
Gut Microbiota Composition and Its Metabolites in Different Stages of Chronic Kidney Disease

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement of the Study Cohort and Sample Collection
2.2. Metadata and Biochemical Analysis
2.3. Bacterial DNA Extraction
2.4. 16S Ribosomal (r)RNA Gene Sequencing
2.5. Metabolites Extraction
2.6. UPLC-MS/MS Analysis
2.7. Bioinformatic Analysis
2.8. UP-LC-MS/MS Data Preprocessing and Annotation
2.9. Statistical Analysis
3. Results
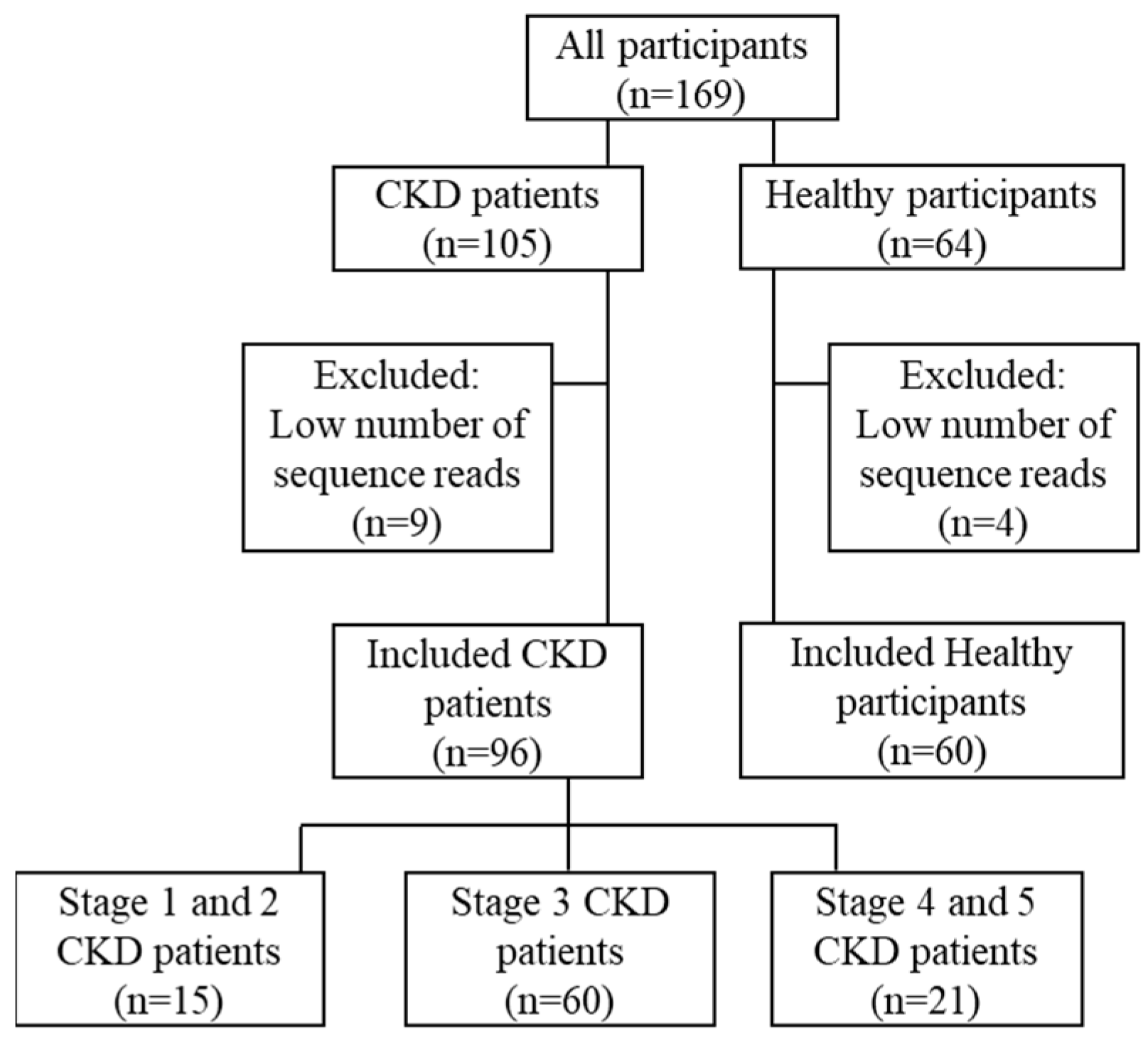
3.1. Demographic Data of Recruited Participants in This Study
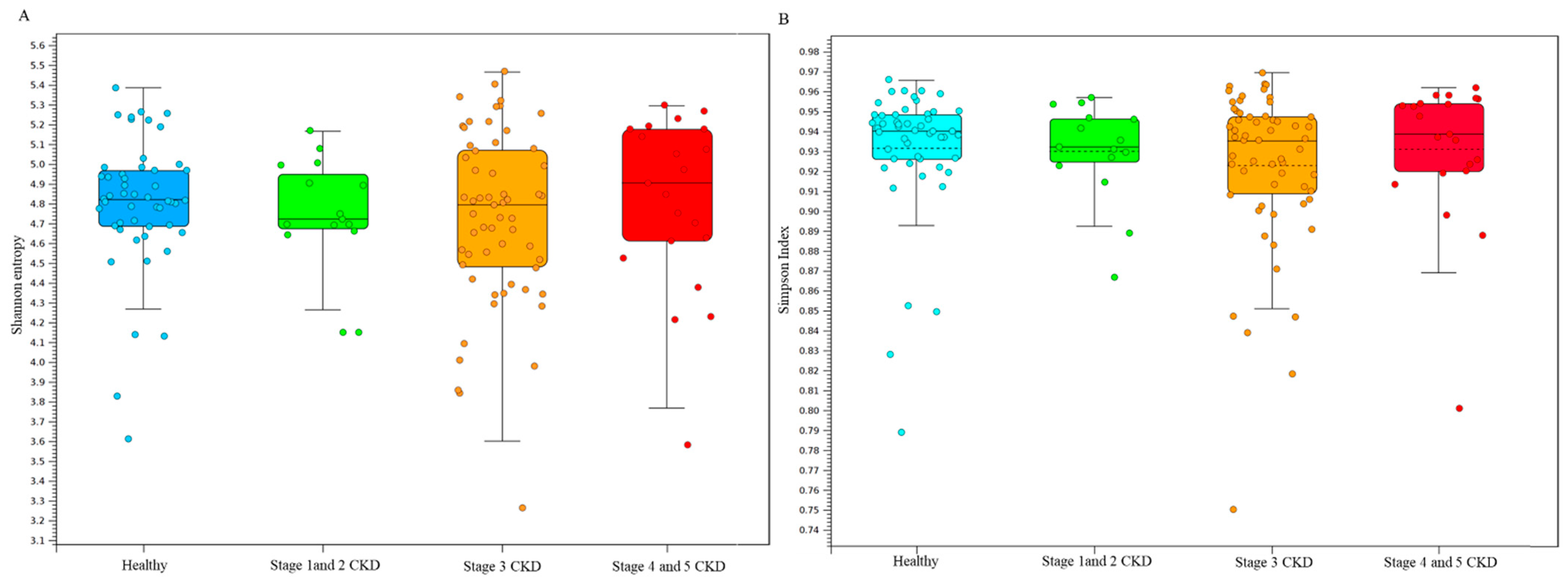
3.2. Statistical Results of Gut Microbial Communities in Enrolled Subjected Assessed with Long-Read Sequencing Results
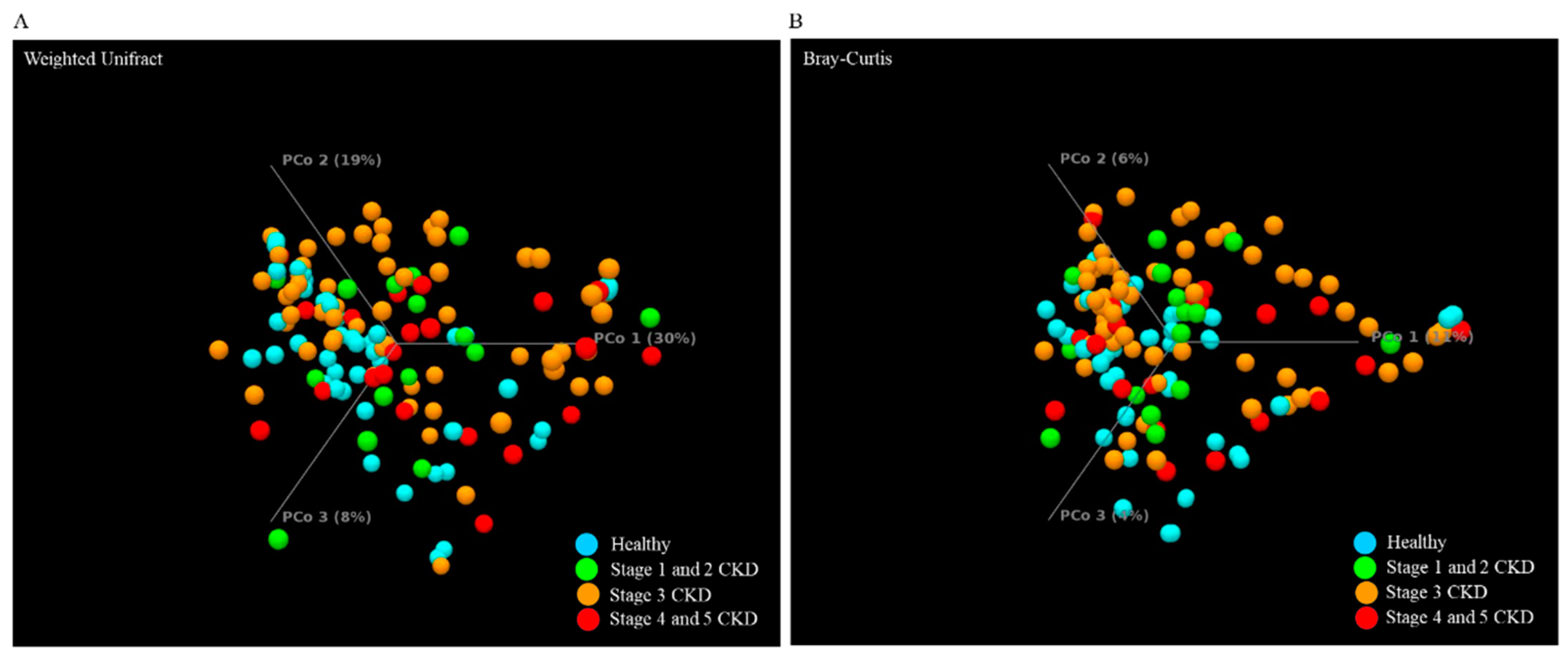
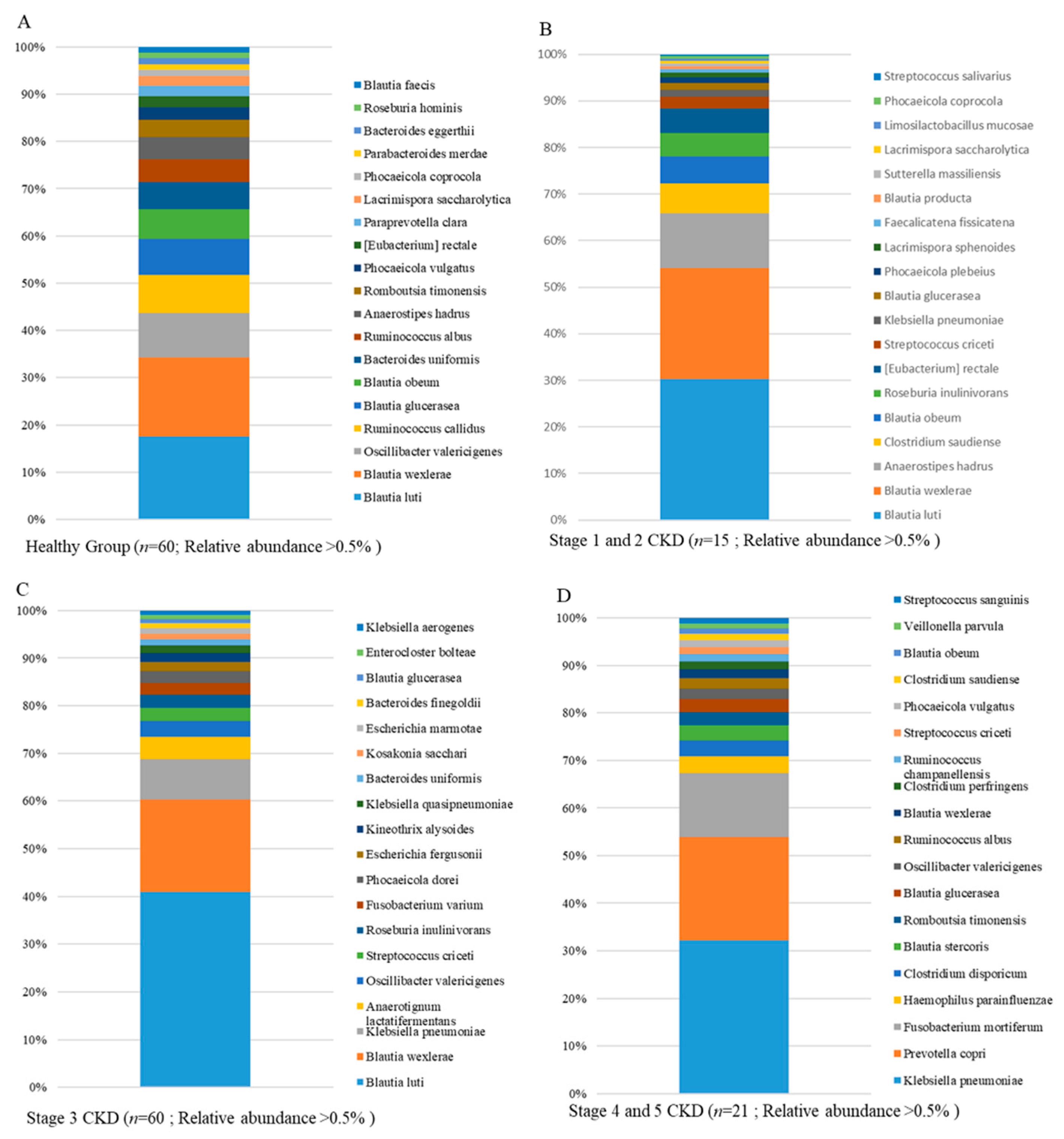
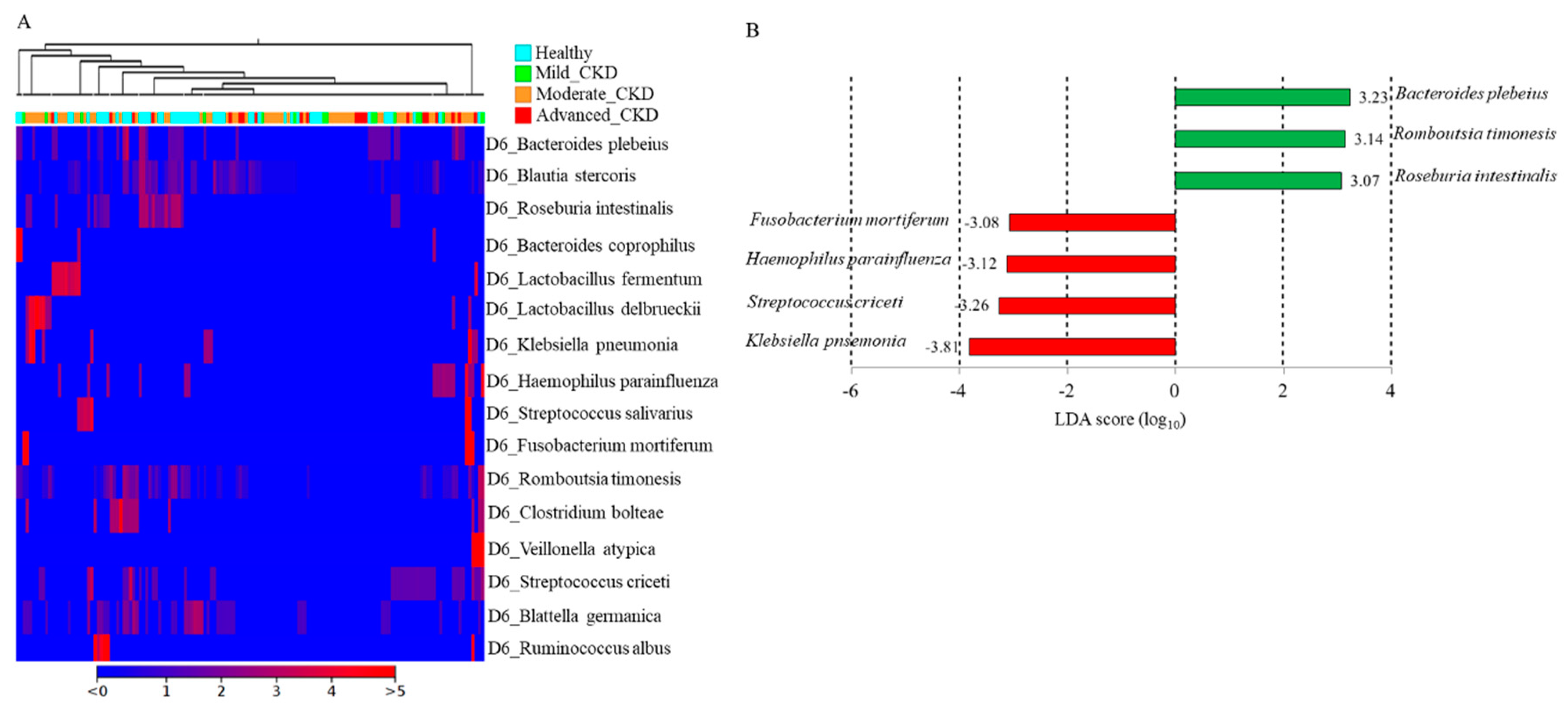
3.3. Identification and Comparison of the Microbial Communities in the Guts of the Healthy Group and CKD Patients Classified Using a Long-Read Sequencing Platform
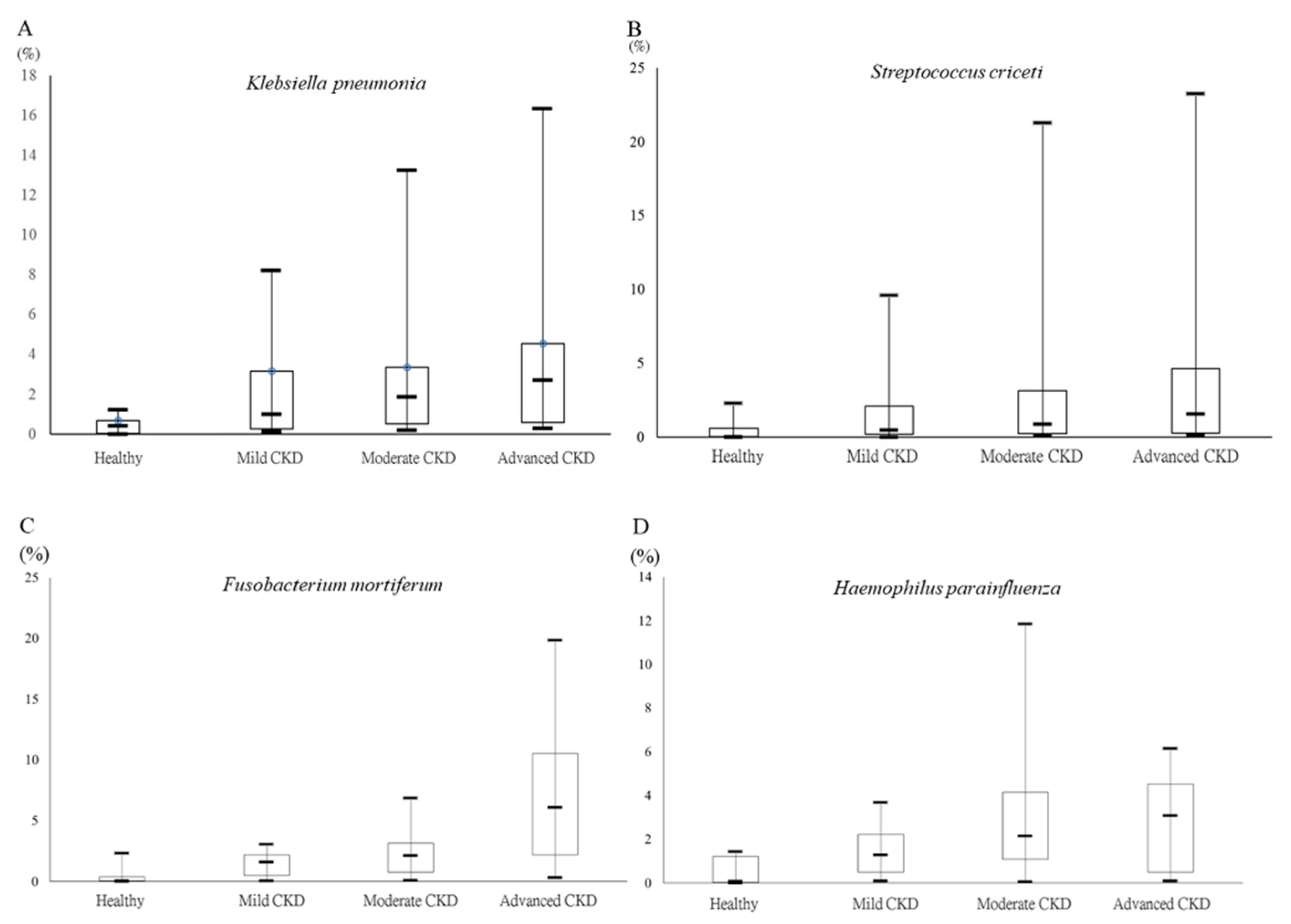
3.4. Differential Abundances of Identified OTUs at the Species Level between Healthy Participants and CKD Patients Evaluated Statistically
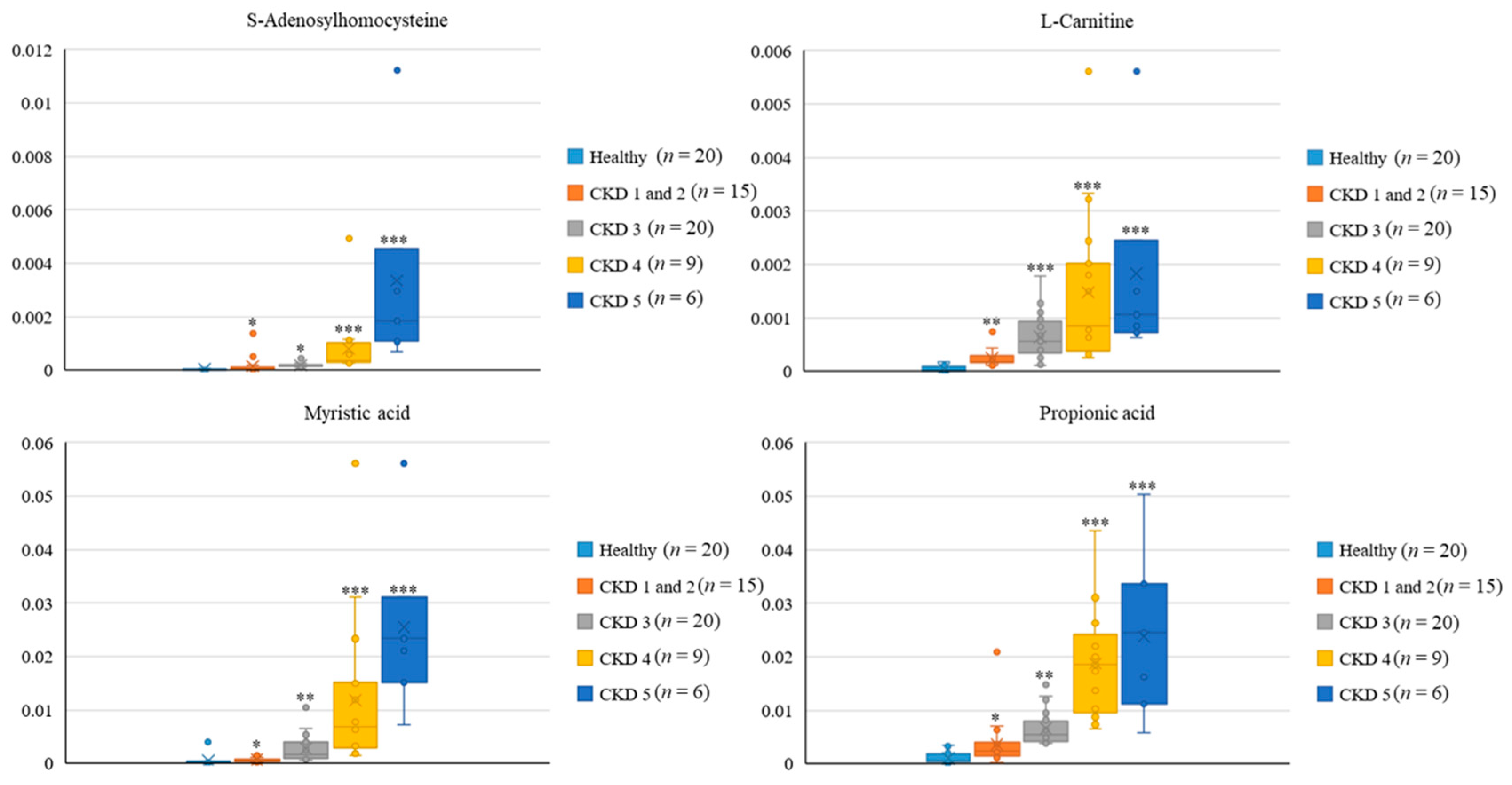
3.5. Metabolic Profiles among Healthy Participants and CKD Patients across Distinct Stages
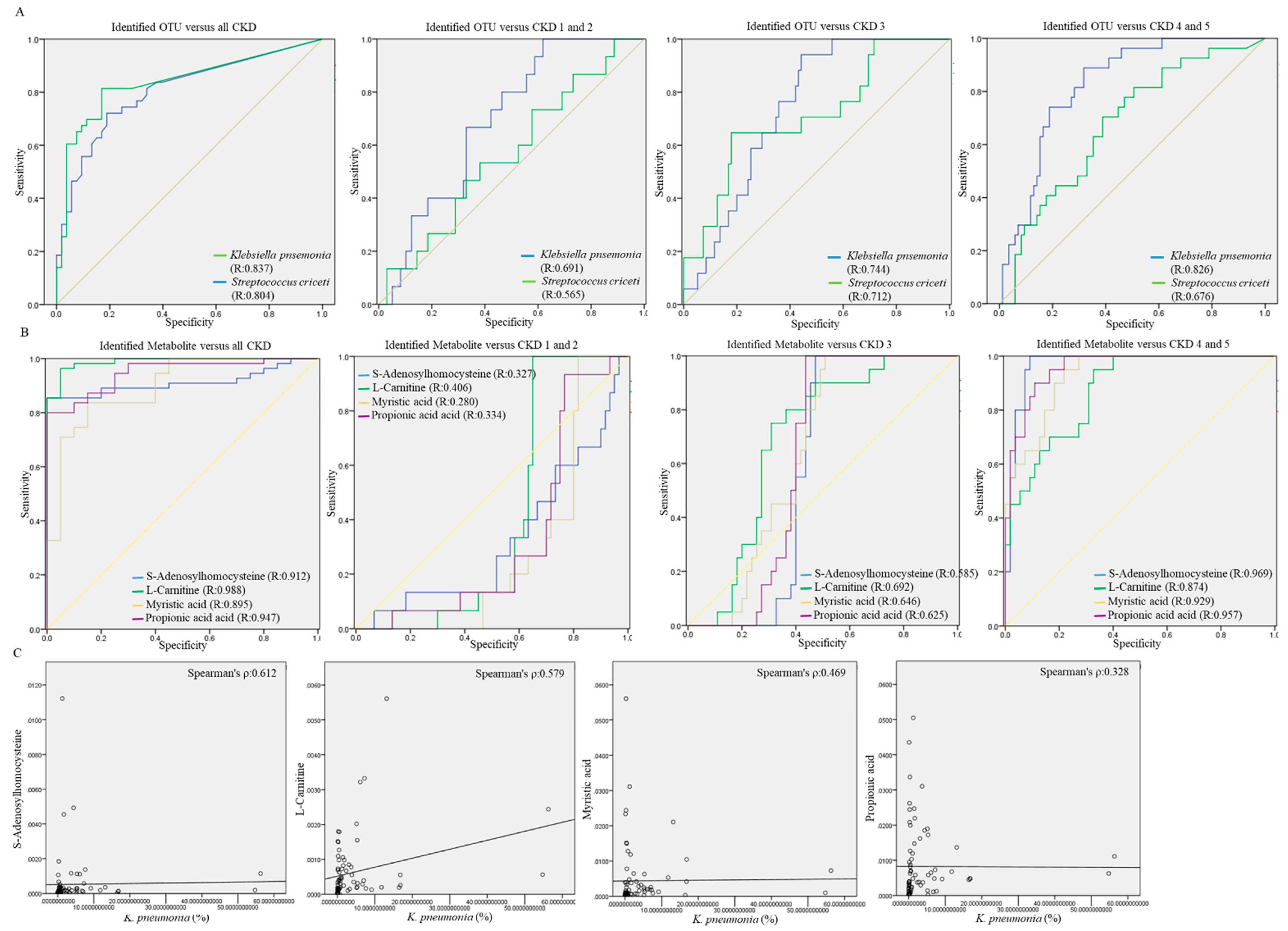
3.6. Potential Utility of Identified OTUs or Gut Metabolites on Differentiating CKD Subjects across Distinct Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, T.K.; Knicely, D.H.; Grams, M.E. Chronic kidney disease diagnosis and management: A review. JAMA 2019, 322, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Kazancioglu, R. Risk factors for chronic kidney disease: An update. Kidney Int. Suppl. 2011, 2013, 368–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taler, S.J.; Agarwal, R.; Bakris, G.L.; Flynn, J.T.; Nilsson, P.M.; Rahman, M.; Sanders, P.W.; Textor, S.C.; Weir, M.R.; Townsend, R.R. KDOQI US commentary on the 2012 KDIGO clinical practice guideline for management of blood pressure in CKD. Am. J. Kidney Dis. 2013, 62, 201–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Zhao, Y.Y.; Pahl, M.V. Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: The nature, mechanisms, consequences and potential treatment. Nephrol. Dial. Transplant. 2016, 31, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenepoel, P.; Poesen, R.; Meijers, B. The gut-kidney axis. Pediatr. Nephrol. 2017, 32, 2005–2014. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; DeSantis, T.Z.; Zhenmin, N.; Nguyen, T.H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Renal Physiol. 2016, 310, F857–F871. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Salgado, J.; Vehaskari, V.M.; Stewart, T.; Ferris, M.; Zhang, Q.; Wang, G.; Blanchard, E.E.; Taylor, C.M.; Kallash, M.; Greenbaum, L.A.; et al. Intestinal microbiota in pediatric patients with end stage renal disease: A Midwest Pediatric Nephrology Consortium study. Microbiome 2016, 4, 50. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Xie, S.; Lv, D.; Wang, P.; He, H.; Zhang, T.; Zhou, Y.; Lin, Q.; Zhou, H.; Jiang, J.; et al. Alteration of the gut microbiota in Chinese population with chronic kidney disease. Sci. Rep. 2017, 7, 2870. [Google Scholar] [CrossRef] [Green Version]
- Kalantar-Zadeh, K.; Fouque, D. Nutritional management of chronic kidney disease. N. Engl. J. Med. 2017, 377, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Iorio, B.R.; Rocchetti, M.T.; De Angelis, M.; Cosola, C.; Marzocco, S.; Di Micco, L.; di Bari, I.; Accetturo, M.; Vacca, M.; Gobbetti, M.; et al. Nutritional therapy modulates intestinal microbiota and reduces serum levels of total and free indoxyl sulfate and p-cresyl sulfate in chronic kidney disease (medika study). J. Clin. Med. 2019, 8, 1424. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Lee, H.Y.; Lin, Y.C. The effect of ketoanalogues on chronic kidney disease deterioration: A meta-analysis. Nutrients 2019, 11, 957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffin, M.R.; Tayebi Khosroshahi, H.; Park, H.; Laffin, L.J.; Madsen, K.; Kafil, H.S.; Abedi, B.; Shiralizadeh, S.; Vaziri, N.D. Amylose resistant starch (HAM-RS2) supplementation increases the proportion of Faecalibacterium bacteria in end-stage renal disease patients: Microbial analysis from a randomized placebo-controlled trial. Hemodial. Int. 2019, 23, 343–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchiyama, K.; Wakino, S.; Irie, J.; Miyamoto, J.; Matsui, A.; Tajima, T.; Itoh, T.; Oshima, Y.; Yoshifuji, A.; Kimura, I.; et al. Contribution of uremic dysbiosis to insulin resistance and sarcopenia. Nephrol. Dial. Transplant. 2020, 35, 1501–1517. [Google Scholar] [CrossRef] [PubMed]
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD Work Group. KDIGO 2012 clinical practice guideline for the evaluation and management of chronic kidney disease. Kidney Int. Suppl. 2013, 3, 1–150. [Google Scholar]
- Wu, I.W.; Gao, S.S.; Chou, H.C.; Yang, H.Y.; Chang, L.C.; Kuo, Y.L.; Dinh, M.C.V.; Chung, W.H.; Yang, C.W.; Lai, H.C.; et al. Integrative metagenomic and metabolomic analyses reveal severity-specific signatures of gut microbiota in chronic kidney disease. Theranostics 2020, 10, 5398–5411. [Google Scholar] [CrossRef]
- Casals-Pascual, C.; González, A.; Vázquez-Baeza, Y.; Song, S.J.; Jiang, L.; Knight, R. Microbial diversity in clinical microbiome studies: Sample size and statistical power considerations. Gastroenterology 2020, 158, 1524–1528. [Google Scholar] [CrossRef]
- Wei, P.L.; Hung, C.S.; Kao, Y.W.; Lin, Y.C.; Lee, C.Y.; Chang, T.H.; Shia, B.C.; Lin, J.C. Characterization of fecal microbiota with clinical specimen using long-read and short-read sequencing platform. Int. J. Mol. Sci. 2020, 21, 7110. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Lee, S.; Go, M.-J.; Lee, S.Y.; Kim, S.C.; Lee, C.H.; Cho, B.-K. Analysis of the mouse gut microbiome using full-length 16S rRNA amplicon sequencing. Sci. Rep. 2016, 6, 29681. [Google Scholar] [CrossRef] [Green Version]
