1. Introduction
The
Drosophila melanogaster innate immune system shares several characteristics with its mammalian counterparts, including origin, type, and timeframe of the immune response [
1]. Of the genes associated with the innate immune response in
D. melanogaster, 77% are shared with humans [
2]. For this reason,
D. melanogaster provide an easily manipulated model for many immune pathways and offer novel target proteins for study in disease models. One group of mammalian proteins, the Toll-like receptor family, was first identified in response to a
Drosophila protein, Toll, involved in activation of the Nuclear factor-Kappa B (NF-κB) pathway [
3]. Further, the NF-κB pathway in
Drosophila continues to be studied to better understand this pathway in mammalian models. In
D. melanogaster, innate immune responses include autophagy, RNA interference (RNAi), and several pathways including the Toll, immune deficiency (IMD), and Janus kinase/signal transducers and activators of transcription (Jak-STAT) pathways [
4]. Autophagy is a cytoplasmic degradation process, involved in starvation and stress responses, development, cell death, and immune responses.
Drosophila autophagy relies upon the stimulation of pattern-recognition receptors inducing antimicrobial peptide production and activation of the Toll and IMD pathways [
5]. RNAi inhibits viral replication by fragmenting the viral genome and using these fragments, small interfering RNAs (siRNAs) to target the viral genome [
6]. The siRNAs are loaded into the RNA-induced silencing complex (RISC) and serve as a guide indicating which sequences to cleave. The RNAi response leads to production of tumor necrosis factor receptor-associated factor (TRAF) [
7]. The IMD pathway is implicated in the recognition of viral particles by peptidoglycan recognition proteins (PGRP-LC and PGRP-LE) and leads to the cleavage of the NF-κB transcription factor, Relish (Rel) by TRAF. The cleaved Relish promotes transcription of IMD-regulated genes, including
vago [
8]. Vago, a secretory, cytokine-like molecule serves to activate the Jak-STAT pathway in conjunction with Unpaired (Upd), and the Domeless (Dome) receptor [
8,
9]. Activation of the Jak-STAT pathway regulates the expression of several genes involved in modulating viral load, including
vir-1 that encodes virus-induced RNA 1 (Vir-1). Vir-1 has been implicated in response to
Drosophila C virus (DCV) and Flock House virus (FHV), and in a potential cold tolerance role [
4,
10,
11]. Upregulated genes in DCV-infected
D. melanogaster included
vago and
vir-1 with 45-fold induction of
vago and 46-fold induction of
vir-1 [
4].
These immune pathways are well understood in response to DCV, but not in response to Nora virus infection. In addition, Vir-1 and Vago response to Nora virus has not been characterized. Nora virus is a small, picorna-like virus that establishes a persistent infection in
D. melanogaster.
D. melanogaster Nora virus consists of four open reading frames (ORFs) [
12].
ORF1 encodes an RNA interference (RNAi) suppressor that possibly contributes to the establishment of the aforementioned persistent infection seen in
D. melanogaster [
13].
ORF2 encodes a picorna-like replicative cassette, which consists of a helicase, protease, and RNA-dependent RNA polymerase. The capsid proteins are encoded in
ORF3 (viral protein 3; VP3) and
ORF4 (VP4 that is processed into VP4A, B, and C) [
14]. As an enteric virus, it replicates primarily in the gut tissue, is localized in the intestines, and is transmitted horizontally [
13]. Our lab has documented a locomotor defect with persistent Nora virus infection. Infected
D. melanogaster are significantly hindered in terms of geotaxis, when compared to their uninfected counterparts [
15]. To explain this phenotype, it is proposed that Nora virus-specific molecules,
ORF1 and VP4B, are circulating in the hemolymph to the brain. Hemocytes can process viral genetic material to produce virus-derived small interfering RNAs (vsRNAs) [
16]. These vsRNAs are packaged into exosomes and taken up by surrounding cells, conferring RNAi immunity. In addition, we hypothesize that both Vago and Vir-1 are being produced in response to Nora virus infection and circulating in the hemolymph. To address the question of whether Vago and Vir-1 are produced in response to Nora virus infection, we analyzed whole fly lysates. In addition, to determine if these molecules are circulating, the hemolymph of Nora virus-infected and uninfected
D. melanogaster was analyzed for presence of Vago, Vir-1, and Nora virus capsid protein (VP4B), as well as Nora virus transcript levels (
ORF1). Lastly, to determine if these levels change as the fly ages, a time course analysis for these molecules was performed, using both hemolymph and whole flies. We predict that if Nora virus is circulating in the hemolymph, Vago and Vir-1 will be present to regulate viral load in Nora virus-infected
