
Caries Management—The Role of Surface Interactions in De- and Remineralization-Processes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction: The Role of Interfacial Phenomena in Cariology
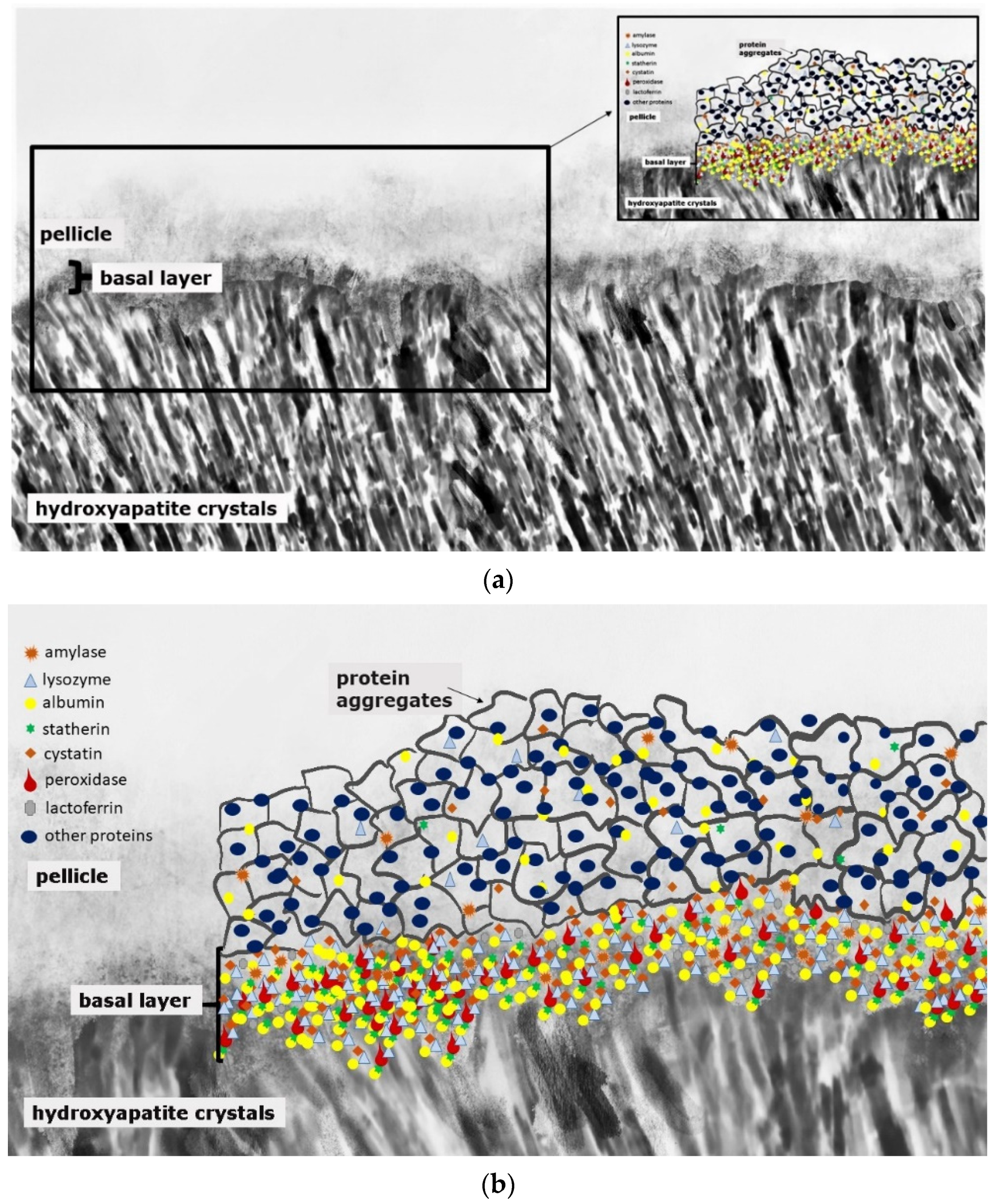
2. From Initial Biofilm to Initial Carious Lesion
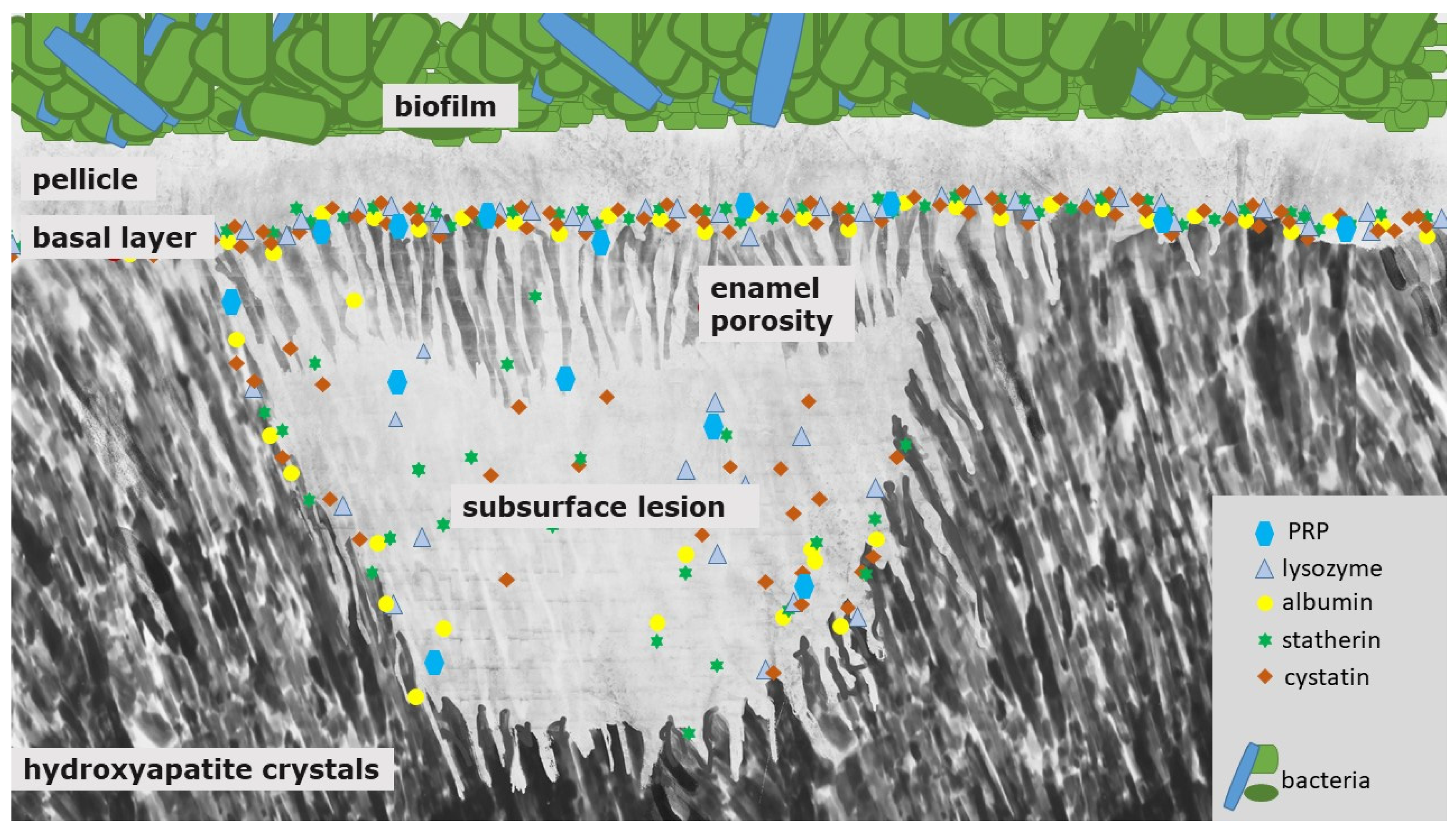
3. Proteins in Initial Carious and in Initial Erosive Lesions—The Subsurface Pellicle
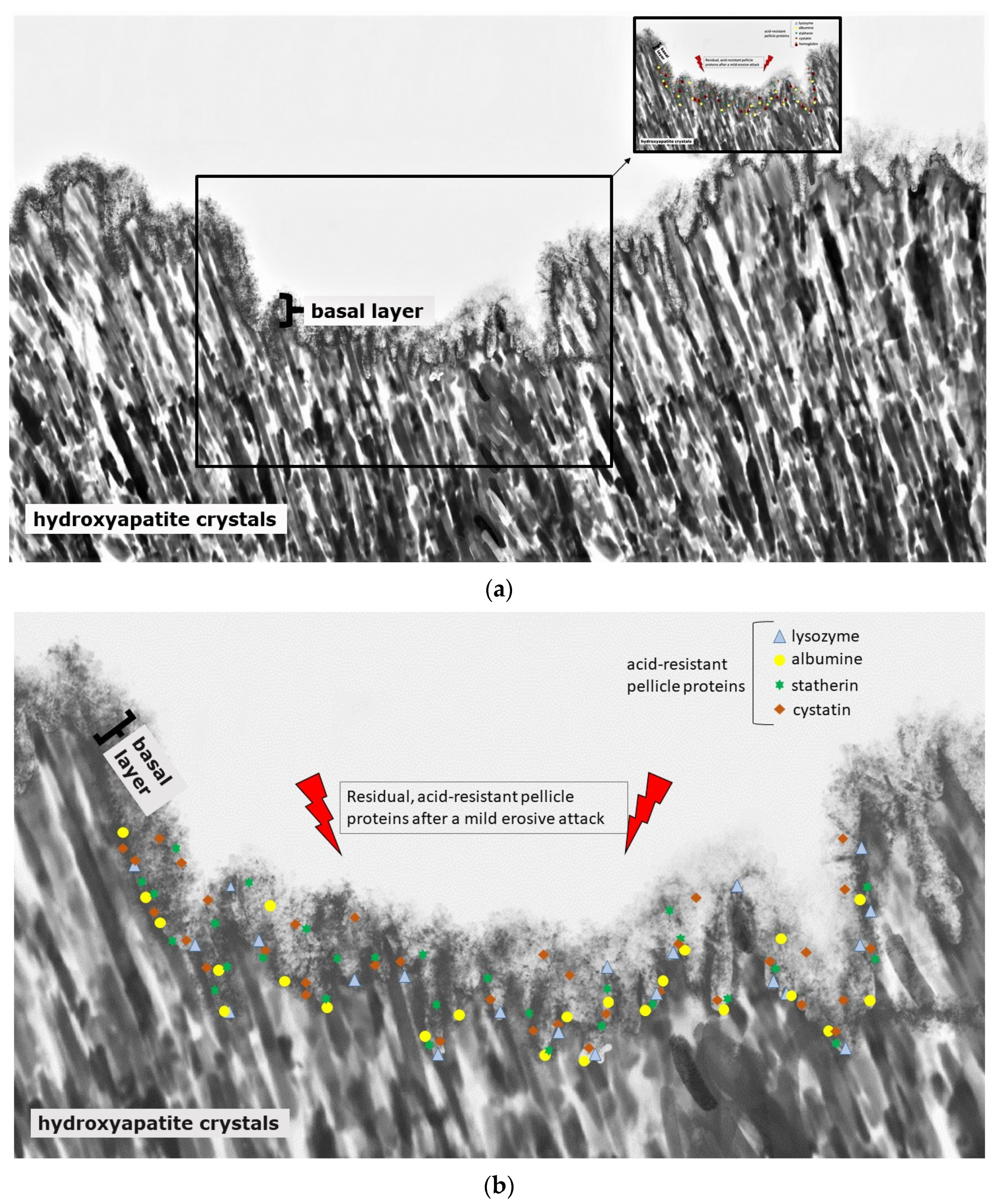
4. Differences between Erosion and Carious Lesions in De- and Remineralization
5. Proteins in the Oral Cavity of Caries-Active and -Inactive Subjects
5.1. Differences in Salivary Protein Composition in Caries-Active and -Inactive Individuals