- Shoskes, D.A.; Altemus, J.; Polackwich, A.; Tucky, B.; Wang, H.; Eng, C. The urinary microbiome differs significantly between patients with chronic prostatitis/chronic pelvic pain syndrome and controls as well as between patients with different clinical phenotypes. Urology 2016, 92, 26–32. [Google Scholar] [CrossRef]
- Fan, X.; Jin, Y.; Chen, G.; Ma, X.; Zhang, L. Gut microbiota dysbiosis drives the development of colorectal cancer. Digestion 2020, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Felizardo, R.J.; Castoldi, A.; Andrade-Oliveira, V.; Câmara, N.O.S. The microbiota and chronic kidney diseases: A double-edged sword. Clin. Transl. Immunol. 2016, 5, e86. [Google Scholar] [CrossRef]
- Kanbay, M.; Onal, E.M.; Afsar, B.; Dagel, T.; Yerlikaya, A.; Covic, A.; Vaziri, N.D. The crosstalk of gut microbiota and chronic kidney disease: Role of inflammation, proteinuria, hypertension, and diabetes mellitus. Int. Urol. Nephrol. 2018, 50, 1453–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.; Fan, Y.; Li, A.; Shen, Q.; Wu, J.; Ren, L.; Lu, H.; Ding, S.; Ren, H.; Liu, C.; et al. Alterations of the Human Gut Microbiome in Chronic Kidney Disease. Adv. Sci. 2020, 7, 2001936. [Google Scholar] [CrossRef]
- Peng, J.; Narasimhan, S.; Marchesi, J.R.; Benson, A.; Wong, F.S.; Wen, L. Long term effect of gut microbiota transfer on diabetes development. J. Autoimmun. 2014, 53, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Bosi, E.; Molteni, L.; Radaelli, M.G.; Folini, L.; Fermo, I.; Bazzigaluppi, E.; Piemonti, L.; Pastore, M.R.; Paroni, R. Increased intestinal permeability precedes clinical onset of type 1 diabetes. Diabetologia 2006, 49, 2824–2827. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Roesch, L.F.; Lorca, G.L.; Casella, G.; Giongo, A.; Naranjo, A.; Pionzio, A.M.; Nan, L.; Mai, V.; Wasserfall, C.H.; Schatz, D.; et al. Culture-independent identification of gut bacteria correlated with the onset of diabetes in a rat model. ISME J. 2009, 3, 536–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushpanathan, P.; Srikanth, P.; Seshadri, K.G.; Selvarajan, S.; Pitani, R.S.; Kumar, T.D.; Janarthanan, R. Gut microbiota in type2 diabetes individuals and correlation with monocyte chemoattractant protein1 and interferon gamma from patients attending a Tertiary Care Centre in Chennai, India. Indian J. Endocrinol. Metab. 2016, 20, 523–530. [Google Scholar] [PubMed]
