

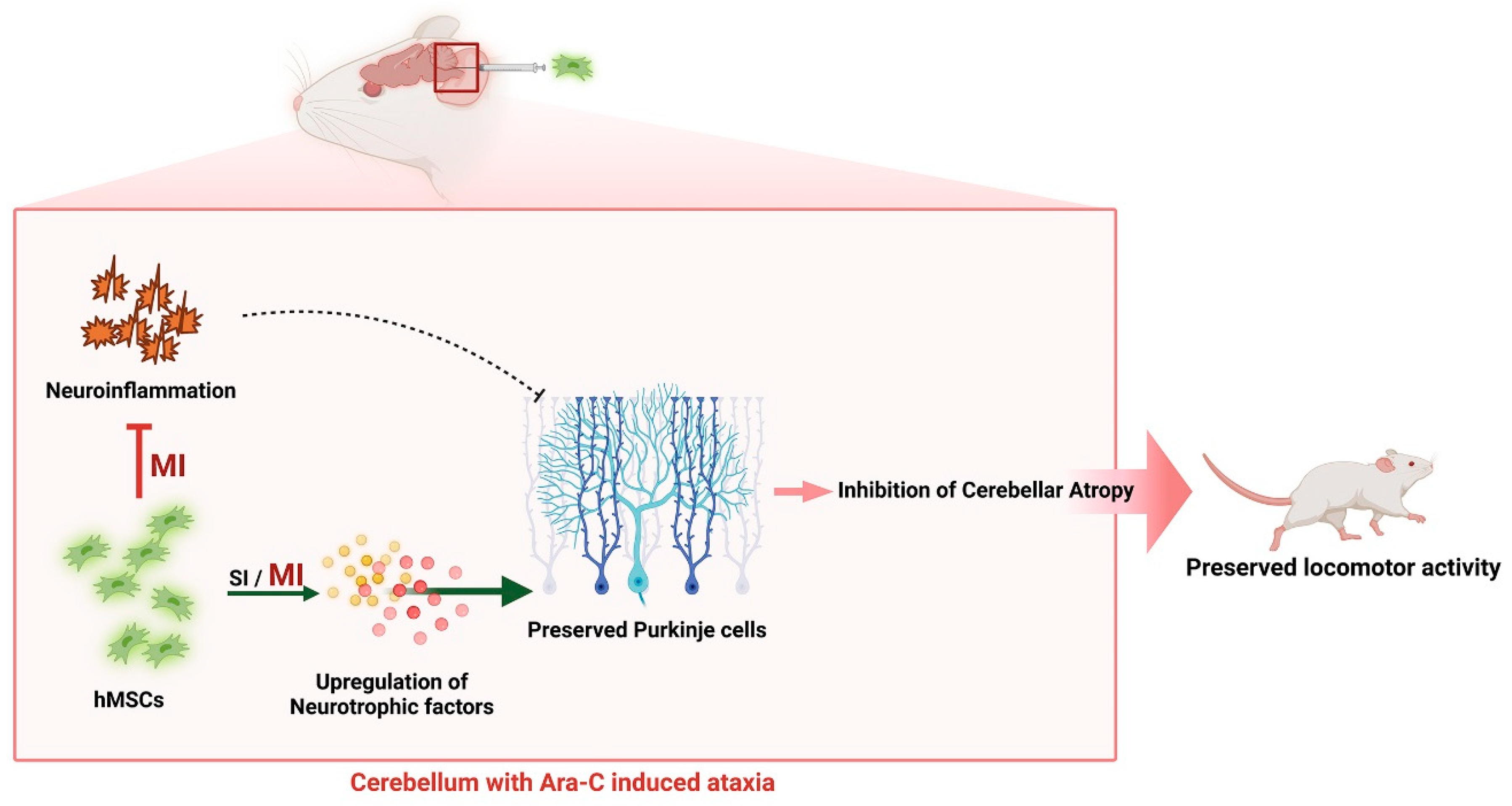
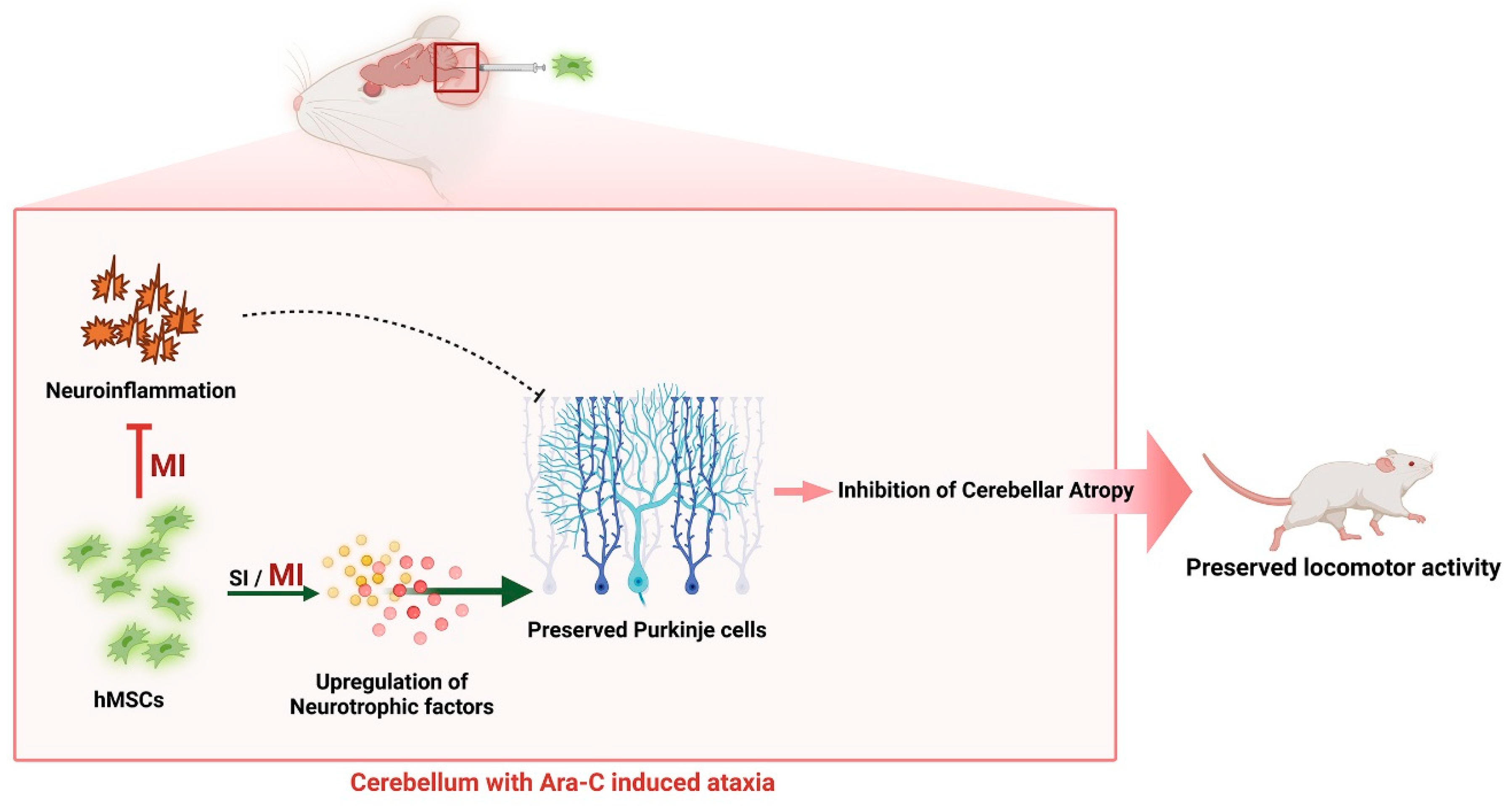
Mesenchymal Stem Cell Transplantation Ameliorates Ara-C-Induced Motor Deficits in a Mouse Model of Cerebellar Ataxia

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Disease Model
2.2. hMSC Implantation
2.3. Behavioral Tests
2.4. Assessment of Cerebellar Weight
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
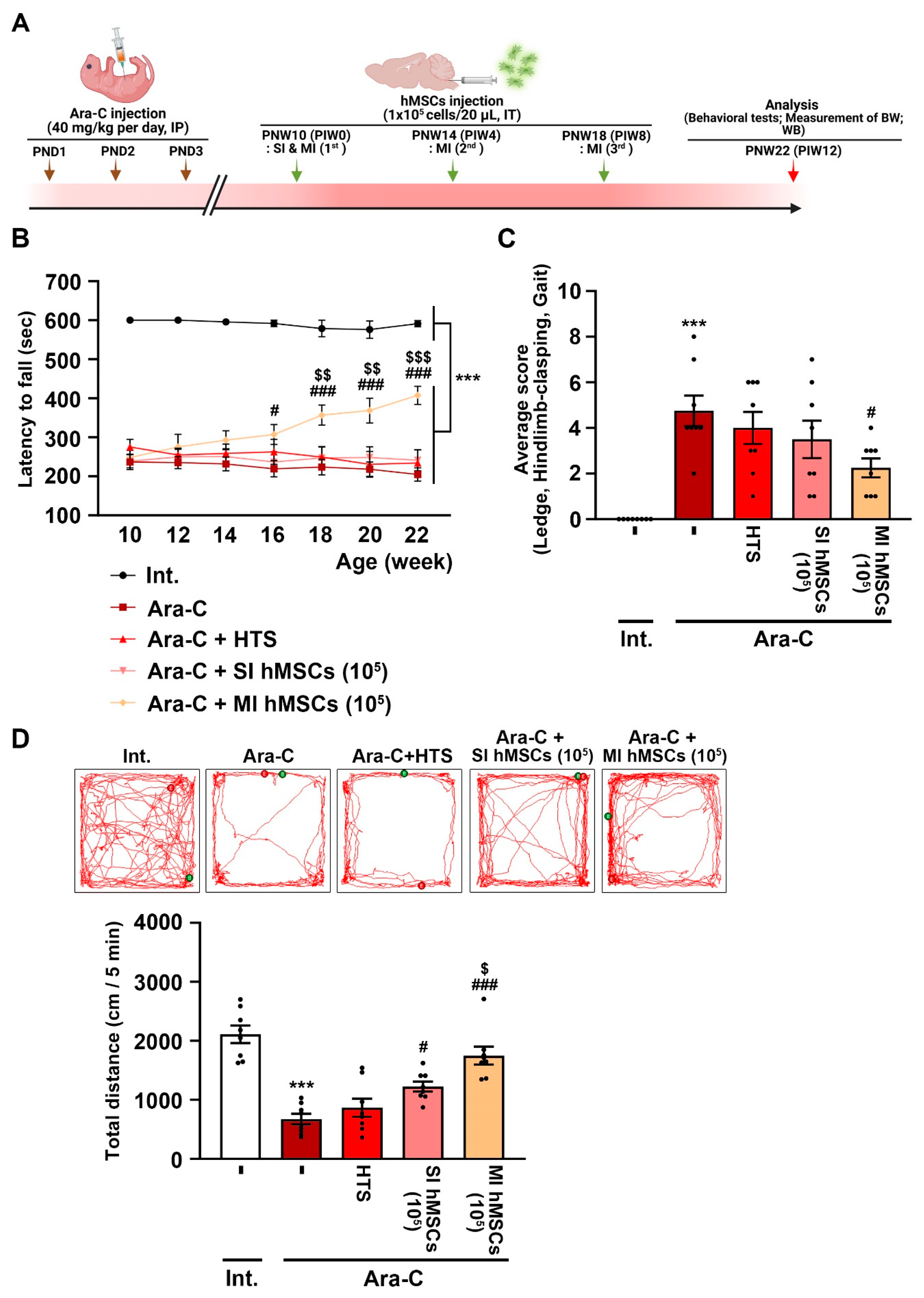
3.1. Effects of hMSC Treatment on Impaired Behaviors of Ara-C-Induced CA Mice
3.2. Effects of hMSC Treatment on Cell Damage in the Cerebellum of Ara-C-Induced CA Mice
3.3. Increased Levels of Neurotrophic Factors and Anti-Inflammatory Effects via hMSC Treatment in the Cerebellum of CA Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klockgether, T. Sporadic ataxia with adult onset: Classification and diagnostic criteria. Lancet Neurol. 2010, 9, 94–104. [Google Scholar] [CrossRef]
- Manto, M.; Marmolino, D. Animal models of human cerebellar ataxias: A cornerstone for the therapies of the twenty-first century. Cerebellum 2009, 8, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Sarva, H.; Shanker, V.L. Treatment Options in Degenerative Cerebellar Ataxia: A Systematic Review. Mov. Disord. Clin. Pract. 2014, 1, 291–298. [Google Scholar] [CrossRef]
- Kwei, K.T.; Kuo, S.H. An Overview of the Current State and the Future of Ataxia Treatments. Neurol. Clin. 2020, 38, 449–467. [Google Scholar] [CrossRef]
- Sarugaser, R.; Hanoun, L.; Keating, A.; Stanford, W.L.; Davies, J.E. Human mesenchymal stem cells self-renew and differentiate according to a deterministic hierarchy. PLoS ONE 2009, 4, e6498. [Google Scholar] [CrossRef] [Green Version]