5.2. Differences of Pellicle Protein Composition in Caries-Active and -Inactive Individuals
5.3. Qualitative Pellicle Proteome Composition in Caries-Active and -Inactive Individuals
5.4. Quantitative Pellicle Proteome Composition in Caries-Active and -Inactive Individuals
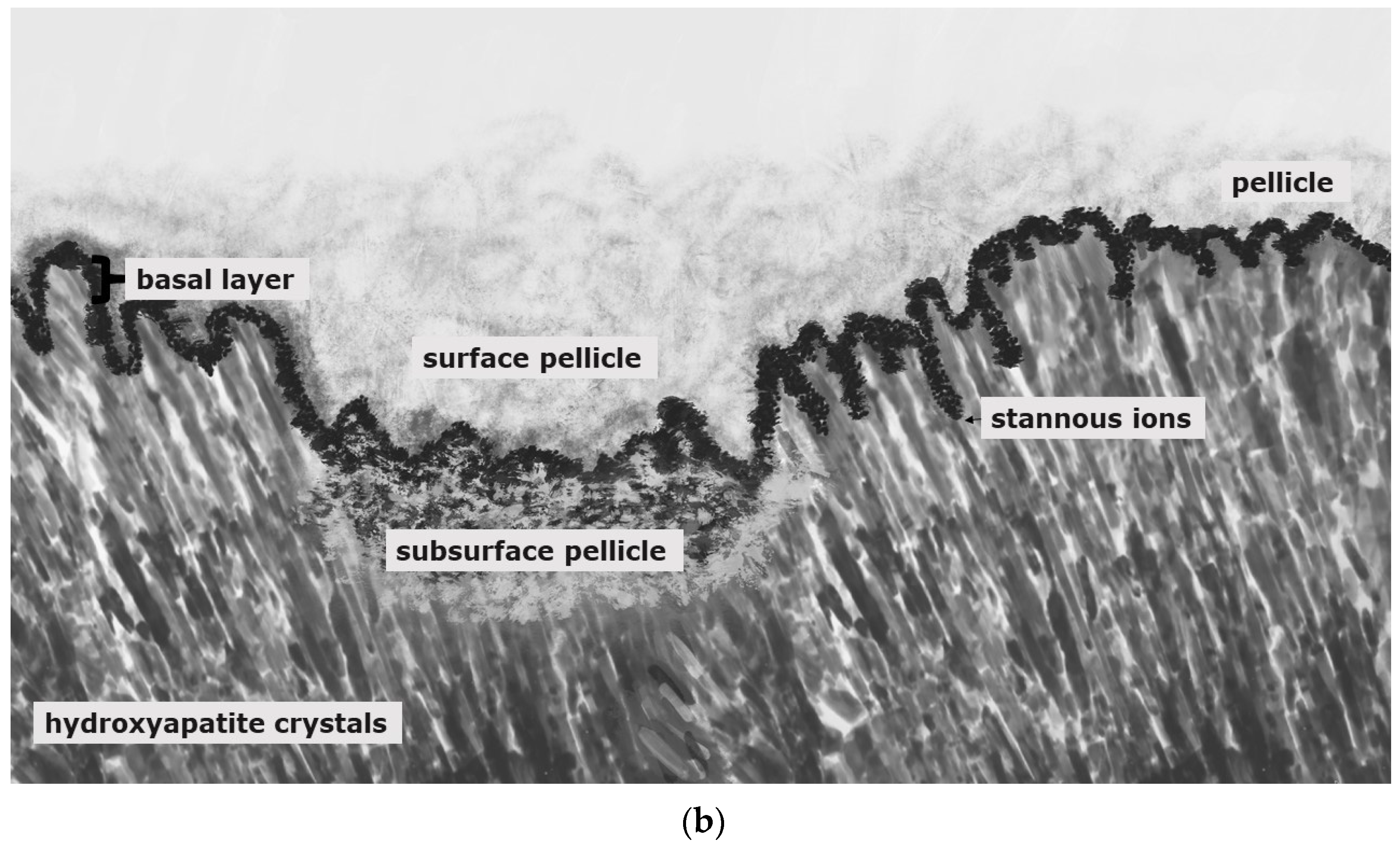
6. Gold Standard in Preventive Dentistry—Fluorides—Models for Preventive Effects
7. Prevention of Demineralization
8. Impact of Proteins on De- and Remineralization
9. Promotion of Remineralization
10. Topics for Future Research
- -
- What is the diffusion time and characteristics of single proteins such as PrP, statherin and ions, like stannous ions, into carious lesions? How do pore sizes and lesion progression influence these diffusion processes?