- Cai, L.; Wu, H.; Li, D.; Zhou, K.; Zou, F. Type 2 Diabetes Biomarkers of Human Gut Microbiota Selected via Iterative Sure Independent Screening Method. PLoS ONE 2015, 10, e0140827. [Google Scholar] [CrossRef]
- Brown, C.T.; Davis-Richardson, A.G.; Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; et al. Gut microbiome metagenomics analysis suggests a functional model for the development of autoimmunity for type 1 diabetes. PLoS ONE 2011, 6, e25792. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: Implications for epithelial protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamoto, N.; Sasaki, N.; Aoki, R.; Miyamoto, K.; Suda, W.; Teratani, T.; Suzuki, T.; Koda, Y.; Chu, P.; Tanikiet, N.; et al. Gut pathobionts underlie intestinal barrier dysfunction and liver T helper 17 cell immune response in primary sclerosing cholangitis. Nat. Microbiol. 2019, 4, 492–503. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761. [Google Scholar] [CrossRef] [Green Version]
- Nymark, M.; Pussinen, P.J.; Tuomainen, A.M.; Forsblom, C.; Groop, P.; Lehto, M. Serum lipopolysaccharide activity is associated with the progression of kidney disease in finnish patients with type 1 diabetes. Diabetes Care 2009, 32, 1689–1693. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, A.; Massy, Z.A.; Meijers, B.; Evenepoel, P.; Vanholder, R.; Raj, D.S. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target. Am. J. Kidney Dis. 2016, 67, 483–498. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Zhang, F.; Shen, A.W.; Sun, B.; Xia, T.Y.; Chen, W.S.; Tao, X.; Yu, S.Q. Metabolomics Evaluation of Patients with Stage 5 Chronic Kidney Disease before Dialysis, Maintenance Hemodialysis, and Peritoneal Dialysis. Front. Physiol. 2021, 11, 630646. [Google Scholar] [CrossRef]
