1. Introduction
The aging of the population, also named the “gray” tsunami, is a growing social and public health issue. The impact of the phenomenon is forecasted by worrying data in Italy, one of the most elderly nations in the world [
1]. In particular, the Northeast Veneto region area of Treviso is characterized by the highest longevity in Italy, especially among women [
2]. The reasons are scantily known.
Aging is a peculiar and manifold process characterized by a gradual deterioration of the body’s capability to react to internal and/or external stress factors, resulting in an increased risk of illness and death [
3]. This decline is the key risk factor of many human diseases, including cancer, diabetes, cardiovascular and neurodegenerative disorders. Not all people become eldery in the same manner [
4]. Consequently, chronological age is not a consistent marker of the physiological decay of an individual [
5], while biological age is expected to reflect the continuing changes within a person [
6].
Accumulating evidence has revealed that leukocyte telomere length (LTL) reduction can be regarded as the early pillar of biological aging and may be the origin of cellular dysfunction [
3]. Telomeres are short sequences of DNA repeated (TTAGGG) in association with various proteins forming a complex that is essential in preserving the genome stability of the cells [
7]. In normal somatic cells, telomeric repeats are reduced by 30–200 bp after each mitotic division. However, telomeres, such as triple G-containing sequences, represent a susceptible target for genotoxic compounds, oxidative stress, and inflammation damages that directly accelerate telomere shortening. In our previous works, we showed that LTL erosion is related to risk factors for age-related diseases, including obesity [
8], oxidative stress [
9] and inflammatory responses [
10], exposure to lifestyle (smoke, alcohol) [
11], occupational pollutants [
12] and cancer [
13]. More recently, we found that in 585 residents in Northeast Italy, the everyday life exposure to polluted air, in particular indoors, loses LTL, particularly in males [
14]. Therefore, the rate at which telomeres shorten provides a sturdy biomarker of aging and general health.
However, one of the most discussed questions in gerontology research is whether LTL is prognostic for a long and healthy life. The problem in interpreting the results of several studies arises from the heterogeneity of the factors that affect LTL, including genetics and environment, and in particular the availability of follow-up data both of LTL and survival of enrolled subjects.
The Treviso Longeva (TRELONG) study is a longitudinal cohort study initiated in 2003, which encompassed 668 seniors, women and men, over the age of 70 years, living in Treviso, the town with the highest longevity in North-East Italy [
2]. The Trelong Study has investigated different topics such as frailty, cognitive impairment, Mediterranean diet and genetic aspects [
15,
16,
17,
18,
19,
20].
In view of the above, we investigated LTL within the prospective TRELONG cohort with two main aims: to ascertain whether this biomarker is predictive of longevity and to identify the main determinants in LTL among the personal characteristics (age, sex, marital status, education, occupation, etc.), physiological and lifestyle features, physical performance and frailty measures, chronic diseases, biochemical measurements and apolipoprotein E (APOE) genotyping.