- Turgeman, G. The therapeutic potential of mesenchymal stem cells in Alzheimer’s disease: Converging mechanisms. Neural. Regen. Res. 2015, 10, 698–699. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Mesenchymal stem cell therapy in Parkinson’s disease animal models. Curr. Res. Transl. Med. 2017, 65, 51–60. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, C.; Teng, J.J.; Zhao, R.L.; Song, Y.Q.; Zhang, C. Multiple administrations of human marrow stromal cells through cerebrospinal fluid prolong survival in a transgenic mouse model of amyotrophic lateral sclerosis. Cytotherapy 2009, 11, 299–306. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef]
- Joshi, M.; Patil, P.B.; He, Z.; Holgersson, J.; Olausson, M.; Sumitran-Holgersson, S. Fetal liver-derived mesenchymal stromal cells augment engraftment of transplanted hepatocytes. Cytotherapy 2012, 14, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.L.; Zhao, Y.L.; Hao, H.J.; Fu, X.B.; Han, W.D. MSCs: Biological characteristics, clinical applications and their outstanding concerns. Ageing Res. Rev. 2011, 10, 93–103. [Google Scholar] [CrossRef]
- In ’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; de Groot-Swings, G.M.; Claas, F.H.; Fibbe, W.E.; Kanhai, H.H. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta. Stem Cells 2004, 22, 1338–1345. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In search of the in vivo identity of mesenchymal stem cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [Green Version]
- Staff, N.P.; Jones, D.T.; Singer, W. Mesenchymal Stromal Cell Therapies for Neurodegenerative Diseases. Mayo Clin. Proc. 2019, 94, 892–905. [Google Scholar] [CrossRef] [Green Version]
