Characteristics of a Novel Target Antigen against Myeloma Cells for Immunotherapy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Reagents
2.3. Detection of CXorf48 Gene Expression by RT–PCR
2.4. Detection of Cancer/Testis Antigen (CTA) Gene Expression by RT-PCR
2.5. Detection of CXorf48 Protein Expression by Immunocytochemical Staining
2.6. Treatment with Demethylating Agent
2.7. Generation of CXorf48-Specific CTLs from Human PBMNC
2.8. Detection of IFN-γ Secretion from CTLs by Enzyme-Linked Immunospot (ELISpot) Assay
2.9. Detection of Cytotoxicity of CTLs by Cytotoxicity Assay
2.10. Dextramer Staining
2.11. Statistical Analysis
3. Results
3.1. CXorf48 Is Expressed in Myeloma Cells
3.2. CXorf48-Specific CTLs Recognized Myeloma Cells with High Expression of CXorf48
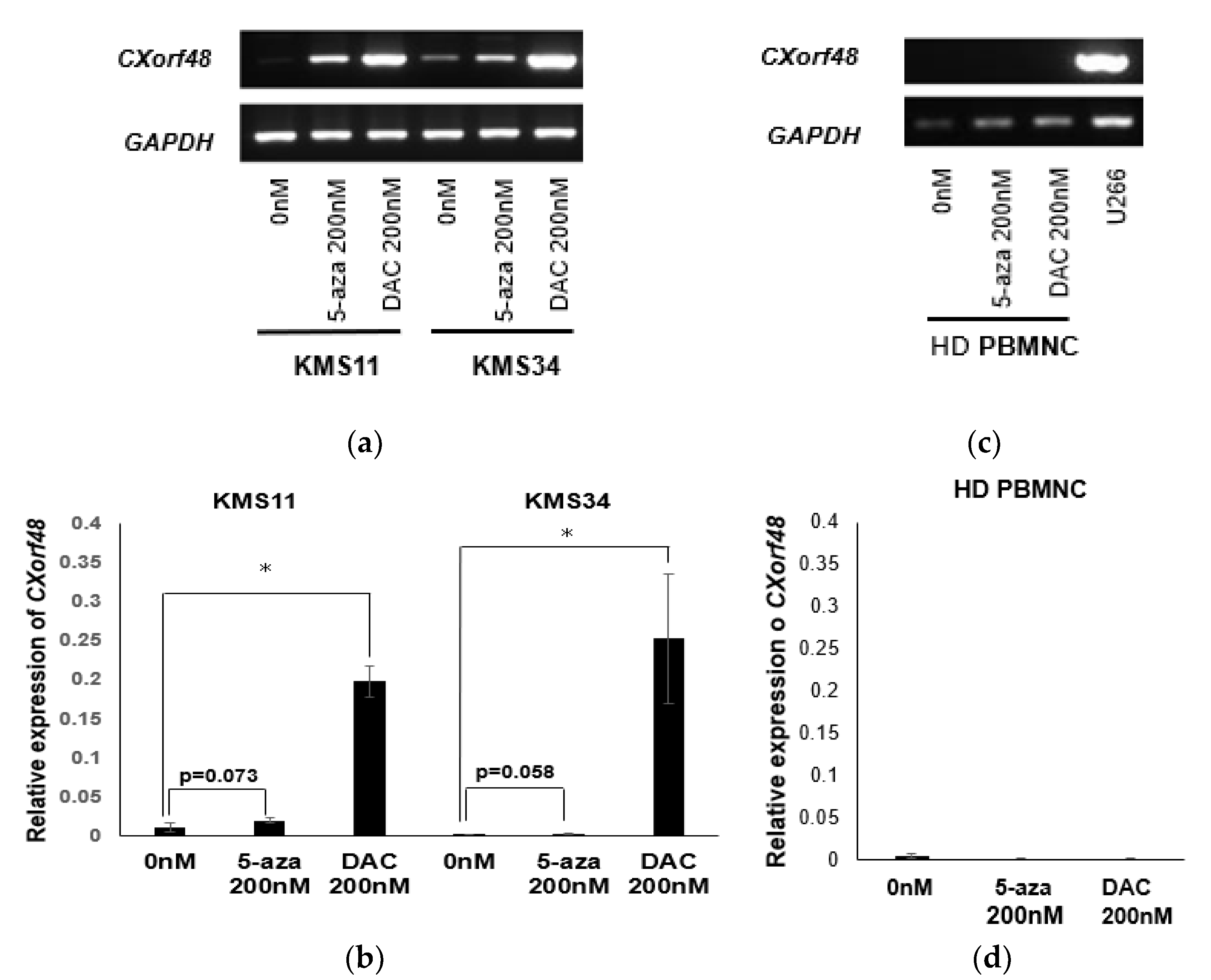
3.3. Up-regulation of CXorf48 Expression by Demethylating Agents
3.4. CXorf48-Specific CTL Recognized DMA-Treated Myeloma Cells with Low CXorf48 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rajkumar, S.V.; Kumar, S. Multiple Myeloma: Diagnosis and Treatment. Mayo Clin. Proc. 2016, 91, 101–119. [Google Scholar] [CrossRef] [PubMed]
- Kazandjian, D. Multiple myeloma epidemiology and survival: A unique malignancy. Semin. Oncol. 2016, 43, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Pinto, V.; Bergantim, R.; Caires, H.R.; Seca, H.; Guimarães, J.E.; Vasconcelos, M.H. Multiple Myeloma: Available Therapies and Causes of Drug Resistance. Cancers 2020, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Abramson, H.N. Monoclonal Antibodies for the Treatment of Multiple Myeloma: An Update. Int. J. Mol. Sci. 2018, 19, 3924. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, B.; Hari, P.N.; Usmani, S.Z.; Hamadani, M. Chimeric antigen receptor T cell therapy in multiple myeloma: Promise and challenges. Bone Marrow Transplant. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mailankody, S.; Korde, N.; Lesokhin, A.M.; Lendvai, N.; Hassoun, H.; Stetler-Stevenson, M.; Landgren, O. Minimal residual disease in multiple myeloma: Bringing the bench to the bedside. Nat. Rev. Clin. Oncol. 2015, 12, 286–295. [Google Scholar] [CrossRef]
- Munshi, N.C.; Avet-Loiseau, H.; Rawstron, A.C.; Owen, R.G.; Child, J.A.; Thakurta, A.; Sherrington, P.; Samur, M.K.; Georgieva, A.; Anderson, K.C.; et al. Association of Minimal Residual Disease With Superior Survival Outcomes in Patients With Multiple Myeloma: A Meta-analysis. JAMA Oncol. 2017, 3, 28–35. [Google Scholar] [CrossRef]