D. melanogaster.
4. Discussion
The current study provides evidence that Nora virus may be circulating in the hemolymph and inducing two candidate immune genes,
vir-1 and
vago. Nora virus was previously identified primarily within the gut tissue of infected
D. melanogaster [
13]. After identifying a locomotor defect in infected
D. melanogaster [
15], it was hypothesized that Nora virus may circulate to other tissues by way of the hemolymph. Nora virus RNA for the
ORF1 gene and capsid protein, VP4b, were found to be present in the hemolymph of infected
D. melanogaster both by Western blot and qRT-PCR (
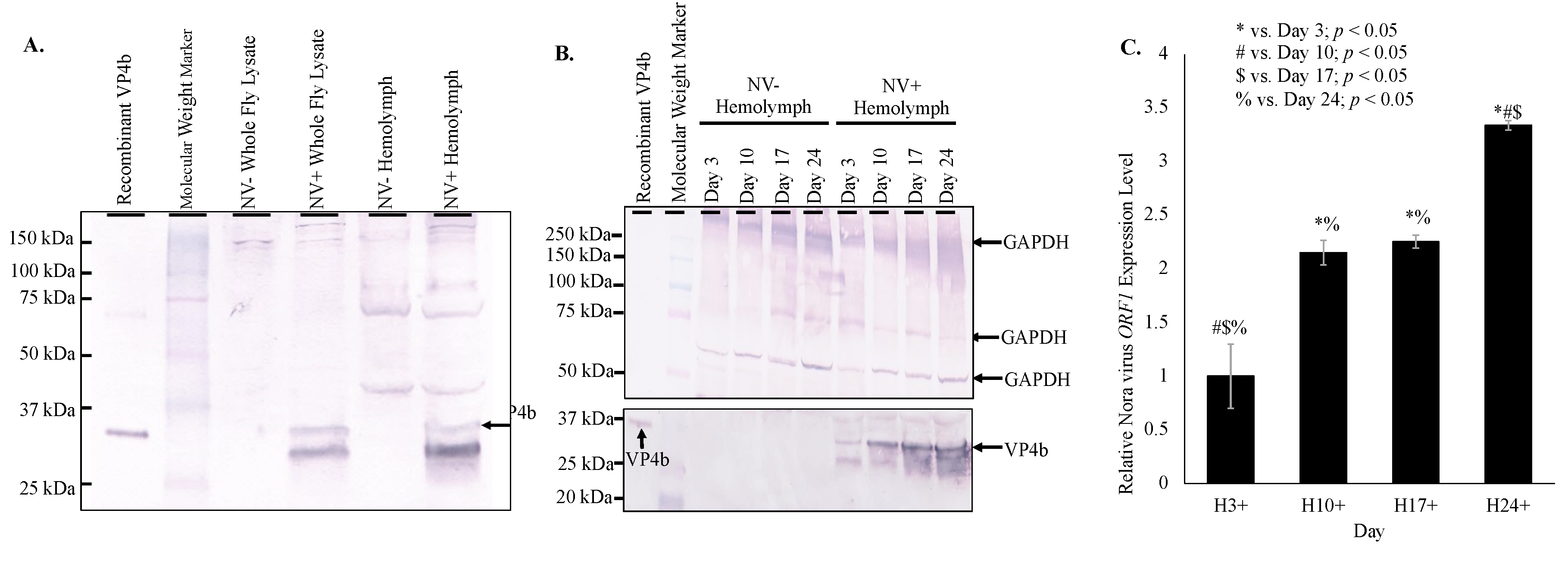
Figure 1). Presence of VP4b in the hemolymph of infected
D. melanogaster indicates that viral capsid components are present in the hemolymph (
Figure 1A), while qRT-PCR amplification of Nora virus
ORF1 in hemolymph samples (
Figure 1C) supports the possibility of full virions circulating in the hemolymph. The lower concentration of
ORF1 transcript levels in the hemolymph compared to whole fly lysates is likely due to two factors. The first is that the site of Nora virus replication is in the midgut of the organism [
12], which is where the highest levels of
ORF1 transcripts would be expected to be found. Second, the hemocytes in the hemolymph act similar to mammalian monocytes/macrophages in that they phagocytose and encapsulate invading pathogens, such as a virus [
21]. Therefore, the lower viral titer in the hemolymph could be due to viral clearance by these cells. Viral circulation in the hemolymph of
D. melanogaster has not previously been demonstrated. However, dissemination of arboviruses into the hemolymph is well-documented in mosquito vectors [
22,
23]. One study found that infecting viral particles may cross the midgut basal lamina into the mosquito hemocoel without replicating within the midgut tissue [
23].