- Paul, G.; Anisimov, S.V. The secretome of mesenchymal stem cells: Potential implications for neuroregeneration. Biochimie 2013, 95, 2246–2256. [Google Scholar] [CrossRef]
- Correia, J.S.; Neves-Carvalho, A.; Mendes-Pinheiro, B.; Pires, J.; Teixeira, F.G.; Lima, R.; Monteiro, S.; Silva, N.A.; Soares-Cunha, C.; Serra, S.C.; et al. Preclinical Assessment of Mesenchymal-Stem-Cell-Based Therapies in Spinocerebellar Ataxia Type 3. Biomedicines 2021, 9, 1754. [Google Scholar] [CrossRef]
- Li, T.; Liu, Y.; Yu, L.; Lao, J.; Zhang, M.; Jin, J.; Lu, Z.; Liu, Z.; Xu, Y. Human Umbilical Cord Mesenchymal Stem Cells Protect Against SCA3 by Modulating the Level of 70 kD Heat Shock Protein. Cell Mol. Neurobiol. 2018, 38, 641–655. [Google Scholar] [CrossRef]
- Chang, Y.K.; Chen, M.H.; Chiang, Y.H.; Chen, Y.F.; Ma, W.H.; Tseng, C.Y.; Soong, B.W.; Ho, J.H.; Lee, O.K. Mesenchymal stem cell transplantation ameliorates motor function deterioration of spinocerebellar ataxia by rescuing cerebellar Purkinje cells. J. Biomed. Sci. 2011, 18, 54. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Yoon, D.; Nam, Y.; Seo, D.; Kim, J.H.; Kim, M.S.; Lee, T.Y.; Kim, K.S.; Ko, P.W.; Lee, H.W.; et al. Lipopolysaccharide administration for a mouse model of cerebellar ataxia with neuroinflammation. Sci. Rep. 2020, 10, 13337. [Google Scholar] [CrossRef]
- Nam, Y.; Yoon, D.; Hong, J.; Kim, M.S.; Lee, T.Y.; Kim, K.S.; Lee, H.W.; Suk, K.; Kim, S.R. Therapeutic Effects of Human Mesenchymal Stem Cells in a Mouse Model of Cerebellar Ataxia with Neuroinflammation. J. Clin. Med. 2020, 9, 3654. [Google Scholar] [CrossRef]