- Oka, Y.; Tsuboi, A.; Nakata, J.; Nishida, S.; Hosen, N.; Kumanogoh, A.; Oji, Y.; Sugiyama, H. Wilms’ Tumor Gene 1 (WT1) Peptide Vaccine Therapy for Hematological Malignancies: From CTL Epitope Identification to Recent Progress in Clinical Studies Including a Cure-Oriented Strategy. Oncol. Res. Treat. 2017, 40, 682–690. [Google Scholar] [CrossRef]
- Ghobrial, I.; Cruz, C.H.; Garfall, A.; Shah, N.; Munshi, N.; Kaufman, J.; Boise, L.H.; Morgan, G.; Adalsteinsson, V.A.; Manier, S.; et al. Immunotherapy in Multiple Myeloma: Accelerating on the Path to the Patient. Clin. Lymphoma Myeloma Leuk. 2019, 19, 332–344. [Google Scholar] [CrossRef]
- Lu, C.; Meng, S.; Jin, Y.; Zhang, W.; Li, Z.; Wang, F.; Wang-Johanning, F.; Wei, Y.; Liu, H.; Tu, H.; et al. A novel multi-epitope vaccine from MMSA-1 and DKK1 for multiple myeloma immunotherapy. Br. J. Haematol. 2017, 178, 413–426. [Google Scholar] [CrossRef]
- Matsushita, M.; Ozawa, K.; Suzuki, T.; Nakamura, M.; Nakano, N.; Kanchi, S.; Ichikawa, D.; Matsuki, E.; Sakurai, M.; Karigane, D.; et al. CXorf48 is a potential therapeutic target for achieving treatment-free remission in CML patients. Blood Cancer J. 2017, 7, e601. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Du, W.; Yamada, T.; Ichikawa, D.; Matsunami, S.; Matsushita, M. A myeloma cell line established from a patient refractory to thalidomide therapy revealed high-risk cytogenetic abnormalities and produced vascular endothelial growth factor. Blood Cancer J. 2013, 3, e115. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matsushita, M.; Otsuka, Y.; Tsutsumida, N.; Tanaka, C.; Uchiumi, A.; Ozawa, K.; Suzuki, T.; Ichikawa, D.; Aburatani, H.; Okamoto, S.; et al. Identification of Novel HLA-A*24:02-Restricted Epitope Derived from a Homeobox Protein Expressed in Hematological Malignancies. PLoS ONE 2016, 11, e0146371. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Bartlett, D.L.; Gorry, M.C.; O’Malley, M.E.; Guo, Z.S. Three epigenetic drugs up-regulate homeobox gene Rhox5 in cancer cells through overlapping and distinct molecular mechanisms. Mol. Pharmacol. 2009, 76, 1072–1081. [Google Scholar] [CrossRef]
- Cruz, C.R.; Gerdemann, U.; Leen, A.M.; Shafer, J.A.; Ku, S.; Tzou, B.; Horton, T.M.; Sheehan, A.; Copeland, A.; Younes, A.; et al. Improving T-cell therapy for relapsed EBV-negative Hodgkin lymphoma by targeting upregulated MAGE-A4. Clin. Cancer Res. 2011, 17, 7058–7066. [Google Scholar] [CrossRef]