Overall, this data supports the possibility of Nora virus circulating in the hemolymph from the gut to the brain, contributing to the locomotor defect observed in infected
D. melanogaster. Viral circulation through
Drosophila hemolymph is not yet well studied. However, the proposed circulation of Nora virus would require the virus to overcome several obstacles. First, Nora virus must be able to cross the barrier of the gut lamina. Even after crossing the gut lamina, the virus must remain infectious and evade destruction by circulating hemocytes. Lastly, the virus must be capable of entering a secondary site and establishing an infection. Additional study of Nora virus circulation is warranted, investigating the infection of secondary tissues. The ability of Nora virus to invade and infect the brain and muscle tissue should be investigated to determine which secondary sites of infection might contribute to the motor defect phenotype. Other picorna-like viruses have been identified, which infect the nervous tissues of various insects, including the novel Kakugo virus, which infects the brain tissue of worker honeybees [
24].
Nora virus is a novel RNA virus that appears to share many features with other RNA viruses. One of these features is the biphasic model of viral load (
Figure 1C and
Figure 2), which is seen in other picornaviruses, such as enterovirus. For example, the load of coxsackievirus B3 in infected mice also follows a biphasic model [
25]. In this study, it was found that Nora virus shows a biphasic model of viral load in both whole fly lysates and in the hemolymph (
Figure 1C and
Figure 2). This suggests an initial period of rapid replication in the first 3 days of infection, quickly opposed by host production of antiviral proteins. As the immune response brings the viral load under control around Day 10 (
Figure 1C and
Figure 2), transcription of antiviral proteins decreased (
Figure 3B). This allows the remaining virus to resume rapid replication, resulting in the late-life peak in viral load near day 24 (
Figure 1C and
Figure 2). Nora virus VP1 proteins have been shown to inhibit the RNAi pathway in vitro [
26]. This inhibition or other unknown mechanisms may allow Nora virus to suppress the host anti-viral response.