4. Discussion
4.1. LTL and Survival
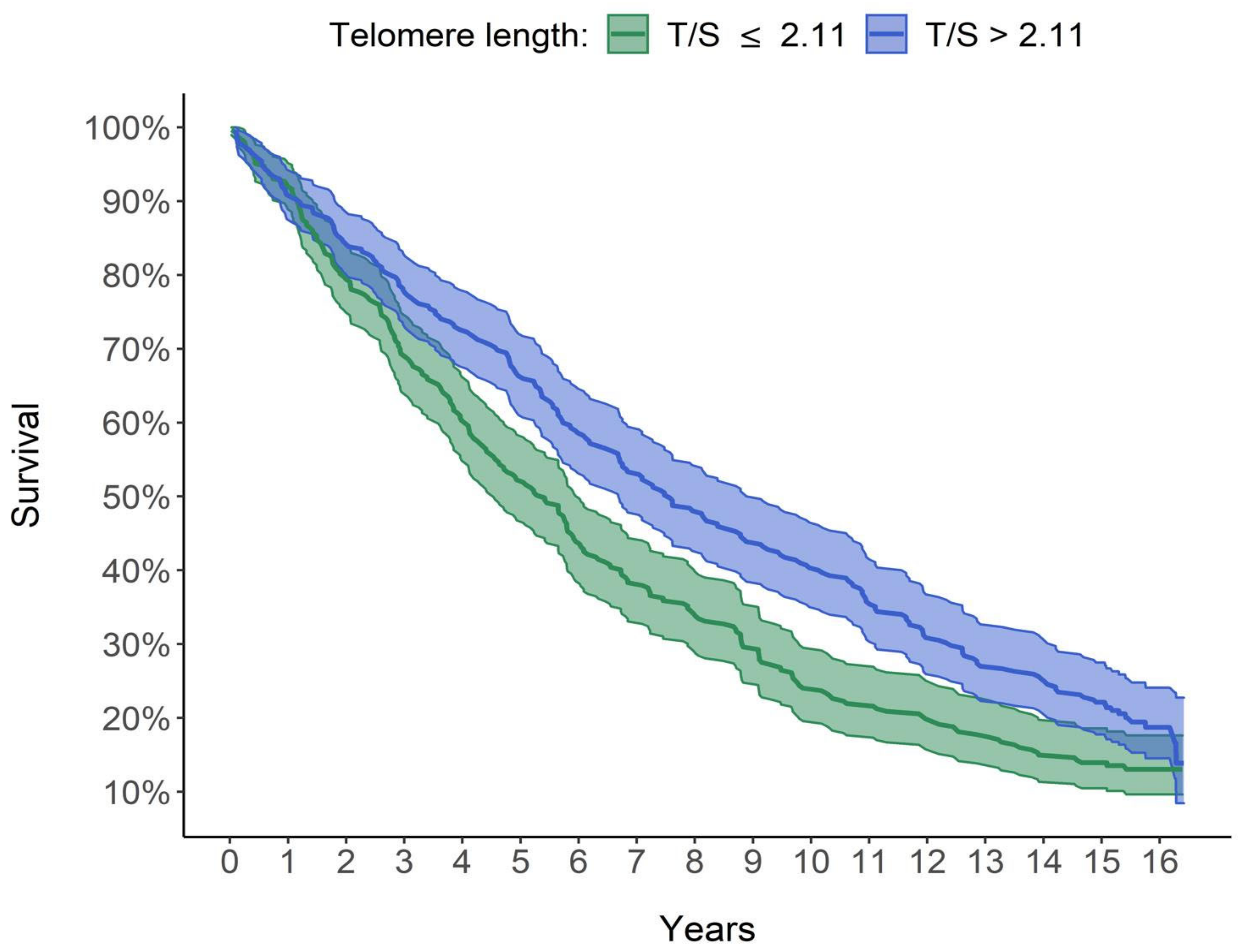
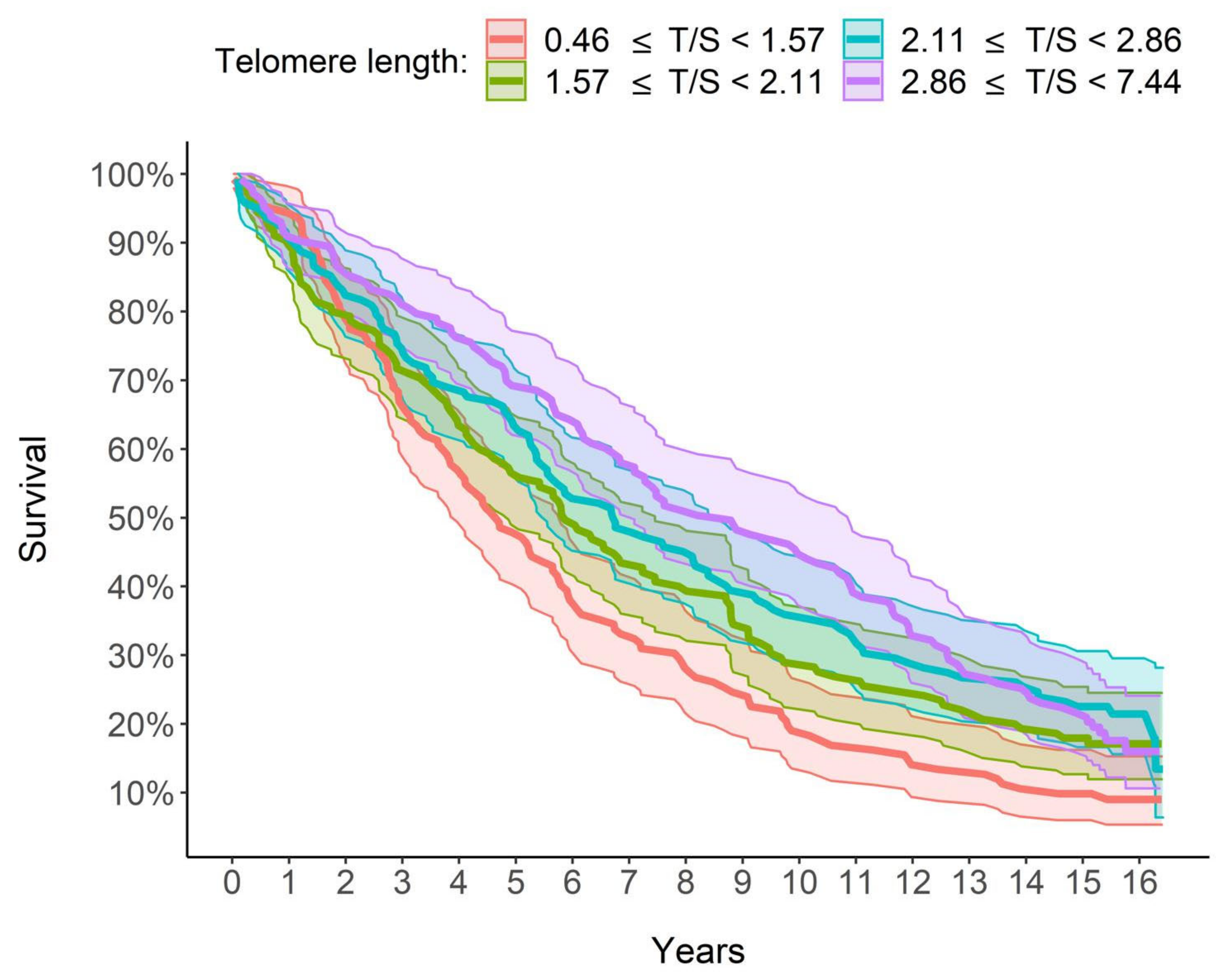
A key finding from our study is the steadily increasing relationship between LTL and survival: the longer the LTL, the greater the survival and, therefore, a lower mortality risk is estimated. In particular, as shown in
Figure 3, the group with the longest quartile LTL (T/S >2.86) had 8.79 median years of survival, which is over 4 years higher than that of the group with the shortest quartile LTL (<1.57 T/S). Furthermore, in a multivariate analysis taking into account the effect of other factors, LTL increased survival and therefore decreased mortality. In detail, for each unit increase in the LTL, the risk of mortality decreased by 22.41%, with the other explanatory variables being equal, age included (
Table 5).
LTL has often been related to life expectancy, but contradictory results have been reported in older adults. Whereas some studies have reported that LTL attrition is a marker of mortality [
30,
31,
32,
33], others have not found such a relationship [
34,
35,
36,
37], making the relationship between LTL and survival in humans uncertain. Those authors that did not find this association mentioned variation in methods of LTL measurements [
35] and variations among investigated populations, including a short age range (>75 years) [
30,
35,
37] and diseases examined, where the elongation of telomeres could be a characteristic of the disease [
36].
4.2. Determinants of Survival
Key survival determinants were not only LTL but also SPPB: for each unit increase in the index, the mortality risk was reduced by 8.78%, considering all other factors being equal, age included (
Table 5). It is known that physical activity is correlated with reduced risks of all cause and cause-specific mortality [
38]. Furthermore, a prospective cohort study showed that middle-aged and older adults achieved considerable longevity advantages by becoming more physically active, regardless of past physical activity [
39]. In our study, we used the SPPB that may be more likely to capture the real health status of older persons. This test, with three different evaluations (walking speed, chair stand and balance time) [
21,
40], was thought an extremely sensitive indicator of global health status or vulnerability, revealing numerous underlying physiological disorders [
41]. Our results are in line with one of the most complete meta-analyses with a proper sample size that definitively discovered the association between SPPB score and all-cause mortality [
42].
