
Immune Protective Efficacy of China’s Traditional Inactivated and Attenuated Vaccines against the Prevalent Strains of Pasteurella multocida in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Animals
2.3. Live Vaccine and Adjuvant
2.4. Inactivated Alh–C44-1 Vaccine Formulation
2.5. Detection of Virulence
2.6. Immunization and Challenge Schedule
2.7. Measurement of Serum Antibody Titers
2.8. Statistical Analysis
3. Results
3.1. Virulence of the P. multocida Strains in Mice
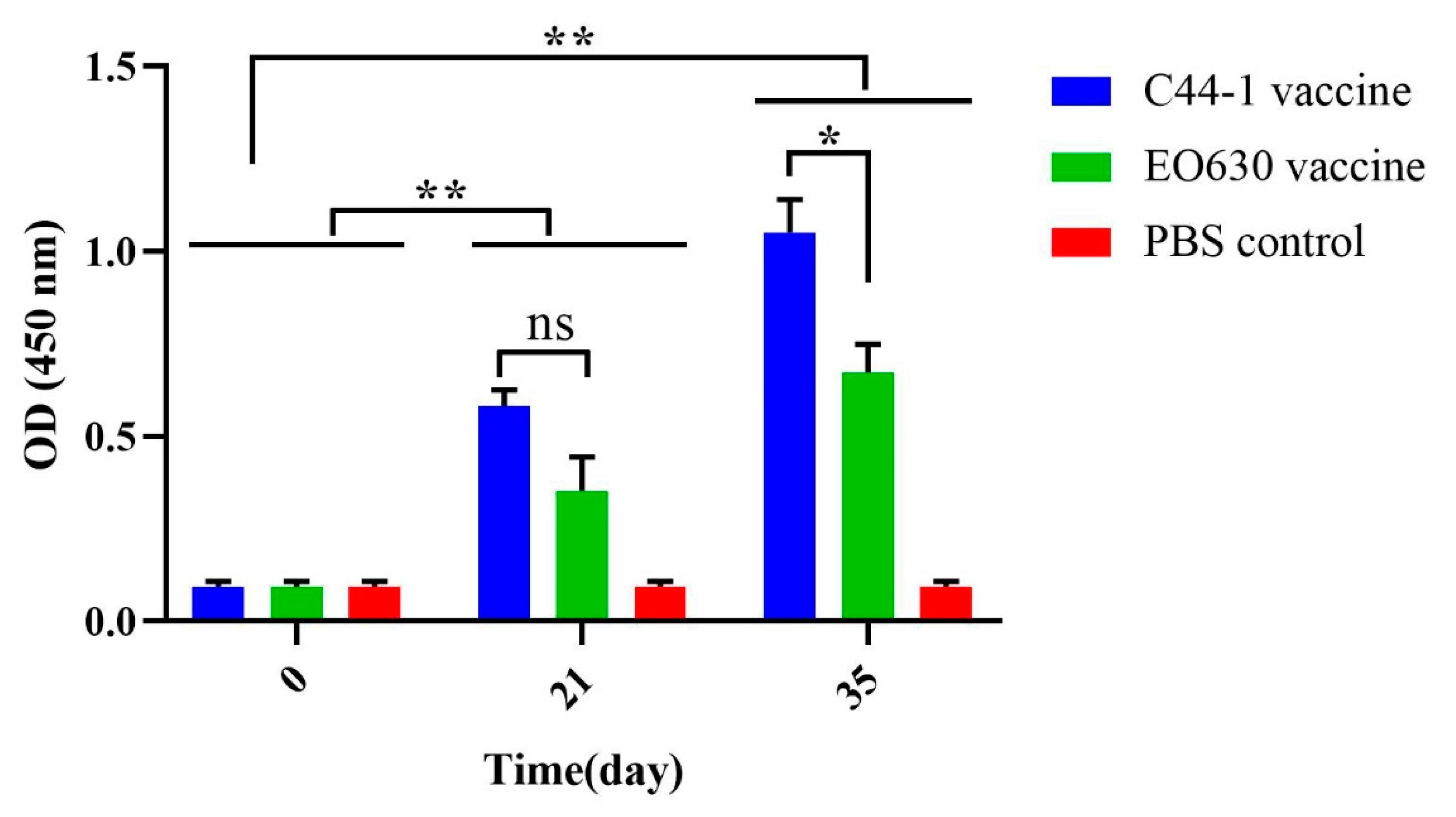
3.2. Serum Antibody Titers
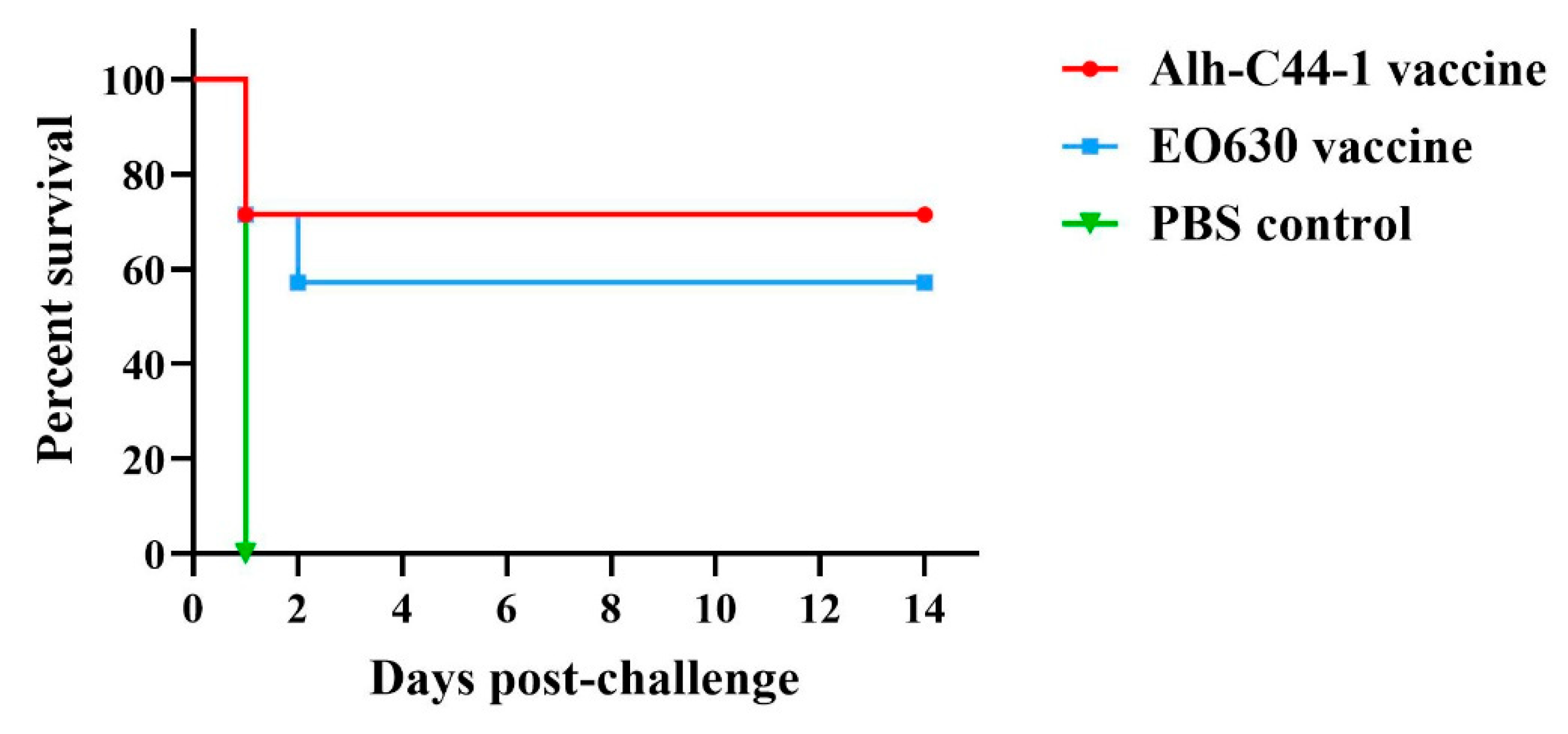
3.3. Protective Effect of Traditional Vaccines against P. multocida Serogroup B Strain
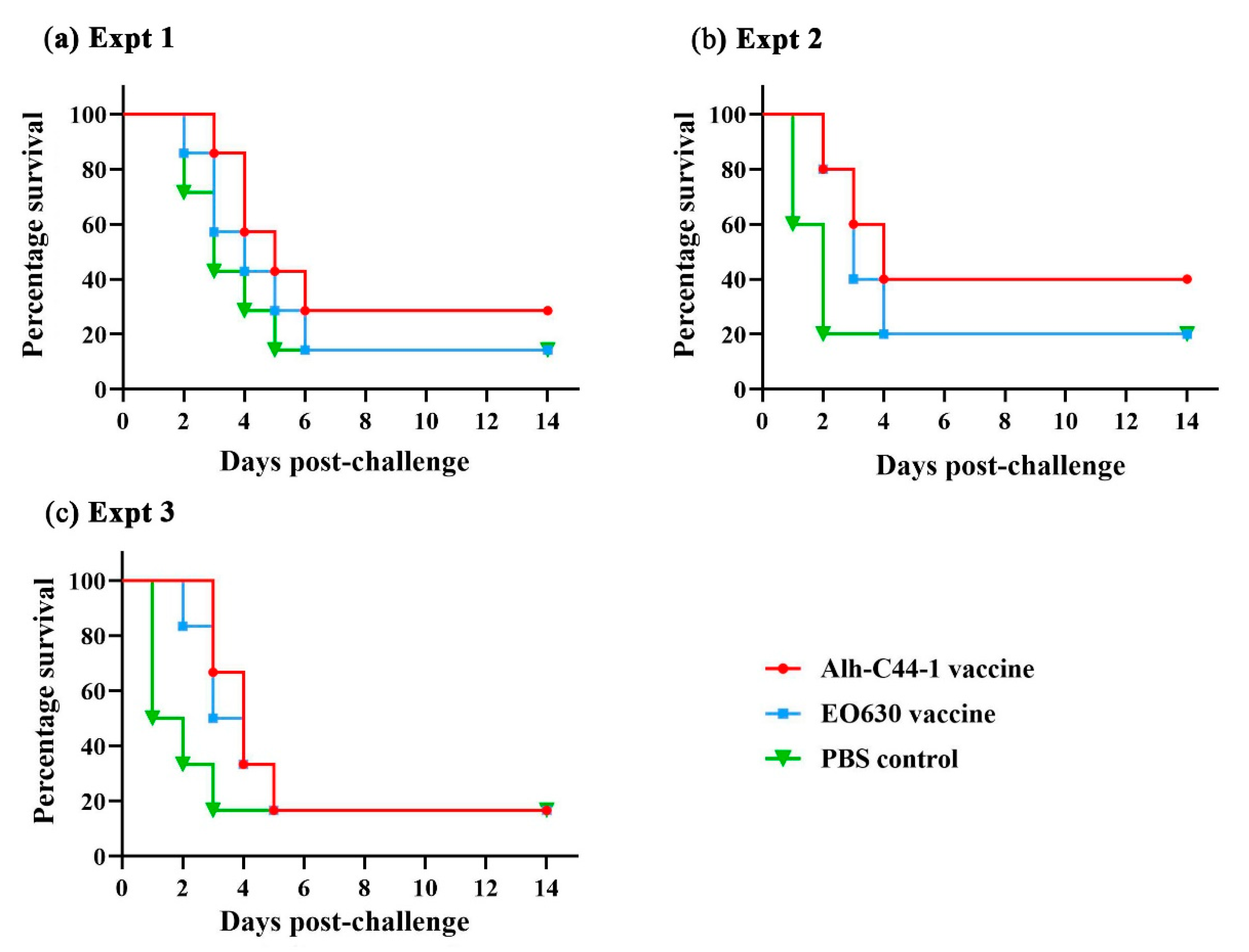
3.4. Protective Effect of Traditional Vaccines against P. multocida Serogroup A Strains
3.5. Protective Effect of Traditional Vaccines against P. multocida Serogroup D Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kubatzky, K.F. Pasteurella multocida and immune cells. Curr. Top. Microbiol. Immunol. 2012, 361, 53–72. [Google Scholar]
- de Cecco, B.S.; Carossino, M.; Del Piero, F.; Wakamatsu, N.; Mitchell, M.S.; Fowlkes, N.W.; Langohr, I.M. Meningoencephalomyelitis in domestic cats: 3 cases of Pasteurella multocida infection and literature review. J. Vet. Diagn. Investig. 2021. [Google Scholar] [CrossRef]
- Zhao, G.; Li, P.; Mu, H.; Li, N.; Peng, Y. L-ascorbic acid shapes bovine Pasteurella multocida serogroup A infection. Front. Vet. Sci. 2021, 8, 687922. [Google Scholar] [CrossRef]
- Abdulrahman, R.F.; Davies, R.L. Diversity and characterization of temperate bacteriophages induced in Pasteurella multocida from different host species. BMC Microbiol. 2021, 21, 97. [Google Scholar] [CrossRef]
- Glisson, J.R.; Hofacre, C.L.; Christensen, J.P. Fowl cholera. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; Wiley-Blackwell: Ames, IA, USA, 2013; pp. 807–823. ISBN 978-0-4709-5899-5. [Google Scholar]