- Guo, F.; Dai, Q.; Zeng, X.; Liu, Y.; Tan, Z.; Zhang, H.; Ouyang, D. Renal function is associated with plasma trimethylamine-N-oxide, choline, L-carnitine and betaine: A pilot study. Int. Urol. Nephrol. 2021, 53, 539–551. [Google Scholar] [CrossRef]
- Carron, C.; Pais de Barros, J.P.; Gaiffe, E.; Deckert, V.; Adda-Rezig, H.; Roubiou, C.; Laheurte, C.; Masson, D.; Simula-Faivre, D.; Louvat, P.; et al. End-Stage Renal Disease-Associated Gut Bacterial Translocation: Evolution and Impact on Chronic Inflammation and Acute Rejection After Renal Transplantation. Front. Immunol. 2019, 10, 1630. [Google Scholar] [CrossRef] [Green Version]
- Shchelochkov, O.A.; Manoli, I.; Sloan, J.L.; Ferry, S.; Pass, A.; Van Ryzin, C.; Myles, J.; Schoenfeld, M.; McGuire, P.; Rosing, D.R.; et al. Chronic kidney disease in propionic acidemia. Genet. Med. 2019, 21, 2830–2835. [Google Scholar] [CrossRef]
- Herrmann, W.; Schorr, H.; Obeid, R.; Makowski, J.; Fowler, B.; Kuhlmann, M.K. Disturbed homocysteine and methionine cycle intermediates S-adenosylhomocysteine and S-adenosylmethionine are related to degree of renal insufficiency in type 2 diabetes. Clin. Chem. 2005, 51, 891–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, A.; Carrero, J.J.; Qureshi, A.R.; Garibotto, G.; Bárány, P.; Axelsson, J.; Lindholm, B.; Stenvinkel, P.; Anderstam, B.; Suliman, M.E. Elevated serum levels of S-adenosylhomocysteine, but not homocysteine, are associated with cardiovascular disease in stage 5 chronic kidney disease patients. Clin. Chim. Acta 2008, 395, 106–110. [Google Scholar] [CrossRef]
- Yang, J.; Fang, P.; Yu, D.; Zhang, L.; Zhang, D.; Jiang, X.; Yang, W.Y.; Bottiglieri, T.; Kunapuli, S.P.; Yu, J.; et al. Chronic Kidney Disease Induces Inflammatory CD40+ Monocyte Differentiation via Homocysteine Elevation and DNA Hypomethylation. Circ. Res. 2016, 119, 1226–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornell, K.A.; Winter, R.W.; Tower, P.A.; Riscoe, M.K. Affinity purification of 5-methylthioribose kinase and 5-methylthioadenosine/S-adenosylhomocysteine nucleosidase from Klebsiella pneumoniae. Biochem. J. 1996, 317, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Pranavathiyani, G.; Prava, J.; Rajeev, A.C.; Pan, A. Novel Target Exploration from Hypothetical Proteins of Klebsiella pneumoniae MGH 78578 Reveals a Protein Involved in Host-Pathogen Interaction. Front. Cell Infect. Microbiol. 