On the other hand, the male gender, CCI and age were factors that threatened the survival, with a risk of mortality growing by 74.99% for males, by 16.57% per unit increase in CCI and by 8.5% for each additional year of age (
Table 5). These data confirmed the current literature on gender inequity: men achieve higher mortality rates than women [
43]. Human females’ longevity benefit may derive from the hormonal effect on inflammatory and immunological reactions [
44]. CCI is a validated, simple and easily applicable method for appraising the risk of death and has been broadly applied as a predictor of long-term prognosis and survival [
24,
45]. In our study, CCI score gave an HR of 1.2 (
Table 5) similar to that reported by Fraccaro et al. [
46]; i.e., 1.3 HR in an adult cohort study from Salford (UK) from 2005 to 2014.
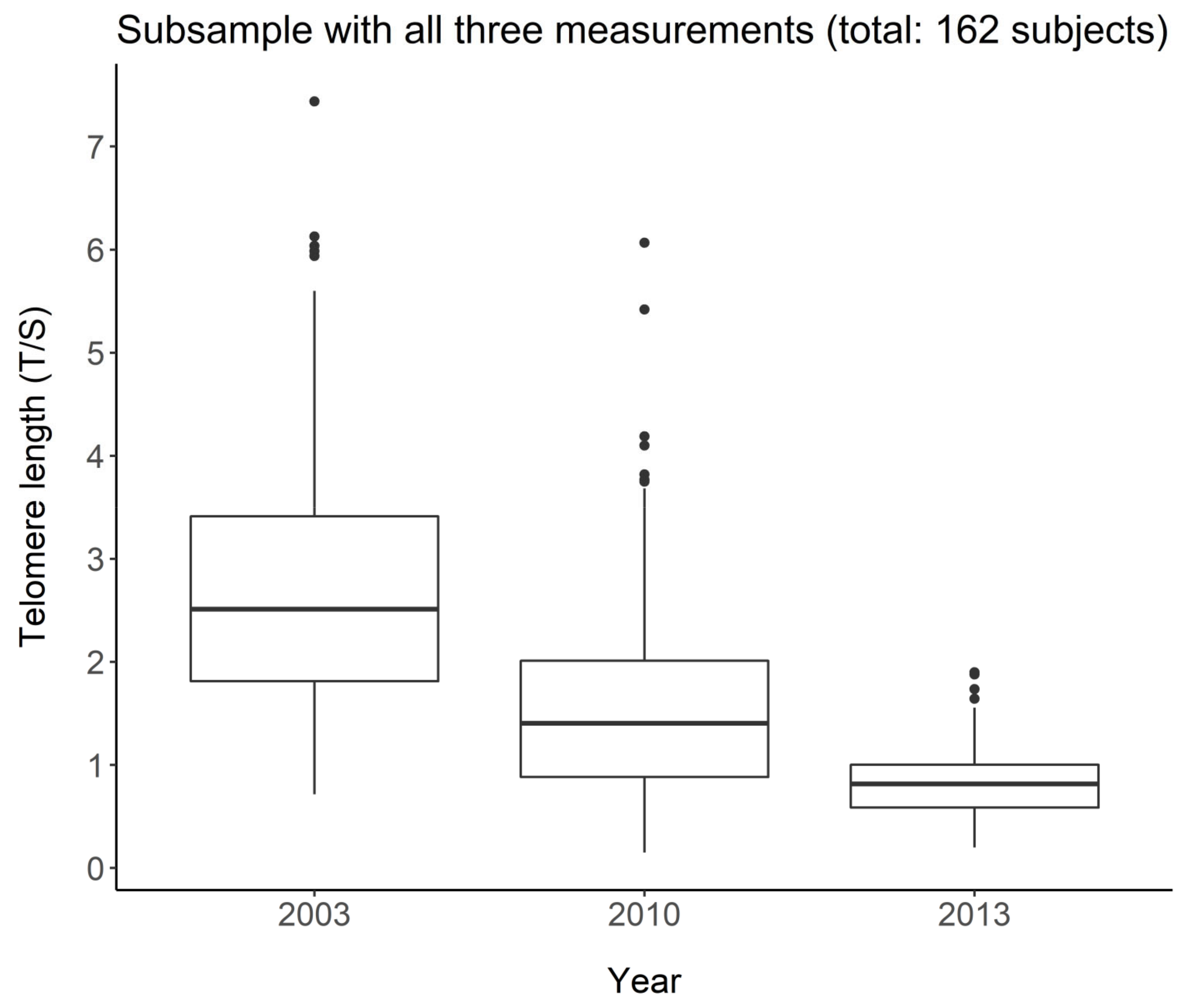
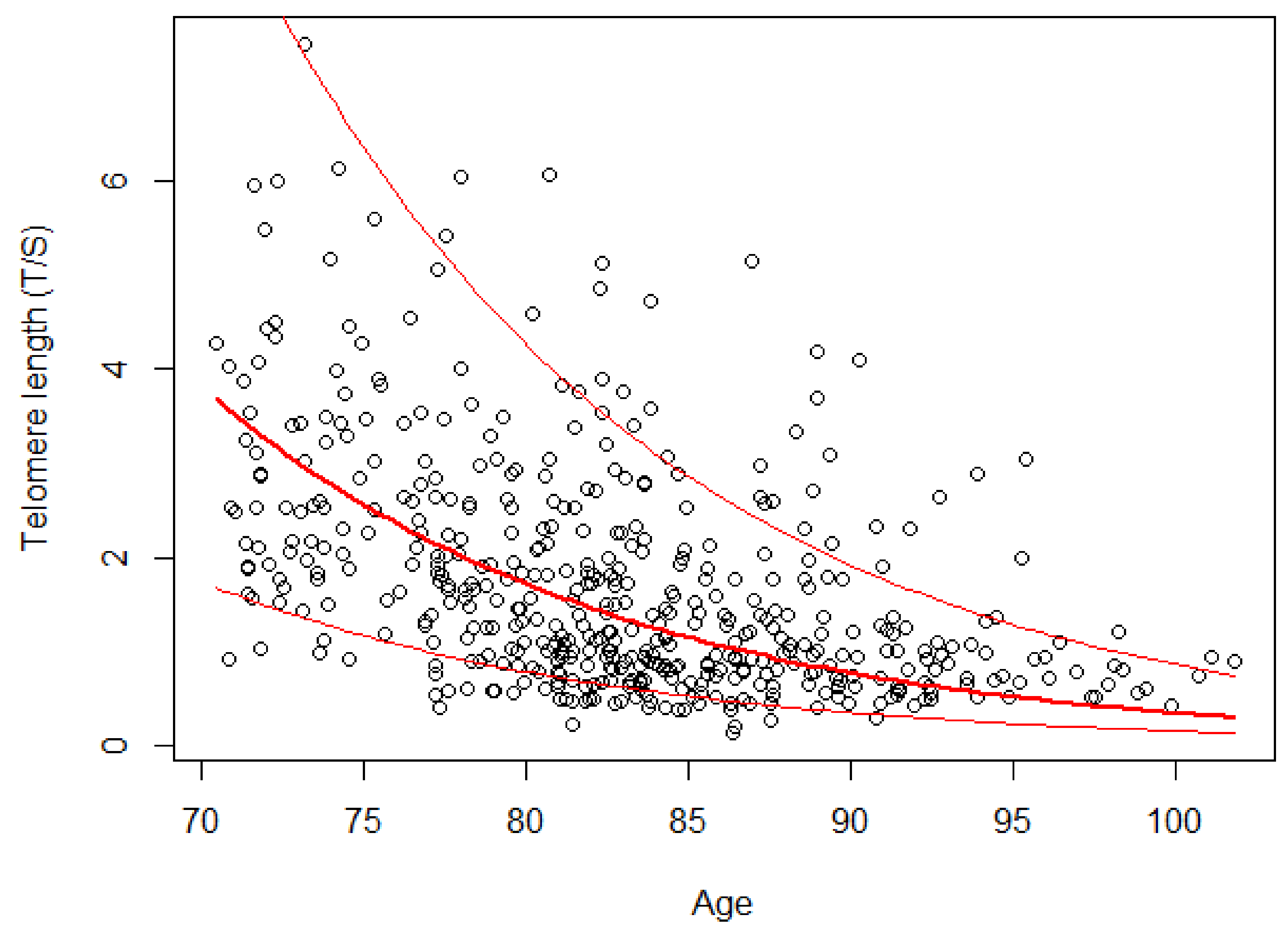
4.3. LTL in the Longitudinal Study
LTL distribution at the baseline in 2003 showed that LTL attrition was related to age (
Table 2). Another key finding, by analyzing LTL at the three waves in the subgroup of 162 subjects, was LTL drop by 7.96% for each additional year of age (
Table 3). The rate of LTL shortening we found was similar to that observed in the longitudinal population-based Bruneck Study (age 45–85 years), in which the telomeres lost on average 455 bp in length after 10 years, corresponding to an average loss of 45.5 bp per year that is equal to about 10% [