- -
- How can proteins close the pores of carious lesions? Which physicochemical processes might lead to such a sealing?
- -
- Which protein content can be found in initial and severe carious lesions with proteome analyses?
- -
- Which protein profile can be found in a subsurface lesion?
- -
- What are the interactions between fluorides and proteins as well as fluoride-bound cations with proteins?
- -
- How do engineered proteins and peptides affect preventive strategies in situ and in vivo?
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tulek, A.; Mulic, A.; Runningen, M.; Lillemo, J.; Utheim, T.P.; Khan, Q.; Sehic, A. Genetic Aspects of Dental Erosive Wear and Dental Caries. Int. J. Dent. 2021, 2021, 5566733. [Google Scholar] [CrossRef]
- Paris, S.; Banerjee, A.; Bottenberg, P.; Breschi, L.; Campus, G.; Doméjean, S.; Ekstrand, K.; Giacaman, R.A.; Haak, R.; Hannig, M.; et al. How to Intervene in the Caries Process in Older Adults: A Joint ORCA and EFCD Expert Delphi Consensus Statement. Caries Res. 2020, 54, 459–465. [Google Scholar] [CrossRef]
- Trautmann, S.; Barghash, A.; Fecher-Trost, C.; Schalkowsky, P.; Hannig, C.; Kirsch, J.; Rupf, S.; Keller, A.; Helms, V.; Hannig, M. Proteomic Analysis of the Initial Oral Pellicle in Caries-Active and Caries-Free Individuals. Proteom. Clin. Appl. 2019, 13, e1800143. [Google Scholar] [CrossRef]
- Kirsch, J.; Hannig, C.; Pötschke, S.; Basche, S.; Bowen, W.H.; Rupf, S.; Trautmann, S.; Umanskaya, N.; Hannig, M. Enzymology and Ultrastructure of the in situ Pellicle in Caries-Active and Caries-Inactive Patients. Caries Res. 2017, 51, 109–118. [Google Scholar] [CrossRef]
- Kidd, E.A.; Fejerskov, O. What constitutes dental caries? Histopathology of carious enamel and dentin related to the action of cariogenic biofilms. J. Dent. Res. 2004, 83, C35–C38. [Google Scholar] [CrossRef]
- Hannig, M.; Hannig, C. The pellicle and erosion. Monogr. Oral Sci. 2014, 25, 206–214. [Google Scholar] [CrossRef]
- Marsh, P.D.; Zaura, E. Dental biofilm: Ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44 (Suppl. 18), S12–S22. [Google Scholar] [CrossRef]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Mark Welch, J.L.; Dewhirst, F.E.; Borisy, G.G. Biogeography of the Oral Microbiome: The Site-Specialist Hypothesis. Annu. Rev. Microbiol. 2019, 73, 335–358. [Google Scholar] [CrossRef]
- Sterzenbach, T.; Helbig, R.; Hannig, C.; Hannig, M. Bioadhesion in the oral cavity and approaches for biofilm management by surface modifications. Clin. Oral Investig. 2020, 24, 4237–4260. [Google Scholar] [CrossRef]
- Lendenmann, U.; Grogan, J.; Oppenheim, F.G. Saliva and dental pellicle—A review. Adv. Dent. Res. 2000, 14, 22–28. [Google Scholar] [CrossRef]
- Trautmann, S.; Künzel, N.; Fecher-Trost, C.; Barghash, A.; Schalkowsky, P.; Dudek, J.; Delius, J.; Helms, V.; Hannig, M. Deep Proteomic Insights into the Individual Short-Term Pellicle Formation on Enamel-An In Situ Pilot Study. Proteom. Clin. Appl. 2020, 14, e1900090. [Google Scholar] [CrossRef]
- Delius, J.; Trautmann, S.; Medard, G.; Kuster, B.; Hannig, M.; Hofmann, T. Label-free quantitative proteome analysis of the surface-bound salivary pellicle. Colloids Surf. B Biointerfaces 2017, 152, 68–76. [Google Scholar] [CrossRef]
- Hannig, M. Transmission electron microscopic study of in vivo pellicle formation on dental restorative materials. Eur. J. Oral Sci. 1997, 105, 422–433. [Google Scholar] [CrossRef]
- Schweigel, H.; Wicht, M.; Schwendicke, F. Salivary and pellicle proteome: A datamining analysis. Sci. Rep. 2016, 6, 38882. [Google Scholar] [CrossRef]
- Chawhuaveang, D.D.; Yu, O.Y.; Yin, I.X.; Lam, W.Y.; Mei, M.L.; Chu, C.H. Acquired salivary pellicle and oral diseases: A literature review. J. Dent. Sci. 2021, 16, 523–529. [Google Scholar] [CrossRef]
- Vukosavljevic, D.; Hutter, J.L.; Helmerhorst, E.J.; Xiao, Y.; Custodio, W.; Zaidan, F.C.; Oppenheim, F.G.; Siqueira, W.L. Nanoscale adhesion forces between enamel pellicle proteins and hydroxyapatite. J. Dent. Res. 2014, 93, 514–519. [Google Scholar] [CrossRef]
- Baron, A.; DeCarlo, A.; Featherstone, J. Functional aspects of the human salivary cystatins in the oral environment. Oral Dis. 1999, 5, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, I.E.; Lindh, L.; Elofsson, U.; Arnebrant, T. Studies on the exchange of early pellicle proteins by mucin and whole saliva. J. Colloid Interface Sci. 2008, 321, 52–59. [Google Scholar] [CrossRef]
- Lee, Y.H.; Zimmerman, J.N.; Custodio, W.; Xiao, Y.; Basiri, T.; Hatibovic-Kofman, S.; Siqueira, W.L. Proteomic evaluation of acquired enamel pellicle during in vivo formation. PLoS ONE 2013, 8, e67919. [Google Scholar] [CrossRef]
- Mandel, I.D.; Bennick, A. Quantitation of human salivary acidic proline-rich proteins in oral diseases. J. Dent. Res. 1983, 62, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Iontcheva, I.; Oppenheim, F.G.; Offner, G.D.; Troxler, R.F. Molecular mapping of statherin- and histatin-binding domains in human salivary mucin MG1 (MUC5B) by the yeast two-hybrid system. J. Dent. Res. 2000, 79, 732–739. [Google Scholar] [CrossRef]
- Iontcheva, I.; Oppenheim, F.G.; Troxler, R.F. Human salivary mucin MG1 selectively forms heterotypic complexes with amylase, proline-rich proteins, statherin, and histatins. J. Dent. Res. 1997, 76, 734–743. [Google Scholar] [CrossRef]
- Hu, H.; Burrow, M.F.; Leung, W.K. A systematic review of the proteomic profile of in vivo acquired enamel pellicle on permanent teeth. J. Dent. 2021, 113, 103799. [Google Scholar] [CrossRef]
- Ruhl, S.; Sandberg, A.L.; Cisar, J.O. Salivary Receptors for the Proline-rich Protein-binding and Lectin-like Adhesins of Oral Actinomyces and Streptococci. J. Dent. Res. 2004, 83, 505–510. [Google Scholar] [CrossRef]
- Cisar, J.O.; Takahashi, Y.; Ruhl, S.; Donkersloot, J.A.; Sandberg, A.L. Specific inhibitors of bacterial adhesion: Observations from the study of gram-positive bacteria that initiate biofilm formation on the tooth surface. Adv. Dent. Res. 1997, 11, 168–175. [Google Scholar] [CrossRef]
- Rupf, S.; Laczny, C.C.; Galata, V.; Backes, C.; Keller, A.; Umanskaya, N.; Erol, A.; Tierling, S.; Lo Porto, C.; Walter, J.; et al. Comparison of initial oral microbiomes of young adults with and without cavitated dentin caries lesions using an in situ biofilm model. Sci. Rep. 2018, 8, 14010. [Google Scholar] [CrossRef]