- Carballo-Pedrares, N.; Sanjurjo-Rodriguez, C.; Senaris, J.; Diaz-Prado, S.; Rey-Rico, A. Chondrogenic Differentiation of Human Mesenchymal Stem Cells via SOX9 Delivery in Cationic Niosomes. Pharmaceutics 2022, 14, 2327. [Google Scholar] [CrossRef]
- Tsai, Y.A.; Liu, R.S.; Lirng, J.F.; Yang, B.H.; Chang, C.H.; Wang, Y.C.; Wu, Y.S.; Ho, J.H.; Lee, O.K.; Soong, B.W. Treatment of Spinocerebellar Ataxia With Mesenchymal Stem Cells: A Phase I/IIa Clinical Study. Cell Transplant. 2017, 26, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, S.; Shuvaev, A.N.; Iizuka, A.; Nakamura, K.; Hirai, H. Mesenchymal stem cells ameliorate cerebellar pathology in a mouse model of spinocerebellar ataxia type 1. Cerebellum 2014, 13, 323–330. [Google Scholar] [CrossRef]
- Dworkin, L.A.; Goldman, R.D.; Zivin, L.S.; Fuchs, P.C. Cerebellar toxicity following high-dose cytosine arabinoside. J. Clin. Oncol. 1985, 3, 613–616. [Google Scholar] [CrossRef]
- Salinsky, M.C.; Levine, R.L.; Aubuchon, J.P.; Schutta, H.S. Acute cerebellar dysfunction with high-dose ARA-C therapy. Cancer 1983, 51, 426–429. [Google Scholar] [CrossRef]
- Tran, P.N.; Kong, X.T. Cytarabine Induced Acute Cerebellar Syndrome during Hyper-CVAD Treatment for B-Cell Acute Lymphoblastic Leukemia. Case Rep. Neurol. 2017, 9, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.J.; Sun, J.J.; Qian, L.; Liu, Z.; Zhang, Z.; Cao, W.; Li, W.; Xu, Y. Human umbilical mesenchymal stem cells enhance the expression of neurotrophic factors and protect ataxic mice. Brain Res. 2011, 1402, 122–131. [Google Scholar] [CrossRef]
- Guyenet, S.J.; Furrer, S.A.; Damian, V.M.; Baughan, T.D.; La Spada, A.R.; Garden, G.A. A simple composite phenotype scoring system for evaluating mouse models of cerebellar ataxia. J. Vis. Exp. 2010, 39, e1787. [Google Scholar] [CrossRef] [Green Version]