47]. An overall lesser decrease (13.6% after 7 years follow-up) than our study was, however, found in a cohort study of 516 subjects aged 65–106 from Southern Italy [
48]. This could be attributed to the major number of centenarians in whose LTL may be expected to increase after 90 years [
49]. Lastly, our LTL drop is comparable to the 7.6% decrease in LTL reported by McCracken et al. [
50] associated with an annual increase in air pollution. It could be speculated that our decrease in LTL may be attributed to the air pollution of the Veneto region, being part of the Pianura Padana, one of the most polluted regions in Europe.
Another noteworthy key finding of our study was the paradoxical increase in LTL in almost 12% of the study population. This finding, with almost exactly the same percentage, has been previously reported [
34]. Loss of telomeric DNA caused by deterioration or incomplete replication seems to be balanced by telomere elongation, which involves the telomerase, a DNA polymerase that synthesizes TTAGGG repeats, inducing the de novo synthesis of additional repeats. This specific reverse transcriptase complex counters the replication-associated telomere shortening with the addition of G-rich telomeric repeats to the chromosome ends, thus efficiently stabilizing TL [
51]. That the gain in LTL is merely a consequence of a high degree of measurement error can be rejected by the accurate quality controls with measurements in triplicate of T/S ratios that resulted in a standard deviation < 0.25 and with a coefficient of variation (CV), on average, less than 9%, in samples analyzed during three successive days, which is comparable to the reproducibility formerly described for this method [
28].
4.4. Determinants of LTL Elongation
Among determinants eliciting LTL elongation were SPPB and years of education, while being male and DCI were factors that contributed to reduced LTL. Until now, few studies have explored the association between LTL and SPPB, with conflicting results. Pereira et al. [
52], in a study with a small number of subjects, reported unexpectedly that shorter LTL was correlated with better physical capability evaluated by SPPB score. No association was found in a cohort of 136 patients aged 45–85 years with knee osteoarthritis pain, in which the SPPB test is considered a clinical measure for assessing physical function [
53]. We can assert that, in general, in observational studies, greater physical activity or exercise was associated with longer LTL in various populations, whereas in a small number of longitudinal studies in which exercise interventions were performed to investigate the potential effect on LTL, such a relationship was not fully established [
54]. It has been reported that physical exercise, affecting the balance between oxidative stress and antioxidants and altering methylation patterns, buffers LTL shortening [
54]. Furthermore, physical training increases telomerase activity in myocytes and circulating mononuclear cells in rats [
55] and in human leukocytes [
55], thus providing another possible and potentially testable pathway to clarify how physical activity protects LTL [
56].