- Heller, D.; Helmerhorst, E.J.; Gower, A.C.; Siqueira, W.L.; Paster, B.J.; Oppenheim, F.G. Microbial Diversity in the Early In Vivo-Formed Dental Biofilm. Appl. Environ. Microbiol. 2016, 82, 1881–1888. [Google Scholar] [CrossRef]
- Aas, J.A.; Griffen, A.L.; Dardis, S.R.; Lee, A.M.; Olsen, I.; Dewhirst, F.E.; Leys, E.J.; Paster, B.J. Bacteria of dental caries in primary and permanent teeth in children and young adults. J. Clin. Microbiol. 2008, 46, 1407–1417. [Google Scholar] [CrossRef]
- Chhour, K.L.; Nadkarni, M.A.; Byun, R.; Martin, F.E.; Jacques, N.A.; Hunter, N. Molecular analysis of microbial diversity in advanced caries. J. Clin. Microbiol. 2005, 43, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Munson, M.A.; Banerjee, A.; Watson, T.F.; Wade, W.G. Molecular analysis of the microflora associated with dental caries. J. Clin. Microbiol. 2004, 42, 3023–3029. [Google Scholar] [CrossRef] [PubMed]
- Wolff, D.; Frese, C.; Maier-Kraus, T.; Krueger, T.; Wolff, B. Bacterial biofilm composition in caries and caries-free subjects. Caries Res. 2013, 47, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Woelber, J.P.; Bremer, K.; Vach, K.; König, D.; Hellwig, E.; Ratka-Kruger, P.; Al-Ahmad, A.; Tennert, C. An oral health optimized diet can reduce gingival and periodontal inflammation in humans—A randomized controlled pilot study. BMC Oral Health 2016, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Inchingolo, A.D.; Malcangi, G.; Semjonova, A.; Inchingolo, A.M.; Patano, A.; Coloccia, G.; Ceci, S.; Marinelli, G.; Di Pede, C.; Ciocia, A.M.; et al. Oralbiotica/Oralbiotics: The Impact of Oral Microbiota on Dental Health and Demineralization: A Systematic Review of the Literature. Children 2022, 9, 1014. [Google Scholar] [CrossRef]
- Karatan, E.; Watnick, P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. MMBR 2009, 73, 310–347. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. MMBR 2014, 78, 510–543. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Falsetta, M.L.; Klein, M.I. The exopolysaccharide matrix: A virulence determinant of cariogenic biofilm. J. Dent. Res. 2013, 92, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Tatevossian, A. Facts and artefacts in research on human dental plaque fluid. J. Dent. Res. 1990, 69, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Yamada, K.M. Dynamic cell-matrix interactions modulate microbial biofilm and tissue 3D microenvironments. Curr. Opin. Cell Biol. 2016, 42, 102–112. [Google Scholar] [CrossRef]
- Tanner, A.C.R.; Kressirer, C.A.; Rothmiller, S.; Johansson, I.; Chalmers, N.I. The Caries Microbiome: Implications for Reversing Dysbiosis. Adv. Dent. Res. 2018, 29, 78–85. [Google Scholar] [CrossRef]
- Takahashi, N. Oral Microbiome Metabolism: From “Who Are They?” to “What Are They Doing?”. J. Dent. Res. 2015, 94, 1628–1637. [Google Scholar] [CrossRef]
- Castiblanco, G.A.; Rutishauser, D.; Ilag, L.L.; Martignon, S.; Castellanos, J.E.; Mejía, W. Identification of proteins from human permanent erupted enamel. Eur. J. Oral Sci. 2015, 123, 390–395. [Google Scholar] [CrossRef]
- Gil-Bona, A.; Bidlack, F.B. Tooth Enamel and its Dynamic Protein Matrix. Int. J. Mol. Sci. 2020, 21, 4458. [Google Scholar] [CrossRef]
- Robinson, C.; Hallsworth, A.S.; Shore, R.C.; Kirkham, J. Effect of surface zone deproteinisation on the access of mineral ions into subsurface carious lesions of human enamel. Caries Res. 1990, 24, 226–230. [Google Scholar] [CrossRef]
- Shellis, R.P.; Hallsworth, A.S.; Kirkham, J.; Robinson, C. Organic material and the optical properties of the dark zone in caries lesions of enamel. Eur. J. Oral Sci. 2002, 110, 392–395. [Google Scholar] [CrossRef]
- Shore, R.C.; Kirkham, J.; Brookes, S.J.; Wood, S.R.; Robinson, C. Distribution of exogenous proteins in caries lesions in relation to the pattern of demineralisation. Caries Res. 2000, 34, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Teranaka, T.; Koulourides, T.; Butler, W.T. Protein content and amino-acid content of consolidated carious lesions in human enamel and of experimental lesions in bovine enamel exposed to the human mouth. Arch. Oral Biol. 1986, 31, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.S.; Darling, C.L.; Featherstone, J.D.; Fried, D. Imaging artificial caries on the occlusal surfaces with polarization-sensitive optical coherence tomography. Caries Res. 2006, 40, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.S.; Fried, D. Remineralization of enamel caries can decrease optical reflectivity. J. Dent. Res. 2006, 85, 804–808. [Google Scholar] [CrossRef]
- Silverstone, L.M. Structure of carious enamel, including the early lesion. Oral Sci. Rev. 1973, 3, 100–160. [Google Scholar]
- Larsen, M.J. Chemical events during tooth dissolution. J. Dent. Res. 1990, 69, 575–634, 575–580; discussion 634–576. [Google Scholar] [CrossRef]
- Inker, L.; Levey, A. Measurement and Estimation of Kidney Function. In Chronic Kidney Disease, Dialysis, and Transplantation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 23–41.e23. [Google Scholar]
- Harvey, N.M.; Carpenter, G.H.; Proctor, G.B.; Klein, J. Normal and frictional interactions of purified human statherin adsorbed on molecularly-smooth solid substrata. Biofouling 2011, 27, 823–835. [Google Scholar] [CrossRef]
- Bibby, B.G. Organic enamel material and caries. Caries Res. 1971, 5, 305–322. [Google Scholar] [CrossRef]
- Robinson, C.; Shore, R.C.; Bonass, W.A.; Brookes, S.J.; Boteva, E.; Kirkham, J. Identification of human serum albumin in human caries lesions of enamel: The role of putative inhibitors of remineralisation. Caries Res. 1998, 32, 193–199. [Google Scholar] [CrossRef]
- Garnett, J.; Dieppe, P. The effects of serum and human albumin on calcium hydroxyapatite crystal growth. Biochem. J. 1990, 266, 863–868. [Google Scholar]
- Robinson, C.; Kirkham, J.; Brookes, S.J.; Shore, R.C. The role of albumin in developing rodent dental enamel: A possible explanation for white spot hypoplasia. J. Dent. Res. 1992, 71, 1270–1274. [Google Scholar] [CrossRef] [PubMed]
- Bjørndal, L.; Thylstrup, A. A structural analysis of approximal enamel caries lesions and subjacent dentin reactions. Eur. J. Oral Sci. 1995, 103, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, A.; Attin, T. Occupational dental erosion from exposure to acids: A review. Occup. Med. 2007, 57, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D. Intrinsic causes of erosion. Monogr. Oral Sci. 2006, 20, 119–139. [Google Scholar] [CrossRef]
