen, A.B.; Torgersen, J.; Aleström, P.; Hellemann, A.L.; Koppang, E.O.; Rimstad, E. Protection of atlantic salmon Salmo salar against infectious pancreatic necrosis after DNA vaccination. Dis. Aquat. Organ. 2004, 60, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Kanellos, T.; Sylvester, I.D.; D’Mello, F.; Howard, C.R.; Mackie, A.; Dixon, P.F.; Chang, K.C.; Ramstad, A.; Midtlyng, P.J.; Russel, P.H. DNA Vaccination Can Protect Cyprinus Carpio Against Spring Viraemia of Carp Virus. Vaccine 2006, 24, 4927–4933. [Google Scholar] [CrossRef]
- Caipang, C.M.; Takano, T.; Hirono, I.; Aoki, T. Genetic Vaccines Protect Red Seabream, Pagrus Major, Upon Challenge With Red Seabream Iridovirus (RSIV). Fish Shellfish Immunol. 2006, 21, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Pasnik, D.J.; Smith, S.A. Immune and histopathologic responses of DNA-vaccinated hybrid striped bass Morone Saxatilis X M. Chrysops after acute Mycobacterium marinum infection. Dis. Aquat. Organ. 2006, 73, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Tan, W.; Wang, W.; Hou, S.; Chen, G.; Xia, L.; Lu, Y. Identification of common antigens of three pathogenic Nocardia species and development of DNA vaccine against fish nocardiosis. Fish Shellfish Immunol. 2019, 95, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Evensen, Ø. Correlates of protective immunity of fish vaccines. Fish Shellfish Immunol. 2019, 85, 132–140. [Google Scholar] [CrossRef]
- Cai, S.H.; Huang, Y.C.; Lu, Y.S.; Wu, Z.H.; Wang, B.; Tang, J.F.; Jian, J.C. Expression and immunogenicity analysis of accessory colonization factor A from Vibrio alginolyticus strain HY9901. Fish Shellfish Immunol. 2013, 43, 454–462. [Google Scholar] [CrossRef]
- Xing, J.; Xu, H.; Wang, Y.; Tang, X.; Sheng, X.; Zhan, W. Protective efficacy of six immunogenic recombinant proteins of Vibrio anguillarum and evaluation them as vaccine candidate for flounder (Paralichthys olivaceus). Microb. Pathog. 2017, 107, 155–163. [Google Scholar] [CrossRef]
- Xing, J.; Xu, H.; Wang, Y.; Tang, X.; Sheng, X.; Zhan, W. Identification of immunogenic proteins and evaluation of four recombinant proteins as potential vaccine antigens from Vibrio anguillarum in flounder (Paralichthys olivaceus). Vaccine 2017, 35, 3196–3203. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ → 3′) | Application | References |
|---|---|---|---|
| Hrp1-F | TTGCTAGCGCCACCATGGCCACGGCACG (NheI) | Plasmid construction, PCR, and RT-PCR | This study |
| Hrp1-R | GTAAGCTTCTAATGATGATGATGATGATGTGTCCAAGGCGCGCA (HindIII) | Plasmid construction | |
| Hrp1-R2 | CTATGTCCA AGGCGCGCAGACC | PCR and RT-PCR | |
| IL-34-F | CGAAGCTTGCCACCATGGTTCTACTGTGCA (HindIII) | Plasmid construction, PCR, and RT-PCR | |