The positive relationship between LTL and years of education agrees with previous findings on a positive association between educational attainment and LTL [
57,
58]. The implication of this finding suggests that education confers health benefits by limiting biological aging. Education, in fact, is one of the leading determinants of overall social status over the course of a person’s lifetime and can reduce exposure to chronic stress and/or increase one’s capacity to effectively deal with potential threats or stressors [
59].
Instead, the role of the male sex as a negative factor for telomere elongation is consistent with the majority of previous cross-sectional and longitudinal studies establishing that women have longer telomeres than men [
60] and that the rate of LTL shortening is slower in women than men [
60]. Several realistic biological reasons can be formulated to elucidate this, including the action of estrogens [
61], which can promote the production of telomerase [
62] and protect against damage caused by reactive oxygen species [
63].
Convincing evidence in the literature suggests the association between shorter LTL and several age-related diseases (such as cardiovascular diseases, neurodegenerative diseases, type 2 diabetes mellitus, premature aging syndromes and cancers) [
30,
64,
65]. Therefore, it is not surprising that the concomitant existence of a higher number of diseases, indicated by a higher DCI, is a risk factor that contributes to reduce LTL.
4.5. Strengths and Limitations
Whether LTL can be considered a predictor for longevity and healthy life is one of the most questioned issues in the field of gerontological research. Problems in results’ interpretation stem from the heterogeneity of factors influencing LTL, including genetics and environment, and in particular from the available follow-up data for both LTL and survival for recruited samples.
Our study was conducted in Treviso, in Veneto, a region of North-East Italy, which is characterized by the highest longevity, in particular among women [
2]. Prevalence of the APOEε4 allele in Treviso is equal to 16.3%, which is lower than the 25% mostly observed for northern European populations without dementia [
26], and this can explain, at least in part, the greater longevity of women in the Treviso area. It should be noted that the Northern population, including Venetians, represents a quite homogeneous population from the genetics point of view, which is different from other populations studied so far in Italy. In the ancestors of Northern Italians, climate-related selective pressures have influenced an adaptive evolution that make Northern Italian people less prone to develop accelerating aging disorders, including diabetes type 2 and obesity [
66].
In our study, we determined LTL during ten-year follow up in the prospective cohort “Treviso Longeva (TRELONG)”, a very well-characterized study population. In particular, this is a systematic sample of subjects aged 70–106 years, drawn from the list of residents of the municipality of Treviso. Therefore, the study sample is highly representative of the community and of the real world [
15]. Furthermore, the cohort is well characterized with several types of information, including physiological variables and lifestyles, physical performance and frailty measures, assessment of chronic disease and other covariates besides biochemical measurements and APOE genotyping [
26].
We used the q-RT PCR technique to assess LTL in triplicate. This fast and highly sensitive method requires relatively small quantities of DNA and allows high throughput. Since an almost totally automated workflow from DNA extraction to LTL analysis was used, pre-analytical factors and analytical factors did not vary amongst analyses and the same pool DNA was used as a reference standard in all q-RT PCR reactions. In this way, the interbach variation was under control, minimizing measurement error and warranting a CV of less than 9%.
Moreover, in this study, the start date of the survey was the day of the interview rather than the date of birth so that the measurements relating to the lengths of the telomeres of the interviewees who had very different ages were comparable. In this case, then, the duration of the episode indicates the remaining survival time for the individual with that LTL.
Another strong point is that the Cox model estimate for time-dependent variables was further improved considering that for all 613 subjects a statistical weight was computed so that the TRELONG study population was representative per gender and age of the Italian population over seventy. Therefore, the results could be more easily extended to the entire population over seventy at the national level. It should be noted, however, that while the sample of the present study was found in a single city, a sample detected at the national level would be affected by the different geographical distribution of the subjects recruited.
5. Conclusions
Our results demonstrate that LTL could be a powerful tool for predicting survival in our elderly population. By identifying SPPB as a key survival determinant, our study suggests that the regular application of the SPPB in clinical practice may offer helpful prognostic information regarding the risk of all-cause mortality. This study also confirms that male gender, CCI, and, therefore chronological age, are threats for survival.
Furthermore, by identifying SPPB scores and years of education as positive and DCI as a negative determinant of LTL elongation, we recognized factors on which to converge to improve prevention strategies.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}