- Boyce, J.D.; Harper, M.; Wilkie, I.W.; Adler, B. Pasteurella. In Pathogenesis of Bacterial Infections in Animals, 4th ed.; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 325–346. ISBN 9780813812373. [Google Scholar]
- Kim, J.; Kim, J.W.; Oh, S.-I.; So, B.; Kim, W.-I.; Kim, H.-Y. Characterisation of Pasteurella multocida isolates from pigs with pneumonia in Korea. BMC Vet. Res. 2019, 15, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisco, L.; Cahuayme-Zuniga, L. Acute epiglottitis due to Pasteurella multocida after contact with a feral cat. Baylor Univ. Med. Cent. Proc. 2019, 32, 574–576. [Google Scholar] [CrossRef] [PubMed]
- Aida, Y.; Kiwamoto, T.; Fujita, K.; Ishikawa, H.; Kitazawa, H.; Watanabe, H.; Hizawa, N. Pasteurella multocida pneumonia with hemoptysis: A case report. Respir. Med. Case Rep. 2019, 26, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Ayas, M.F.; Youssef, D.; Johnson, L. A rare case of bacteremic community-acquired pneumonia due to Pasteurella multocida presenting with hemoptysis. Cureus 2021, 13, e14232. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Chung, J.Y.; Adler, B. Pasteurella multocida capsule: Composition, function and genetics. J. Biotechnol. 2000, 83, 153–160. [Google Scholar] [CrossRef]
- Carter, G.R. The type specific capsular antigen of Pasteurella multocida. Can. J. Med. Sci. 1952, 30, 48–53. [Google Scholar] [CrossRef]
- Rimler, R.B.; Rhoades, K.R. Serogroup F, a new capsule serogroup of Pasteurella multocida. J. Clin. Microbiol. 1987, 25, 615–618. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhao, Z.; Xi, X.; Xue, Q.; Long, T.; Xue, Y. Occurrence of Pasteurella multocida among pigs with respiratory disease in China between 2011 and 2015. Ir. Vet. J. 2017, 70, 2. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Liang, W.; Ai, W.C.; Wang, F.; Hua, L.; Tang, X.B.; Chen, H.C.; Wu, B. Genotypical characteristics of swine Pasteurella multocida in China. Acta Vet. Zootech. Sin. 2019, 50, 1064–1072. (In Chinese) [Google Scholar]