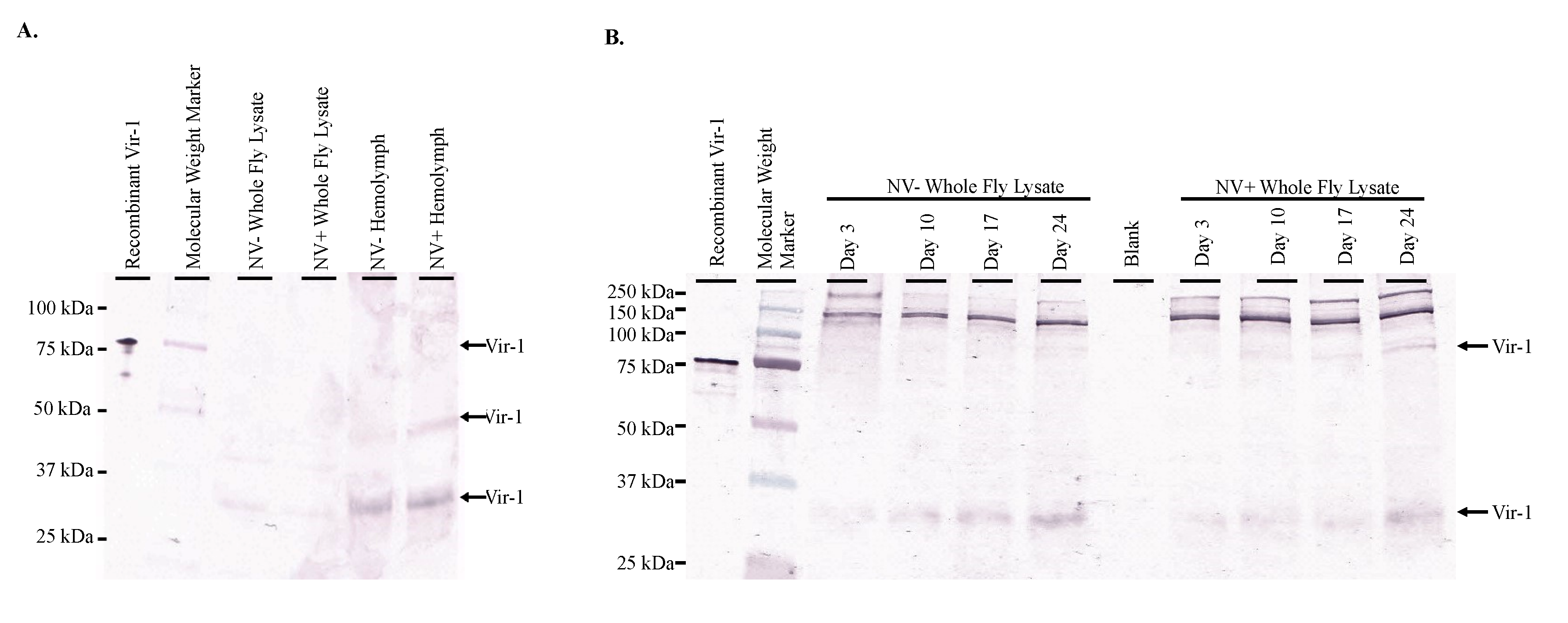
The role of candidate immune system genes in response to Nora virus infection is of interest when determining how the virus is regulated. In previous studies, two genes upregulated by Nora virus,
vago and
vir-1, were identified [
17,
18], but not characterized. In one study, microarray analysis of Nora virus-infected
D. melanogaster identified 46 upregulated and 12 downregulated genes. The study further investigated two of these genes,
Chorion protein 16 and
Troponin C isoform 4, but left the remaining identified genes, including
vago, for future analysis [
17]. A later study used next generation sequencing to identify 35 upregulated immune response genes in Nora virus-infected
D. melanogaster including
vir-1 [
18]. In the current study, the production of Vago and Vir-1 were determined in response to Nora virus infection using a time course analysis, qRT-PCR and Western blot analyses. The data here show Vago was present (
Figure 3A) and
vago transcription was upregulated (
Figure 3B,C) in response to Nora virus infection. Additionally,
vago transcription was increased in the hemolymph of infected
D. melanogaster (
Figure 3B), following the viral load (
Figure 1C). This suggests that Vago plays a role in the
D. melanogaster immune response to Nora virus. This expression of Vago in response to viral infection was previously reported with DCV infection at day 4 post-infection. In addition, flies with mutant
vago that were infected with DCV contained higher levels of virus than wild-type controls [
9]. Therefore, the decreased level of virus in the hemolymph as compared to the whole body samples could be due to the increased expression of
vago. Another study determined that Nora virus load was not impacted by
D. melanogaster null mutations in RNAi components:
r2d2, AGO2, or
Dicer-2. It is proposed that Nora virus either disables the RNAi pathway or that replication occurs in a compartment inaccessible to the RNAi machinery, allowing the RNAi resistant replication of Nora virus [
27]. The RNAi machinery is necessary for transcription of
vago, as Relish requires TRAF-mediated dephosphorylation [
8]. The increasing
vago transcription demonstrated in this study suggests that the RNAi pathway is still functional in Nora virus-infected
D. melanogaster and it is unlikely that Nora virus produces an RNAi inhibitor. This also supports the proposed replication compartment model, where the RNAi pathway continues to operate, promoting the transcription of
vago, while allowing Nora virus replicates in an isolated compartment evading the effects of the RNAi machinery [
13].