| IL-34-R | GAATTCTCAATGATGATGATGATGATGGCTTTCGGGCTGTAA (EcoRI) | Plasmid construction | |
| IL-34-R2 | TCAGCTTTCGGGCTGTAAGTCTTTC | PCR and RT-PCR | |
| RPS40 | F: CAGAAATGGCACGATAAGCA | qRT-PCR | Martyniuk et al., 2016 |
| R: GACCTTTACGCCCAAATCC | |||
| β-actin | F: CCACCACAGCCGAGAGGGAA | qRT-PCR | Omkar et al, 2016 |
| R: TCATGGTGGATGGGGCCAGG | |||
| IL-1β | F: TTGCCATAGAGAGGTTTA | qRT-PCR | |
| R: ACACTATATGCTCTTCCA | |||
| IL-12p40 | F: TCTTCCATCCTTGTGGTCTTCC | qRT-PCR | |
| R: CAGTTCCAGGTCAAAGTGGTC | |||
| TNFα | F: CTAGTGAAGAACCAGATTGT | qRT-PCR | |
| R: AGGAGACTCTGAACGATG | |||
| IL-8 | F: GAGCCATTTTTCCTGGTGACT | qRT-PCR | |
| R: TCCTCATTGGTGCTGAAAGATC | |||
| NF-κB | F: AGGATGACTGAAGCTCCGTT | qRT-PCR | |
| R: GGACACGAGGAGGATCGGAGT | |||
| IFNγ | F: TGCAGGCTCTCAAACACATC | qRT-PCR | Hoang et al, 2020 |
| R: TGTTTTCGGTCAGTGTGCTC | |||
| MHCI-α | F: GTGGTTCAACGTCAACATCG | qRT-PCR | |
| R: ACCCAGACTTGTTCGGTGTC | |||
| MHCII-α | F: GAGGACCTTGCTGTCATTGG | qRT-PCR | |
| R: GCGTACCAAACCTCTTCACC | |||
| CD4-1 | F: GCTCCAGCGGGGAATAATTT | qRT-PCR | |
| R: GCCAGGCAAGCTCAAAGTTA | |||
| CD8-α | F: GGAAGGGGATCCTGTTGACA | qRT-PCR | |
| R: CCAGCACTCGAAACCAGATG | |||
| IgM | F: CTGGACCAGTCTCCCTCTGA | qRT-PCR | |
| R: CGAGGTACTGAGTGCTGCTG | |||
| STAT3 | F: CCACCCAAAGAACGTGAACT | qRT-PCR | |
| R: TCAATGGTCAGGCCTCTCTT | |||
| CCL20 | F: ACAACCACGGAAAACTGCCG | qRT-PCR | |
| R: TCCTCACCCACTCATCCTTC | |||
| STAT1 | F: TAAAACTCCGGTTCCTGGTG | qRT-PCR | This study |
| R: CCGTTTGACTCCTCCATGTT | |||
| IL-18 | F: TTGATGGCAAGAAGATGGTGG | qRT-PCR | |
| R: AAGCCTTGTGTGCAGTTTCCT | |||
| IL-6 | F: GGAACCCTGAACAGGTAACG | qRT-PCR | |
| R: TGTGCGGTCATCTTTCTGTGG | |||
| CCL2 | F: GCGAGTGGTCAGCTACATCA | qRT-PCR | |
| R: GATGAGCTCCTTCACCCAAG | |||
| CXCL9 | F: GGAAGATGTTTGTGTCCACAG | qRT-PCR | |
| R: GGCGTTTTGGGTAGACTGTG | |||
| CXCL10 | F: GAATCGGGACAGCAGTGTCT | qRT-PCR | |
| R: CAGTTGCTGGGTAGATCTGGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, H.H.; Wang, P.-C.; Chen, S.-C. Interleukin 34 Serves as a Novel Molecular Adjuvant against Nocardia Seriolae Infection in Largemouth Bass (Micropterus Salmoides). Vaccines 2020, 8, 151. https://doi.org/10.3390/vaccines8020151
Hoang HH, Wang P-C, Chen S-C. Interleukin 34 Serves as a Novel Molecular Adjuvant against Nocardia Seriolae Infection in Largemouth Bass (Micropterus Salmoides). Vaccines. 2020; 8(2):151. https://doi.org/10.3390/vaccines8020151
Chicago/Turabian StyleHoang, Huy Hoa, Pei-Chi Wang, and Shih-Chu Chen. 2020. "Interleukin 34 Serves as a Novel Molecular Adjuvant against Nocardia Seriolae Infection in Largemouth Bass (Micropterus Salmoides)" Vaccines 8, no. 2: 151. https://doi.org/10.3390/vaccines8020151