- Zhang, J.; Wang, M.; Zhou, N.; Shen, Y.; Li, Y. Evaluation of carbopol as an adjuvant on the effectiveness of progressive atrophic rhinitis vaccine. Vaccine 2018, 36, 4477–4484. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, E.N.T.; Walker, J.; Peters, A.; Pastoret, P.-P.; Jungersen, G. Current status of veterinary vaccines. Clin. Microbiol. Rev. 2007, 20, 489–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Test report of attenuated EO630 strain of swine plague. Chengdu Vet. Bio-Pharm. Fact. 1976, 2, 18–27. (In Chinese)
- Wu, F.G.; Qian, X.Y. Serotype of Pasteurella multocida heat-resistant antigen in China (First report). Chin. J. Vet. Med. 1987, 12, 2–4. (In Chinese) [Google Scholar]
- Wu, F.G. Serological identification and interactive immunoassay of Pasteurella multocida in China. Chin. J. Vet. Med. 1997, 8, 14–15. (In Chinese) [Google Scholar]
- Zhao, Z.Q.; Liu, Q.Y.; Xi, X.J.; Wang, L.; Deng, W.; Zhang, C.J. A comparative study on biological characterization of Pasteurella multocida serogroups A and D isolates from swine in China. Chin. J. Prev. Vet. Med. 2006, 38, 366–370. (In Chinese) [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Harper, M.; Boyce, J.D. The myriad properties of Pasteurella multocida lipopolysaccharide. Toxins 2017, 9, 254. [Google Scholar] [CrossRef] [Green Version]
- Puspitasari, Y.; Salleh, A.; Zamri-Saad, M. Ultrastructural changes in endothelial cells of buffaloes following in-vitro exposure to Pasteurella multocida B:2. BMC Vet. Res. 2020, 16, 186. [Google Scholar] [CrossRef]