The role of Vago during viral infection is not well characterized and the data reported here is the first characterization of it during Nora virus infection. As previously mentioned, viral infection triggers expression of a number of genes, including
vago, which encodes an 18 kDa polypeptide that may function either as an antiviral molecule targeting virions or as a cytokine stimulating an interferon-like response. The mechanism of how Vago is involved in establishing an immune response has been hypothesized as involving RNAi and JAK-STAT pathways. During infection with DCV,
vago induction was dependent on Dicer-2, specifically the DExD/H-box helicase domain. This domain may act as a sensor that can induce
vago transcription for an additional antiviral response. Further evaluation has shown similarities of the DExD/H-box helicase domain and the products of the RLR helicases of retinoic acid-inducible gene I (RIG-1), melanoma differentiation-associated protein 5 (MDA5), and laboratory of genetics and physiology 2 (LGP2) in mammals to sense viral RNA [
9]. In mammals, these helicases respond to viral RNA by inducing interferon and secretions of interferons to stimulate antiviral activity in other cells, which provides further evidence for the role of Vago functioning as a cytokine. Another study evaluated the role of Vago during infection of
Culex mosquitoes with the West Nile virus (WNV). During WNV, infection leads to secretion of Vago via Dicer-2 mediated stimulation of TRAF and Rel2, which leads to Rel2 inducing
vago transcription. Once Vago is secreted, it activates the JAK-STAT pathway in neighboring cells to stimulate an antiviral response. This study found a proposed mechanism that involves Vago induction via Dicer-2 to stimulate an antiviral response via the JAK-STAT pathway. Again, this proposed pathway is similar to the RIG-1/TRAF/NF-kB interferon pathway in mammals [
7]. However, NF-kB mediated induction of Vago in
Drosophila was unable to be well studied, which suggests additional investigation to determine if the proposed mechanism in
Culex is similar or an alternative mechanism exists for
Drosophila.
As a proposed mechanism for Vago was developed, the interaction between Vago and vir-1 was further studied. Vir-1 has been implicated in antiviral defense, but its involvement is unclear and this is the first report of the Vir-1 response to Nora virus. During DCV infection,
vago-mutant flies were created and
vir-1 was found to be fully inducible, which suggests Vago may not directly be involved in
vir-1 induction or may involve an alternative mechanism. In addition, this analysis was limited by the inability to evaluate the Vago protein as it was unstable. The authors concluded additional research would need to be conducted to determine the roles and interaction of Vago protein with Vir-1 [
9]. In
Culex mosquitos infected with WNV,
vir-1 was found upregulated via the JAK-STAT pathway indicating that Vago may play a role in inducing transcription of
vir-1 via the aforementioned proposed mechanism. In addition, vir-1 may also be induced by alternative unidentified virus-activated cytokines in absence of Vago expression in
Culex and
Drosophila [
7]. As the antiviral defense mechanism for Vago is characterized, the role of Vir-1 may be better identified.