- Hannig, M.; Balz, M. Protective properties of salivary pellicles from two different intraoral sites on enamel erosion. Caries Res. 2001, 35, 142–148. [Google Scholar] [CrossRef]
- Ganss, C. Definition of erosion and links to tooth wear. Monogr. Oral Sci. 2006, 20, 9–16. [Google Scholar] [CrossRef]
- Lussi, A.; Schlueter, N.; Rakhmatullina, E.; Ganss, C. Dental erosion—An overview with emphasis on chemical and histopathological aspects. Caries Res. 2011, 45 (Suppl. 1), 2–12. [Google Scholar] [CrossRef]
- Meurman, J.H.; Drysdale, T.; Frank, R.M. Experimental erosion of dentin. Scand. J. Dent. Res. 1991, 99, 457–462. [Google Scholar] [CrossRef]
- Ganss, C.; Klimek, J.; Starck, C. Quantitative analysis of the impact of the organic matrix on the fluoride effect on erosion progression in human dentine using longitudinal microradiography. Arch. Oral Biol. 2004, 49, 931–935. [Google Scholar] [CrossRef]
- Van Nieuw Amerongen, A.; Bolscher, J.G.; Veerman, E.C. Salivary proteins: Protective and diagnostic value in cariology? Caries Res. 2004, 38, 247–253. [Google Scholar] [CrossRef]
- Gao, X.; Jiang, S.; Koh, D.; Hsu, C.Y. Salivary biomarkers for dental caries. Periodontol. 2000 2016, 70, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, W.L.; Helmerhorst, E.J.; Zhang, W.; Salih, E.; Oppenheim, F.G. Acquired enamel pellicle and its potential role in oral diagnostics. Ann. N. Y. Acad. Sci. 2007, 1098, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Hannig, M.; Joiner, A. The structure, function and properties of the acquired pellicle. Monogr. Oral Sci. 2006, 19, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; de Morais Guedes, S.; Ferreira, R.; Lobo, M.J.; Duarte, J.; Ferrer-Correia, A.J.; Tomer, K.B.; Domingues, P.M.; Amado, F.M. Two-dimensional electrophoresis study of in vitro pellicle formation and dental caries susceptibility. Eur. J. Oral Sci. 2006, 114, 147–153. [Google Scholar] [CrossRef]
- Luo, J.; Wang, Y.; Wang, K.; Jiang, W.; Li, X.; Zhang, L. Comparative proteomic analysis on acquired enamel pellicle at two time points in caries-susceptible and caries-free subjects. J. Dent. 2020, 94, 103301. [Google Scholar] [CrossRef]
- Frenkel, E.S.; Ribbeck, K. Salivary mucins protect surfaces from colonization by cariogenic bacteria. Appl. Environ. Microbiol. 2015, 81, 332–338. [Google Scholar] [CrossRef]
- Gabryel-Porowska, H.; Gornowicz, A.; Bielawska, A.; Wójcicka, A.; Maciorkowska, E.; Grabowska, S.Z.; Bielawski, K. Mucin levels in saliva of adolescents with dental caries. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2014, 20, 72–77. [Google Scholar] [CrossRef]
- Angwaravong, O.; Pitiphat, W.; Bolscher, J.G.; Chaiyarit, P. Evaluation of salivary mucins in children with deciduous and mixed dentition: Comparative analysis between high and low caries-risk groups. Clin. Oral Investig. 2015, 19, 1931–1937. [Google Scholar] [CrossRef]
- Szkaradkiewicz-Karpińska, A.K.; Ronij, A.; Goślińska-Kuźniarek, O.; Przybyłek, I.; Szkaradkiewicz, A. MUC7 Level As A New Saliva Risk Factor For Dental Caries In Adult Patients. Int. J. Med. Sci. 2019, 16, 241–246. [Google Scholar] [CrossRef]
- O’Mullane, D.M.; Baez, R.J.; Jones, S.; Lennon, M.A.; Petersen, P.E.; Rugg-Gunn, A.J.; Whelton, H.; Whitford, G.M. Fluoride and Oral Health. Community Dent. Health 2016, 33, 69–99. [Google Scholar]
- Walsh, T.; Worthington, H.V.; Glenny, A.M.; Marinho, V.C.; Jeroncic, A. Fluoride toothpastes of different concentrations for preventing dental caries. Cochrane Database Syst. Rev. 2019, 3, Cd007868. [Google Scholar] [CrossRef] [PubMed]
- Gluzman, R.; Katz, R.V.; Frey, B.J.; McGowan, R. Prevention of root caries: A literature review of primary and secondary preventive agents. Spec. Care Dent. Off. Publ. Am. Assoc. Hosp. Dent. Acad. Dent. Handicap. Am. Soc. Geriatr. Dent. 2013, 33, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Marinho, V.C.; Worthington, H.V.; Walsh, T.; Clarkson, J.E. Fluoride varnishes for preventing dental caries in children and adolescents. Cochrane Database Syst. Rev. 2013, 7, Cd002279. [Google Scholar] [CrossRef]
- Marinho, V.C.; Worthington, H.V.; Walsh, T.; Chong, L.Y. Fluoride gels for preventing dental caries in children and adolescents. Cochrane Database Syst. Rev. 2015, 2015, Cd002280. [Google Scholar] [CrossRef]
- Šket, T.; Kukec, A.; Kosem, R.; Artnik, B. The history of public health use of fluorides in caries prevention. Zdr. Varst. 2017, 56, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Zero, D.T. Dentifrices, mouthwashes, and remineralization/caries arrestment strategies. In BMC Oral Health; BioMed Central: London, UK, 2006; Volume 6, p. S9. [Google Scholar] [CrossRef]
- Weyant, R.J.; Tracy, S.L.; Anselmo, T.T.; Beltrán-Aguilar, E.D.; Donly, K.J.; Frese, W.A.; Hujoel, P.P.; Iafolla, T.; Kohn, W.; Kumar, J.; et al. Topical fluoride for caries prevention: Executive summary of the updated clinical recommendations and supporting systematic review. J. Am. Dent. Assoc. 2013, 144, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Ogaard, B.; Rølla, G.; Helgeland, K. Alkali soluble and alkali insoluble fluoride retention in demineralized enamel in vivo. Scand. J. Dent. Res. 1983, 91, 200–204. [Google Scholar] [CrossRef]
- Rølla, G. On the role of calcium fluoride in the cariostatic mechanism of fluoride. Acta Odontol. Scand. 1988, 46, 341–345. [Google Scholar] [CrossRef]
- Lagerlöf, F.; Ekstrand, J.; Rølla, G. Effect of fluoride addition on ionized calcium in salivary sediment and in saliva. Scand. J. Dent. Res. 1988, 96, 399–404. [Google Scholar] [CrossRef]
- Chow, L.C.; Takagi, S.; Carey, C.M.; Sieck, B.A. Remineralization effects of a two-solution fluoride mouthrinse: An in situ study. J. Dent. Res. 2000, 79, 991–995. [Google Scholar] [CrossRef]
- Chow, L.C.; Takagi, S.; Frukhtbeyn, S.; Sieck, B.A.; Parry, E.E.; Liao, N.S.; Schumacher, G.E.; Markovic, M. Remineralization effect of a low-concentration fluoride rinse in an intraoral model. Caries Res. 2002, 36, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Saxegaard, E.; Rølla, G. Kinetics of acquisition and loss of calcium fluoride by enamel in vivo. Caries Res. 1989, 23, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.J.; Richards, A. The influence of saliva on the formation of calcium fluoride-like material on human dental enamel. Caries Res. 2001, 35, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Marinho, V.C.; Chong, L.Y.; Worthington, H.V.; Walsh, T. Fluoride mouthrinses for preventing dental caries in children and adolescents. Cochrane Database Syst. Rev. 2016, 7, Cd002284. [Google Scholar] [CrossRef] [PubMed]