- Ewers, C.; Lubkebecker, A.; Bethe, A.; Kiesling, S.; Filter, M.; Wieler, L. Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet. Microbiol. 2006, 114, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Soriano-Vargas, E.; Vega-Sánchez, V.; Zamora-Espinosa, J.L.; Acosta-Dibarrat, J.; Aguilar-Romero, F.; Negrete-Abascal, E. Identification of Pasteurella multocida capsular types isolated from rabbits and other domestic animals in Mexico with respiratory diseases. Trop. Anim. Health Prod. 2012, 44, 935–937. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Meng, X.-Z.; Geng, H.-L.; Chang, C.; Chen, X.; Wen, X.; Ni, H. Generation of porcine Pasteurella multocida ghost vaccine and examination of its immunogenicity against virulent challenge in mice. Microb. Pathog. 2019, 132, 208–214. [Google Scholar] [CrossRef]
- Qureshi, S.; Saxena, H.M. Estimation of titers of antibody against Pasteurella multocida in cattle vaccinated with haemorrhagic septicemia alum precipitated vaccine. Vet. World 2014, 7, 224–228. [Google Scholar] [CrossRef]
- Okerman, L.; Devriese, L.A. Failure of oil adjuvants to enhance immunity induced in mice by an inactivated rabbit Pasteurella multocida vaccine. Vaccine 1987, 5, 315–318. [Google Scholar] [CrossRef]
- Arif, J.; Rahman, S.-U.; Arshad, M.; Akhtar, P. Immunopotentiation of outer membrane protein through anti-idiotype Pasteurella multocida vaccine in rabbits. Biologicals 2013, 41, 339–344. [Google Scholar] [CrossRef]
- Hsuan, S.-L.; Liao, C.-M.; Huang, C.; Winton, J.R.; Chen, Z.-W.; Lee, W.-C.; Liao, J.-W.; Chen, T.-H.; Chiou, C.-J.; Yeh, K.-S.; et al. Efficacy of a novel Pasteurella multocida vaccine against progressive atrophic rhinitis of swine. Vaccine 2009, 27, 2923–2929. [Google Scholar] [CrossRef]
- Register, K.B.; Sacco, R.E.; Brockmeier, S.L. Immune response in mice and swine to DNA vaccines derived from the Pasteurella multocida toxin gene. Vaccine 2007, 25, 6118–6128. [Google Scholar] [CrossRef]