2020, 10, 109. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Group (n = 60) | Stage 1 and 2 CKD (n = 15) | Stage 3 CKD (n = 60) | Stage 4 and 5 CKD (n = 21) | p Value | |
|---|---|---|---|---|---|
| Ethnicity (Taiwanese) | 60 (100%) | 15 (100%) | 60 (100%) | 21 (100%) | No Difference |
| Age (Median(IQR)) | 66 (41–87) | 67 (50–84) | 71 (33–90) | 71 (43–87) | >0.5 |
| Sex Female Male | >0.5 | ||||
| 32 (53.3%) 28 (46.7%) | 4 (26.67%) 11 (73.33%) | 30 (50%) 30 (50%) | 10 (47.62%) 11 (52.38%) | ||
| T2DM | 0 (0%) | 6 (40%) | 9 (15%) | 7 (33.3%) | >0.5 |
| Fasting Blood Glucose (mg/dL) (Median(IQR)) | 89 (61–100) | 114 (91–425) | 101 (81–216) | 108 (77–278) | >0.5 |
| PleaseHbA1c (%) (Median(IQR)) | 5.1 (4.2–6.0) | 7.05 (5.2–9.4) | 5.8 (4.1–8.6) | 6 (4.7–8.5) | >0.5 |
| Serum Creatinine (mg/dL) (Median(IQR)) | 0.72 (0.5–1.15) | 1.05 (0.73–1.41) | 1.475 (0.86–3.36) | 3.12 (1.96–13.15) | 0.031 |
| eGFR (ml/min/1.73 m2) (Median(IQR)) | 92.4 (63.9–134.2) | 69.88 (63.57–81.18) | 43.37 (37.32–51.7) | 18.11 (10.04–25.73) | 0.018 |
| Group | Healthy Group (n = 60) | Stage 1 and 2 CKD (n = 15) | Stage 3 CKD (n = 60) | Stage 4 and 5 CKD (n = 21) | p Value |
|---|---|---|---|---|---|
| Number of Raw reads per sample | 86,121 (±7321) | 101,233 (±15,702) | 91,775 (±13,488) | 113,206 (±23,517) | >0.5 |
| Number of qualified reads per sample | 63,879 (±5121) | 93,508 (±8332) | 80,034 (±7540) | 96,681 (±8116) | >0.5 |
| Correctly classified (% (SD)) | 89.22 (5.64) | 90.13 (4.97) | 91.59 (5.06) | 87.31 (7.21) | >0.5 |
| Metabolite | KEGG | HMDB | Microbe | Hsa | Healthy | CKD | VIP | p Value | Fold Change | RSD (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| S-Adenosylhomocysteine | C00021 | HMDB0000939 | + | + | 1.81 × 10−5 | 0.000258 | 2.51 | 0.029 | 6.94 | 3.37 |