- Li, L.; Goedegebuure, S.P.; Gillanders, W.E. Preclinical and clinical development of neoantigen vaccines. Ann. Oncol. 2017, 28, xii11–xii17. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef]
- Paul, B.; Kang, S.; Zheng, Z.; Kang, Y. The challenges of checkpoint inhibition in the treatment of multiple myeloma. Cell Immunol. 2018, 334, 87–98. [Google Scholar] [CrossRef]
- Gjerstorff, M.F.; Andersen, M.H.; Ditzel, H.J. Oncogenic cancer/testis antigens: Prime candidates for immunotherapy. Oncotarget 2015, 6, 15772–15787. [Google Scholar] [CrossRef]
- Tsuboi, A.; Oka, Y.; Nakajima, H.; Fukuda, Y.; Elisseeva, O.A.; Yoshihara, S.; Hosen, N.; Ogata, A.; Kito, K.; Fujiki, F.; et al. Wilms tumor gene WT1 peptide-based immunotherapy induced a minimal response in a patient with advanced therapy-resistant multiple myeloma. Int. J. Hematol. 2007, 86, 414–417. [Google Scholar] [CrossRef]
- Dawood, R.M.; Moustafa, R.I.; Abdelhafez, T.H.; El-Shenawy, R.; El-Abd, Y.; Bader El Din, N.G.; Dubuisson, J.; El Awady, M.K. A multiepitope peptide vaccine against HCV stimulates neutralizing humoral and persistent cellular responses in mice. BMC Infect Dis. 2019, 19, 932. [Google Scholar] [CrossRef] [PubMed]
- Zamunér, F.T.; Karia, B.T.; de Oliveira, C.Z.; Santos, C.R.; Carvalho, A.L.; Vettore, A.L. A Comprehensive Expression Analysis of Cancer Testis Antigens in Head and Neck Squamous Cell Carcinoma Revels MAGEA3/6 as a Marker for Recurrence. Mol. Cancer Ther. 2015, 14, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Santra, M.; Zhan, F.; Tian, E.; Barlogie, B.; Shaughnessy, J. A subset of multiple myeloma harboring the t(4;14)(p16;q32) translocation lacks FGFR3 expression but maintains an IGH/MMSET fusion transcript. Blood 2003, 101, 2374–2376. [Google Scholar] [CrossRef]
- Chesi, M.; Nardini, E.; Brents, L.A.; Schröck, E.; Ried, T.; Kuehl, W.M.; Bergsagel, P.L. Frequent translocation t(4;14)(p16.3;q32.3) in multiple myeloma is associated with increased expression and activating mutations of fibroblast growth factor receptor 3. Nat. Genet. 1997, 16, 260–264. [Google Scholar] [CrossRef]
- Matsushita, M.; Yamazaki, R.; Ikeda, H.; Kawakami, Y. Preferentially expressed antigen of melanoma (PRAME) in the development of diagnostic and therapeutic methods for hematological malignancies. Leuk. Lymphoma 2003, 44, 439–444. [Google Scholar] [CrossRef]