- Silveira, M.; Vargas, S.; Mendonça, M.; Cunha, C.E.; Hartwig, D.; Seixas, A.; Bampi, S.; Rodrigues, A.; Conceição, F.; Moreira, Â. Xanthan gum enhances humoral immune response elicited by a DNA vaccine against leptospirosis in mice. BMC Proc. 2014, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Devi, L.B.; Bora, D.P.; Das, S.K.; Sharma, R.K.; Mukherjee, S.; Hazarika, R.A. Virulence gene profiling of porcine Pasteurella multocida isolates of Assam. Vet. World 2018, 11, 348–354. [Google Scholar] [CrossRef]
- Katsuda, K.; Hoshinoo, K.; Ueno, Y.; Kohmoto, M.; Mikami, O. Virulence genes and antimicrobial susceptibility in Pasteurella multocida isolates from calves. Vet. Microbiol. 2013, 167, 737–741. [Google Scholar] [CrossRef]
- Guan, L.; Xue, Y.; Ding, W.; Zhao, Z. Biosynthesis and regulation mechanisms of the Pasteurella multocida capsule. Res. Vet. Sci. 2019, 127, 82–90. [Google Scholar] [CrossRef]
- Cappellano, G.; Abreu, H.; Casale, C.; Dianzani, U.; Chiocchetti, A. Nano-microparticle platforms in developing next-generation vaccines. Vaccines 2021, 9, 606. [Google Scholar] [CrossRef]
- Homayoon, M.; Tahamtan, Y.; Kargar, M.; Hosseini, S.M.H.; Akhavan Sepahy, A. Pasteurella multocida inactivated with ferric chloride and adjuvanted with bacterial DNA is a potent and efficacious vaccine in Balb/c mice. J. Med. Microbiol. 2018, 67, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.H.; Jacobs, A.A.; Shah, N.H.; de Graaf, F.K. Safety and efficacy of an oil-adjuvant vaccine against haemorrhagic septicaemia in buffalo calves: Cross-protection between the serotypes B:2,5 and E:2,5. Vet. Rec. 2001, 149, 583–587. [Google Scholar] [CrossRef]
- Abusalab, S.; Elhaj, M.O.; Mohamed, L.I.; Mekki, N.T.; Ahmed, A.M. Test of cross reaction between the two local sudanese strains of P. mulocida (B and E). Pakistan J. Biol. Sci. 2013, 16, 1388–1392. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Chung, J.Y.; Adler, B. Genetic organisation of the capsule biosynthetic locus of Pasteurella multocida M1404 (B:2). Vet. Microbiol. 