| Propionic acid | C00163 | HMDB0000619 | + | + | 0.000239 | 0.012946 | 2.36 | 0.021 | 3.17 | 23.17 |
| Myristic acid | C06424 | HMDB0000806 | + | + | 0.005311 | 0.001116 | 2.35 | 0.019 | 3.49 | 9.85 |
| L-Carnitine | C00318 | HMDB0000062 | NA | + | 7.44 × 10−6 | 0.000509 | 2.25 | 0.131 | 2.72 | 19.38 |
| Capsaicin | C06866 | HMDB0002227 | + | + | 0.000343 | 0.002697 | 1.75 | 0.034 | 6.02 | 11.52 |
| L-Tyrosine | C00082 | HMDB0000158 | + | + | 0.000186 | 0.005451 | 1.74 | 0.037 | 4.81 | 24.62 |
| Ephedrine | C01575 | HMDB0015451 | + | + | 4.51 × 10−6 | 0.000865 | 2.03 | 0.019 | 6.25 | 17.22 |
| gamma-Terpinene | C09900 | HMDB0005806 | NA | NA | 0.000131 | 0.001716 | 1.39 | 0.039 | 4.43 | 20.15 |
| Tricetin | C10192 | HMDB0029620 | NA | NA | 0.120803 | 0.000461 | 1.27 | 0.124 | 3.09 | 20.54 |
| Trehalose | C01083 | HMDB0000975 | + | + | 0.000273 | 0.02608 | 1.21 | 0.042 | 6.18 | 24.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.-H.; Liu, C.-W.; Ho, Y.-H.; Huang, C.-K.; Hung, C.-S.; Smith, B.H.; Lin, J.-C. Gut Microbiota Composition and Its Metabolites in Different Stages of Chronic Kidney Disease. J. Clin. Med. 2021, 10, 3881. https://doi.org/10.3390/jcm10173881
Chen T-H, Liu C-W, Ho Y-H, Huang C-K, Hung C-S, Smith BH, Lin J-C. Gut Microbiota Composition and Its Metabolites in Different Stages of Chronic Kidney Disease. Journal of Clinical Medicine. 2021; 10(17):3881. https://doi.org/10.3390/jcm10173881
Chicago/Turabian StyleChen, Tso-Hsiao, Chao-Wei Liu, Yi-Hsien Ho, Chun-Kai Huang, Ching-Sheng Hung, Barry H. Smith, and Jung-Chun Lin. 2021. "Gut Microbiota Composition and Its Metabolites in Different Stages of Chronic Kidney Disease" Journal of Clinical Medicine 10, no. 17: 3881. https://doi.org/10.3390/jcm10173881
APA StyleChen, T.-H., Liu, C.-W., Ho, Y.-H., Huang, C.-K., Hung, C.-S., Smith, B. H., & Lin, J.-C. (2021). Gut Microbiota Composition and Its Metabolites in Different Stages of Chronic Kidney Disease. Journal of Clinical Medicine, 10(17), 3881. https://doi.org/10.3390/jcm10173881