- Vogel, G.L.; Tenuta, L.M.; Schumacher, G.E.; Chow, L.C. No calcium-fluoride-like deposits detected in plaque shortly after a sodium fluoride mouthrinse. Caries Res. 2010, 44, 108–115. [Google Scholar] [CrossRef]
- Wegehaupt, F.J.; Schneiders, V.; Wiegand, A.; Schmidlin, P.R.; Attin, T. Influence of two different fluoride compounds and an in vitro pellicle on the amount of KOH-soluble fluoride and its retention after toothbrushing. Acta Odontol. Scand. 2009, 67, 355–359. [Google Scholar] [CrossRef]
- Ganss, C.; Schlueter, N.; Klimek, J. Retention of KOH-soluble fluoride on enamel and dentine under erosive conditions--A comparison of in vitro and in situ results. Arch. Oral Biol. 2007, 52, 9–14. [Google Scholar] [CrossRef]
- Wegehaupt, F.J.; Tauböck, T.T.; Sener, B.; Attin, T. Retention of KOH-soluble fluoride formed after application of a SnCl(2)/AmF/NaF containing mouth rinse under erosive conditions. Acta Odontol. Scand. 2012, 70, 272–278. [Google Scholar] [CrossRef]
- Rošin-Grget, K.; Peroš, K.; Sutej, I.; Bašić, K. The cariostatic mechanisms of fluoride. Acta Med. Acad. 2013, 42, 179–188. [Google Scholar] [CrossRef]
- Damé-Teixeira, N.; Deng, D.; Do, T. Streptococcus mutans transcriptome in the presence of sodium fluoride and sucrose. Arch. Oral Biol. 2019, 102, 186–192. [Google Scholar] [CrossRef]
- Koo, H. Strategies to enhance the biological effects of fluoride on dental biofilms. Adv. Dent. Res. 2008, 20, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Jung, J.E.; Choi, H.M.; Jeon, J.G. Effect of brief periodic fluoride treatments on the virulence and composition of a cariogenic biofilm. Biofouling 2018, 34, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Van Loveren, C.; Hoogenkamp, M.A.; Deng, D.M.; ten Cate, J.M. Effects of different kinds of fluorides on enolase and ATPase activity of a fluoride-sensitive and fluoride-resistant Streptococcus mutans strain. Caries Res. 2008, 42, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Van Loveren, C. Antimicrobial activity of fluoride and its in vivo importance: Identification of research questions. Caries Res. 2001, 35 (Suppl. 1), 65–70. [Google Scholar] [CrossRef] [PubMed]
- Kulshrestha, S.; Khan, S.; Hasan, S.; Khan, M.E.; Misba, L.; Khan, A.U. Calcium fluoride nanoparticles induced suppression of Streptococcus mutans biofilm: An in vitro and in vivo approach. Appl. Microbiol. Biotechnol. 2016, 100, 1901–1914. [Google Scholar] [CrossRef]
- Rose, R.K.; Shellis, R.P.; Lee, A.R. The role of cation bridging in microbial fluoride binding. Caries Res. 1996, 30, 458–464. [Google Scholar] [CrossRef]
- Vogel, G.L. Oral fluoride reservoirs and the prevention of dental caries. Monogr. Oral Sci. 2011, 22, 146–157. [Google Scholar] [CrossRef]
- Souza, J.G.S.; Del Bel Cury, A.A.; Cury, J.A.; Tenuta, L.M.A. On the release of fluoride from biofilm reservoirs during a cariogenic challenge: An in situ study. Biofouling 2020, 36, 870–876. [Google Scholar] [CrossRef]
- Stookey, G.K.; Mau, M.S.; Isaacs, R.L.; Gonzalez-Gierbolini, C.; Bartizek, R.D.; Biesbrock, A.R. The relative anticaries effectiveness of three fluoride-containing dentifrices in Puerto Rico. Caries Res. 2004, 38, 542–550. [Google Scholar] [CrossRef]
- Shapira, L.; Shapira, M.; Tandlich, M.; Gedalia, I. Effect of amine fluoride-stannous fluoride containing toothpaste (Meridol) on plaque and gingivitis in adults: A six-month clinical study. J. Int. Acad. Periodontol. 1999, 1, 117–120. [Google Scholar]
- Ganss, C.; Schlueter, N.; Hardt, M.; Schattenberg, P.; Klimek, J. Effect of fluoride compounds on enamel erosion in vitro: A comparison of amine, sodium and stannous fluoride. Caries Res. 2008, 42, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Rugg-Gunn, A.; Bánóczy, J. Fluoride toothpastes and fluoride mouthrinses for home use. Acta Med. Acad. 2013, 42, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Hannig, C.; Gaeding, A.; Basche, S.; Richter, G.; Helbig, R.; Hannig, M. Effect of conventional mouthrinses on initial bioadhesion to enamel and dentin in situ. Caries Res. 2013, 47, 150–161. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, S.; Bartlett, D.W.; Moazzez, R. Efficacy of sodium and stannous fluoride mouthrinses when used before single and multiple erosive challenges. Aust. Dent. J. 2016, 61, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Mengel, R.; Wissing, E.; Schmitz-Habben, A.; Florès-de-Jacoby, L. Comparative study of plaque and gingivitis prevention by AmF/SnF2 and NaF. A clinical and microbiological 9-month study. J. Clin. Periodontol. 1996, 23, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Kensche, A.; Buschbeck, E.; König, B.; Koch, M.; Kirsch, J.; Hannig, C.; Hannig, M. Effect of fluoride mouthrinses and stannous ions on the erosion protective properties of the in situ pellicle. Sci. Rep. 2019, 9, 5336. [Google Scholar] [CrossRef]
- Kirsch, J.; Hannig, M.; Winkel, P.; Basche, S.; Leis, B.; Pütz, N.; Kensche, A.; Hannig, C. Influence of pure fluorides and stannous ions on the initial bacterial colonization in situ. Sci. Rep. 2019, 9, 18499. [Google Scholar] [CrossRef]
- Clark-Perry, D.; Levin, L. Comparison of new formulas of stannous fluoride toothpastes with other commercially available fluoridated toothpastes: A systematic review and meta-analysis of randomised controlled trials. Int. Dent. J. 2020, 70, 418–426. [Google Scholar] [CrossRef]
- Fernández, C.E.; Fontana, M.; Samarian, D.; Cury, J.A.; Rickard, A.H.; González-Cabezas, C. Effect of Fluoride-Containing Toothpastes on Enamel Demineralization and Streptococcus mutans Biofilm Architecture. Caries Res. 2016, 50, 151–158. [Google Scholar] [CrossRef]
- Sharma, N.; He, T.; Barker, M.L.; Biesbrock, A.R. Plaque control evaluation of a stabilized stannous fluoride dentifrice compared to a triclosan dentifrice in a six-week trial. J. Clin. Dent. 2013, 24, 31–36. [Google Scholar]
- Algarni, A.A.; Mussi, M.C.; Moffa, E.B.; Lippert, F.; Zero, D.T.; Siqueira, W.L.; Hara, A.T. The impact of stannous, fluoride ions and its combination on enamel pellicle proteome and dental erosion prevention. PLoS ONE 2015, 10, e0128196. [Google Scholar] [CrossRef]