- Yerger, J.; Cougnoux, A.C.; Abbott, C.B.; Luke, R.; Clark, T.S.; Cawley, N.X.; Porter, F.D.; Davidson, C.D. Phenotype assessment for neurodegenerative murine models with ataxia and application to Niemann-Pick disease, type C1. Biol. Open 2022, 11, bio059052. [Google Scholar] [CrossRef]
- Leem, E.; Kim, H.J.; Choi, M.; Kim, S.; Oh, Y.S.; Lee, K.J.; Choe, Y.S.; Um, J.Y.; Shin, W.H.; Jeong, J.Y.; et al. Upregulation of neuronal astrocyte elevated gene-1 protects nigral dopaminergic neurons in vivo. Cell Death Dis. 2018, 9, 449. [Google Scholar] [CrossRef]
- Airey, D.C.; Lu, L.; Williams, R.W. Genetic control of the mouse cerebellum: Identification of quantitative trait loci modulating size and architecture. J. Neurosci. 2001, 21, 5099–5109. [Google Scholar] [CrossRef] [Green Version]
- Scheper, V.; Schwieger, J.; Hamm, A.; Lenarz, T.; Hoffmann, A. BDNF-overexpressing human mesenchymal stem cells mediate increased neuronal protection in vitro. J. Neurosci. Res. 2019, 97, 1414–1429. [Google Scholar] [CrossRef] [Green Version]
- Ashizawa, T.; Xia, G. Ataxia. Contin. (Minneap Minn) 2016, 22, 1208–1226. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, H.; Manto, M.; Gandini, J. Recent Advances in the Treatment of Cerebellar Disorders. Brain Sci. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Sarukhan, A.; Zanotti, L.; Viola, A. Mesenchymal stem cells: Myths and reality. Swiss Med. Wkly. 2015, 145, w14229. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.D.V.; Pickard, M.R.; Johnson, W.E.B. The Comparative Effects of Mesenchymal Stem Cell Transplantation Therapy for Spinal Cord Injury in Humans and Animal Models: A Systematic Review and Meta-Analysis. Biology 2021, 10, 230. [Google Scholar] [CrossRef]