2000, 72, 121–134. [Google Scholar] [CrossRef]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic organization of Pasteurella multocida cap loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Wang, M. The Research on Cattle Bivalent Vaccine to Both Type A and B Pasteurella multocida; Huazhong Agricultural University: Wuhan, China, 2013. (In Chinese) [Google Scholar]
- Tao, Q.X.C.; Wu, Q.; Gu, X.X.; Ma, X.; Li, J.; Zhou, X.; Zhang, X.X.; Huang, X.; Wu, T.Z.; Han, M.L.; et al. Preparation of bovine type A Pm12 inactivated vaccine and attack test in mice. Heilongjiang Anim. Sci. Vet. Med. 2021, 5, 127–130. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunization Groups | Immune Dose (µL) | Challenge Strain | Challenge Dose (CFU) | No. of Mice | No. of Mice Survived | Protective Efficacy |
|---|---|---|---|---|---|---|
| Alh-C44-1 inactivated vaccine | 200 | C44-1 | 12 LD50 | 7 | 5 | 71% |
| EO630 live vaccine | 200 | C44-1 | 12 LD50 | 7 | 4 | 57% |
| Control | 200 | C44-1 | 12 LD50 | 7 | 0 | 0 |
| Experiment | Immunization Group | Immune Dose (µL) | Challenge Strain | Challenge Dose (CFU) | No. of Mice | No. of Mice Survived | Protective Efficacy |
|---|---|---|---|---|---|---|---|
| Experiment 1 | Alh-C44-1 inactivated vaccines | 200 | PM-5 | 18 LD50 | 7 | 2 | 29% |
| EO630 live vaccine | 200 | PM-5 | 18 LD50 | 7 | 1 | 14% | |
| Control | 200 | PM-5 | 18 LD50 | 7 | 1 | 14% | |
| Experiment 2 | Alh-C44-1 inactivated vaccines | 200 | PM-5 | 29 LD50 | 5 | 2 | 40% |
| EO630 live vaccine | 200 | PM-5 | 29 LD50 | 5 | 1 | 20% | |
| Control | 200 | PM-5 | 29 LD50 | 5 | 1 | 20% | |
| Experiment 3 | Alh-C44-1 inactivated vaccines | 200 | PM-10 | 10 LD50 | 6 | 1 | 17% |
| EO630 live vaccine | 200 | PM-10 | 10 LD50 | 6 | 1 | 17% | |
| Control | 200 | PM-10 | 10 LD50 | 6 | 1 | 17% |
| Experiment | Immunization Group | Immune Dose (µL) | Challenge Strain | Challenge Dose (CFU) | No. of Mice | No. of Mice Survived | Protective Efficacy |