- Oppermann, R.V.; Johansen, J.R. Effect of fluoride and non-fluoride salts of copper, silver and tin on the acidogenicity of dental plaque in vivo. Scand. J. Dent. Res. 1980, 88, 476–480. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Cox, E.R.; Gwynn, A.V. Effect of a stabilized stannous fluoride dentifrice on plaque acid (toxin) production. J. Clin. Dent. 1995, 6, 84–88. [Google Scholar]
- Forbes, S.; Latimer, J.; Sreenivasan, P.K.; McBain, A.J. Simultaneous Assessment of Acidogenesis-Mitigation and Specific Bacterial Growth-Inhibition by Dentifrices. PLoS ONE 2016, 11, e0149390. [Google Scholar] [CrossRef] [PubMed]
- Tinanoff, N.; Brady, J.M.; Gross, A. The effect of NaF and SnF2 mouthrinses on bacterial colonization of tooth enamel: TEM and SEM studies. Caries Res. 1976, 10, 415–426. [Google Scholar] [CrossRef]
- Rykke, M.; Ellingsen, J.E.; Sønju, T. Chemical analysis and scanning electron microscopy of acquired pellicle formed in vivo on stannous fluoride treated enamel. Scand. J. Dent. Res. 1991, 99, 205–211. [Google Scholar] [CrossRef]
- Schlueter, N.; Klimek, J.; Ganss, C. In vitro efficacy of experimental tin- and fluoride-containing mouth rinses as anti-erosive agents in enamel. J. Dent. 2009, 37, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Ganss, C.; Neutard, L.; von Hinckeldey, J.; Klimek, J.; Schlueter, N. Efficacy of a tin/fluoride rinse: A randomized in situ trial on erosion. J. Dent. Res. 2010, 89, 1214–1218. [Google Scholar] [CrossRef]
- Siqueira, W.L.; Bakkal, M.; Xiao, Y.; Sutton, J.N.; Mendes, F.M. Quantitative proteomic analysis of the effect of fluoride on the acquired enamel pellicle. PLoS ONE 2012, 7, e42204. [Google Scholar] [CrossRef]
- Buzalaf, M.A.; Hannas, A.R.; Kato, M.T. Saliva and dental erosion. J. Appl. Oral Sci. Rev. FOB 2012, 20, 493–502. [Google Scholar] [CrossRef]
- Hannig, C.; Berndt, D.; Hoth-Hannig, W.; Hannig, M. The effect of acidic beverages on the ultrastructure of the acquired pellicle--an in situ study. Arch. Oral Biol. 2009, 54, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Hannig, M.; Fiebiger, M.; Güntzer, M.; Döbert, A.; Zimehl, R.; Nekrashevych, Y. Protective effect of the in situ formed short-term salivary pellicle. Arch. Oral Biol. 2004, 49, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.T.; Ando, M.; González-Cabezas, C.; Cury, J.A.; Serra, M.C.; Zero, D.T. Protective effect of the dental pellicle against erosive challenges in situ. J. Dent. Res. 2006, 85, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Vukosavljevic, D.; Custodio, W.; Buzalaf, M.A.; Hara, A.T.; Siqueira, W.L. Acquired pellicle as a modulator for dental erosion. Arch. Oral Biol. 2014, 59, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Delecrode, T.R.; Siqueira, W.L.; Zaidan, F.C.; Bellini, M.R.; Moffa, E.B.; Mussi, M.C.; Xiao, Y.; Buzalaf, M.A. Identification of acid-resistant proteins in acquired enamel pellicle. J. Dent. 2015, 43, 1470–1475. [Google Scholar] [CrossRef] [PubMed]
- Mutahar, M.; O’Toole, S.; Carpenter, G.; Bartlett, D.; Andiappan, M.; Moazzez, R. Reduced statherin in acquired enamel pellicle on eroded teeth compared to healthy teeth in the same subjects: An in-vivo study. PLoS ONE 2017, 12, e0183660. [Google Scholar] [CrossRef] [PubMed]
- Amaechi, B.T.; Higham, S.M.; Edgar, W.M.; Milosevic, A. Thickness of acquired salivary pellicle as a determinant of the sites of dental erosion. J. Dent. Res. 1999, 78, 1821–1828. [Google Scholar] [CrossRef]
- Carvalho, T.S.; Araújo, T.T.; Ventura, T.M.O.; Dionizio, A.; Câmara, J.V.F.; Moraes, S.M.; Pelá, V.T.; Martini, T.; Leme, J.C.; Derbotolli, A.L.B.; et al. Acquired pellicle protein-based engineering protects against erosive demineralization. J. Dent. 2020, 102, 103478. [Google Scholar] [CrossRef]
- Moussa, D.G.; Siqueira, W.L. Bioinspired caries preventive strategy via customizable pellicles of saliva-derived protein/peptide constructs. Sci. Rep. 2021, 11, 17007. [Google Scholar] [CrossRef]
- Martini, T.; Rios, D.; Cassiano, L.P.S.; Silva, C.M.S.; Taira, E.A.; Ventura, T.M.S.; Pereira, H.; Magalhães, A.C.; Carvalho, T.S.; Baumann, T.; et al. Proteomics of acquired pellicle in gastroesophageal reflux disease patients with or without erosive tooth wear. J. Dent. 2019, 81, 64–69. [Google Scholar] [CrossRef]
- Taira, E.A.; Ventura, T.M.S.; Cassiano, L.P.S.; Silva, C.M.S.; Martini, T.; Leite, A.L.; Rios, D.; Magalhães, A.C.; Buzalaf, M.A.R. Changes in the Proteomic Profile of Acquired Enamel Pellicles as a Function of Their Time of Formation and Hydrochloric Acid Exposure. Caries Res. 2018, 52, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Hannig, C.; Hannig, M.; Attin, T. Enzymes in the acquired enamel pellicle. Eur. J. Oral Sci. 2005, 113, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Hannig, C.; Attin, T.; Hannig, M.; Henze, E.; Brinkmann, K.; Zech, R. Immobilisation and activity of human alpha-amylase in the acquired enamel pellicle. Arch. Oral Biol. 2004, 49, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Hannig, C.; Spitzmüller, B.; Miller, M.; Hellwig, E.; Hannig, M. Intrinsic enzymatic crosslinking and maturation of the in situ pellicle. Arch. Oral Biol. 2008, 53, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, J.; Jung, A.; Hille, K.; König, B.; Hannig, C.; Kölling-Speer, I.; Speer, K.; Hannig, M. Effect of fragaria vesca, hamamelis and tormentil on the initial bacterial colonization in situ. Arch. Oral Biol. 2020, 118, 104853. [Google Scholar] [CrossRef]
- Bennick, A.; Cannon, M.; Madapallimattam, G. The nature of the hydroxyapatite-binding site in salivary acidic proline-rich proteins. Biochem. J. 1979, 183, 115–126. [Google Scholar] [CrossRef]
- Hay, D.I. The interaction of human parotid salivary proteins with hydroxyapatite. Arch. Oral Biol. 1973, 18, 1517–1529. [Google Scholar] [CrossRef]
- Moreno, E.C.; Kresak, M.; Hay, D.I. Adsorption of two human parotid salivary macromolecules on hydroxy-, fluorhydroxy- and fluorapatites. Arch. Oral Biol. 1978, 23, 525–533. [Google Scholar] [CrossRef]
- Vassilakos, N.; Arnebrant, T.; Glantz, P.O. Adsorption of whole saliva onto hydrophilic and hydrophobic solid surfaces: Influence of concentration, ionic strength and pH. Scand. J. Dent. Res. 1992, 100, 346–353. [Google Scholar] [CrossRef]