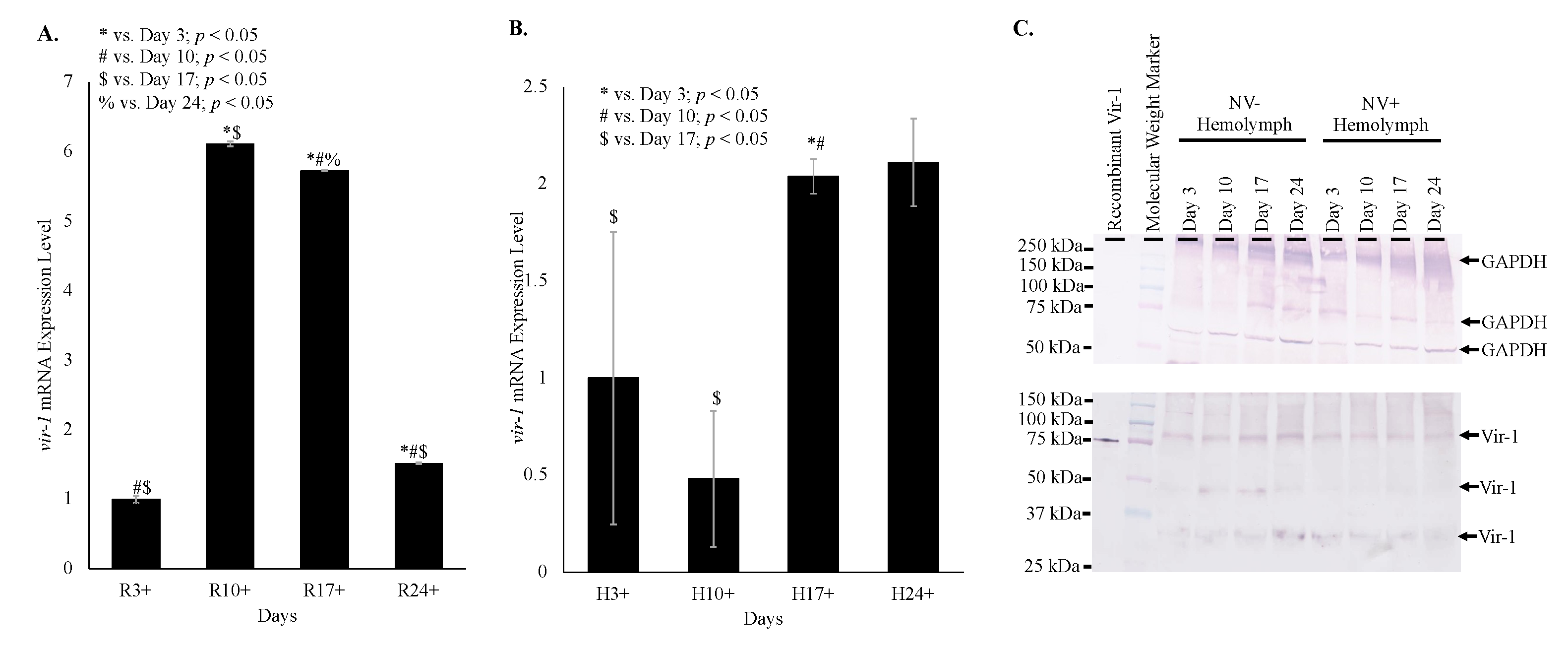
Time course data shows increasing transcription of
vir-1 in Nora virus-infected
D. melanogaster as the Nora virus load increased (
Figure 4). Increased transcription of
vir-1 has been demonstrated within 24 h after
D. melanogaster infection with DCV [
4]. Little is known about Vir-1, but the current study found a transition with age from high
vir-1 transcription in whole fly lysate early in life to increased transcription in hemolymph later in life (
Figure 5). This may suggest a cytokine-like role of Vago in response to Nora virus infection. In this role, Vago may be produced and secreted by infected intestinal cells and act upon the JAK/STAT pathway of hemocytes. In this way, transcription of
vir-1 may increase in the hemolymph despite decreasing
vago transcription levels by day 24. Production of another cytokine-like molecule, Upd3, has also been shown to be required for cytokine-like immune signaling in the hemolymph of septic
D. melanogaster infected with
Escherichia coli or
Micrococcus luteus [
28]. In this study, Vir-1 was found to be present in virus-free
D. melanogaster (
Figure 4 and
Figure 5). This previously unidentified presence of Vir-1 may suggest baseline expression or an additional role of Vir-1. It is unlikely that this Vir-1 presence is due to infection with another virus as the stocks are regularly dechorionated and checked for the main
Drosophila viruses. One study investigating the immune response in cold tolerance identified upregulation of
vir-1 in response to heat shock [
11]. Further studies are needed to determine the function of Vir-1 in virus-free
D. melanogaster, but this role may include a baseline expression level allowing for more rapid response to heat shock or viral infection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