- Nguyen, L.P.; Galtier, N.; Nabholz, B. Gene expression, chromosome heterogeneity and the fast-X effect in mammals. Boil. Lett. 2015, 11, 20150010. [Google Scholar] [CrossRef] [PubMed]
- Pujol, J.L.; De Pas, T.; Rittmeyer, A.; Vallières, E.; Kubisa, B.; Levchenko, E.; Wiesemann, S.; Masters, G.A.; Shen, R.; Tjulandin, S.A.; et al. Safety and Immunogenicity of the PRAME Cancer Immunotherapeutic in Patients with Resected Non-Small Cell Lung Cancer: A Phase I Dose Escalation Study. J. Thorac. Oncol. 2016, 11, 2208–2217. [Google Scholar] [CrossRef]
- Azizi, A.; Ediriwickrema, A.; Dutta, R.; Patel, S.A.; Shomali, W.; Medeiros, B.; Iberri, D.; Gotlib, J.; Mannis, G.; Greenberg, P.; et al. Venetoclax and hypomethylating agent therapy in high risk myelodysplastic syndromes: A retrospective evaluation of a real-world experience. Leuk. Lymphoma 2020, 16, 1–8. [Google Scholar] [CrossRef]
- Li, N.; Liu, L.; Xiang, P.; Liang, L.; Wang, J.; Wang, Y.; Luo, S.; Song, Y.; Fang, B. Addition of low-dose decitabine to bortezomib and dexamethasone as second-line therapy in multiple myeloma. Br. J. Haematol. 2020, 189, e258–e262. [Google Scholar] [CrossRef]
- Zhou, J.; Shen, Q.; Lin, H.; Hu, L.; Li, G.; Zhang, X. Decitabine shows potent anti-myeloma activity by depleting monocytic myeloid-derived suppressor cells in the myeloma microenvironment. J. Cancer. Res. Clin. Oncol. 2019, 145, 329–336. [Google Scholar] [CrossRef]
- Grieve, S.; Wajnberg, G.; Lees, M.; Chacko, S.; Weir, J.; Crapoulet, N.; Reiman, T. TAZ functions as a tumor suppressor in multiple myeloma by downregulating MYC. Blood Adv. 2019, 3, 3613–3625. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsushita, M.; Saito, S.; Yokoe, S.; Ichikawa, D.; Hattori, Y. Characteristics of a Novel Target Antigen against Myeloma Cells for Immunotherapy. Vaccines 2020, 8, 579. https://doi.org/10.3390/vaccines8040579
Matsushita M, Saito S, Yokoe S, Ichikawa D, Hattori Y. Characteristics of a Novel Target Antigen against Myeloma Cells for Immunotherapy. Vaccines. 2020; 8(4):579. https://doi.org/10.3390/vaccines8040579
Chicago/Turabian StyleMatsushita, Maiko, Saku Saito, Shinya Yokoe, Daiju Ichikawa, and Yutaka Hattori. 2020. "Characteristics of a Novel Target Antigen against Myeloma Cells for Immunotherapy" Vaccines 8, no. 4: 579. https://doi.org/10.3390/vaccines8040579
APA StyleMatsushita, M., Saito, S., Yokoe, S., Ichikawa, D., & Hattori, Y. (2020). Characteristics of a Novel Target Antigen against Myeloma Cells for Immunotherapy. Vaccines, 8(4), 579. https://doi.org/10.3390/vaccines8040579