- Vassilakos, N.; Arnebrant, T.; Rundegren, J.; Glantz, P.O. In vitro interactions of anionic and cationic surfactants with salivary fractions on well-defined solid surfaces. Acta Odontol. Scand. 1992, 50, 179–188. [Google Scholar] [CrossRef]
- Vassilakos, N.; Arnebrant, T.; Glantz, P.O. An in vitro study of salivary film formation at solid/liquid interfaces. Scand. J. Dent. Res. 1993, 101, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.L.; Lamkin, M.S.; Oppenheim, F.G. Adsorption of human salivary proteins to hydroxyapatite: A comparison between whole saliva and glandular salivary secretions. J. Dent. Res. 1992, 71, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Lamkin, M.S.; Arancillo, A.A.; Oppenheim, F.G. Temporal and compositional characteristics of salivary protein adsorption to hydroxyapatite. J. Dent. Res. 1996, 75, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Lamkin, M.S.; Oppenheim, F.G. Pellicle precursor proteins: Acidic proline-rich proteins, statherin, and histatins, and their crosslinking reaction by oral transglutaminase. J. Dent. Res. 1999, 78, 1696–1703. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Berg, E.A.; Costello, C.E.; Troxler, R.F.; Oppenheim, F.G. Identification of protein components in human acquired enamel pellicle and whole saliva using novel proteomics approaches. J. Biol. Chem. 2003, 278, 5300–5308. [Google Scholar] [CrossRef]
- Yao, Y.; Grogan, J.; Zehnder, M.; Lendenmann, U.; Nam, B.; Wu, Z.; Costello, C.E.; Oppenheim, F.G. Compositional analysis of human acquired enamel pellicle by mass spectrometry. Arch. Oral Biol. 2001, 46, 293–303. [Google Scholar] [CrossRef]
- Oppenheim, F.G.; Helmerhorst, E.J.; Lendenmann, U.; Offner, G.D. Anti-candidal activity of genetically engineered histatin variants with multiple functional domains. PLoS ONE 2012, 7, e51479. [Google Scholar] [CrossRef]
- Marin, L.M.; Xiao, Y.; Cury, J.A.; Siqueira, W.L. Engineered Salivary Peptides Reduce Enamel Demineralization Provoked by Cariogenic, S. mutans Biofilm. Microorganisms 2022, 10, 742. [Google Scholar] [CrossRef]
- Siqueira, W.L.; Custodio, W.; McDonald, E.E. New insights into the composition and functions of the acquired enamel pellicle. J. Dent. Res. 2012, 91, 1110–1118. [Google Scholar] [CrossRef]
- Araújo, T.T.; Carvalho, T.S.; Dionizio, A.; Debortolli, A.L.B.; Ventura, T.M.O.; Souza, B.M.; Feitosa, C.; Barbosa, H.A.P.; Ribeiro, C.; Martini, T.; et al. Protein-based engineering of the initial acquired enamel pellicle in vivo: Proteomic evaluation. J. Dent. 2022, 116, 103874. [Google Scholar] [CrossRef]
- Basiri, T.; Johnson, N.D.; Moffa, E.B.; Mulyar, Y.; Serra Nunes, P.L.; Machado, M.; Siqueira, W.L. Duplicated or Hybridized Peptide Functional Domains Promote Oral Homeostasis. J. Dent. Res. 2017, 96, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Valente, M.T.; Moffa, E.B.; Crosara, K.T.B.; Xiao, Y.; de Oliveira, T.M.; Machado, M.; Siqueira, W.L. Acquired Enamel Pellicle Engineered Peptides: Effects on Hydroxyapatite Crystal Growth. Sci. Rep. 2018, 8, 3766. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Karttunen, M.; Jalkanen, J.; Mussi, M.C.; Liao, Y.; Grohe, B.; Lagugné-Labarthet, F.; Siqueira, W.L. Hydroxyapatite Growth Inhibition Effect of Pellicle Statherin Peptides. J. Dent. Res. 2015, 94, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Kosoric, J.; Hector, M.P.; Anderson, P. An in vitro scanning microradiography study of the reduction in hydroxyapatite demineralization rate by statherin-like peptides as a function of increasing N-terminal length. Eur. J. Oral Sci. 2011, 119 (Suppl. 1), 13–18. [Google Scholar] [CrossRef]
- Kosoric, J.; Williams, R.A.D.; Hector, M.P.; Anderson, P. A Synthetic Peptide Based on a Natural Salivary Protein Reduces Demineralisation in Model Systems for Dental Caries and Erosion. Int. J. Pept. Res. Ther. 2007, 13, 497–503. [Google Scholar] [CrossRef]
- Tang, R.; Hass, M.; Wu, W.; Gulde, S.; Nancollas, G.H. Constant composition dissolution of mixed phases. II. Selective dissolution of calcium phosphates. J. Colloid Interface Sci. 2003, 260, 379–384. [Google Scholar] [CrossRef]
- Zahradnik, R.T.; Moreno, E.C.; Burke, E.J. Effect of salivary pellicle on enamel subsurface demineralization in vitro. J. Dent. Res. 1976, 55, 664–670. [Google Scholar] [CrossRef]
- Goobes, G.; Goobes, R.; Shaw, W.J.; Gibson, J.M.; Long, J.R.; Raghunathan, V.; Schueler-Furman, O.; Popham, J.M.; Baker, D.; Campbell, C.T.; et al. The structure, dynamics, and energetics of protein adsorption-lessons learned from adsorption of statherin to hydroxyapatite. Magn. Reason. Chem. 2007, 45 (Suppl. 1), S32–S47. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, B.; Li, M.; Wang, Y.; Yang, X.; Li, J. Salivary acquired pellicle-inspired DpSpSEEKC peptide for the restoration of demineralized tooth enamel. Biomed. Mater. 2017, 12, 025007. [Google Scholar] [CrossRef]
- Flemming, J.; Meyer-Probst, C.T.; Speer, K.; Kölling-Speer, I.; Hannig, C.; Hannig, M. Preventive Applications of Polyphenols in Dentistry—A Review. Int. J. Mol. Sci. 2021, 22, 4892. [Google Scholar] [CrossRef]
- Schestakow, A.; Nekrashevych, Y.; Hoth-Hannig, W.; Hannig, M. Influence of periodic polyphenol treatment on the anti-erosive potential of the acquired enamel pellicle-A qualitative exploratory study. J. Dent. 2022, 124, 104236. [Google Scholar] [CrossRef] [PubMed]
- Schestakow, A.; Pütz, N.; Guth, M.S.; Eisenmenger, T.A.; Dudek, J.; Hannig, M. Influence of a hydroxyapatite suspension on 48-h dental biofilm formation in-situ. Arch. Oral Biol. 2022, 136, 105388. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flemming, J.; Hannig, C.; Hannig, M. Caries Management—The Role of Surface Interactions in De- and Remineralization-Processes. J. Clin. Med. 2022, 11, 7044. https://doi.org/10.3390/jcm11237044
Flemming J, Hannig C, Hannig M. Caries Management—The Role of Surface Interactions in De- and Remineralization-Processes. Journal of Clinical Medicine. 2022; 11(23):7044. https://doi.org/10.3390/jcm11237044
Chicago/Turabian StyleFlemming, Jasmin, Christian Hannig, and Matthias Hannig. 2022. "Caries Management—The Role of Surface Interactions in De- and Remineralization-Processes" Journal of Clinical Medicine 11, no. 23: 7044. https://doi.org/10.3390/jcm11237044
APA StyleFlemming, J., Hannig, C., & Hannig, M. (2022). Caries Management—The Role of Surface Interactions in De- and Remineralization-Processes. Journal of Clinical Medicine, 11(23), 7044. https://doi.org/10.3390/jcm11237044