|---|---|---|---|---|---|---|---|
| Experiment 4 | Alh-C44-1 inactivated vaccines | 200 | PM-15 | 8 LD50 | 7 | 2 | 29% |
| EO630 live vaccine | 200 | PM-15 | 8 LD50 | 7 | 2 | 29% | |
| Control | 200 | PM-15 | 8 LD50 | 7 | 1 | 14% | |
| Experiment 5 | Alh-C44-1 inactivated vaccine | 200 | PM-15 | 10 LD50 | 5 | 1 | 20% |
| EO630 live vaccine | 200 | PM-15 | 10 LD50 | 5 | 1 | 20% | |
| Control | 200 | PM-15 | 10 LD50 | 5 | 0 | 0 | |
| Experiment 6 | Alh-C44-1 inactivated vaccine | 200 | PM-G1 | 25 LD50 | 5 | 1 | 20% |
| EO630 live vaccine | 200 | PM-G1 | 25 LD50 | 5 | 1 | 20% | |
| Control | 200 | PM-G1 | 25 LD50 | 5 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, L.-J.; Song, J.-J.; Xue, Y.; Ai, X.; Liu, Z.-J.; Si, L.-F.; Li, M.-Y.; Zhao, Z.-Q. Immune Protective Efficacy of China’s Traditional Inactivated and Attenuated Vaccines against the Prevalent Strains of Pasteurella multocida in Mice. Vaccines 2021, 9, 1155. https://doi.org/10.3390/vaccines9101155
Guan L-J, Song J-J, Xue Y, Ai X, Liu Z-J, Si L-F, Li M-Y, Zhao Z-Q. Immune Protective Efficacy of China’s Traditional Inactivated and Attenuated Vaccines against the Prevalent Strains of Pasteurella multocida in Mice. Vaccines. 2021; 9(10):1155. https://doi.org/10.3390/vaccines9101155
Chicago/Turabian StyleGuan, Li-Jun, Ji-Jian Song, Yun Xue, Xia Ai, Zhi-Jun Liu, Li-Fang Si, Meng-Yun Li, and Zhan-Qin Zhao. 2021. "Immune Protective Efficacy of China’s Traditional Inactivated and Attenuated Vaccines against the Prevalent Strains of Pasteurella multocida in Mice" Vaccines 9, no. 10: 1155. https://doi.org/10.3390/vaccines9101155
APA StyleGuan, L.-J., Song, J.-J., Xue, Y., Ai, X., Liu, Z.-J., Si, L.-F., Li, M.-Y., & Zhao, Z.-Q. (2021). Immune Protective Efficacy of China’s Traditional Inactivated and Attenuated Vaccines against the Prevalent Strains of Pasteurella multocida in Mice. Vaccines, 9(10), 1155. https://doi.org/10.3390/vaccines9101155