- Hare, J.M.; Traverse, J.H.; Henry, T.D.; Dib, N.; Strumpf, R.K.; Schulman, S.P.; Gerstenblith, G.; DeMaria, A.N.; Denktas, A.E.; Gammon, R.S.; et al. A randomized, double-blind, placebo-controlled, dose-escalation study of intravenous adult human mesenchymal stem cells (prochymal) after acute myocardial infarction. J. Am. Coll. Cardiol. 2009, 54, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Wu, W.; Xu, X.; Liao, L.; Zheng, F.; Messinger, S.; Sun, X.; Chen, J.; Yang, S.; Cai, J.; et al. Induction therapy with autologous mesenchymal stem cells in living-related kidney transplants: A randomized controlled trial. JAMA 2012, 307, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Perico, N.; Casiraghi, F.; Introna, M.; Gotti, E.; Todeschini, M.; Cavinato, R.A.; Capelli, C.; Rambaldi, A.; Cassis, P.; Rizzo, P.; et al. Autologous mesenchymal stromal cells and kidney transplantation: A pilot study of safety and clinical feasibility. Clin. J. Am. Soc. Nephrol. 2011, 6, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Katakowski, M.; Li, Y.; Lu, D.; Wang, L.; Zhang, L.; Chen, J.; Xu, Y.; Gautam, S.; Mahmood, A.; et al. Human bone marrow stromal cell cultures conditioned by traumatic brain tissue extracts: Growth factor production. J. Neurosci. Res. 2002, 69, 687–691. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chen, J.; Cui, Y.; Lu, M.; Elias, S.B.; Mitchell, J.B.; Hammill, L.; Vanguri, P.; Chopp, M. Human bone marrow stromal cell treatment improves neurological functional recovery in EAE mice. Exp. Neurol. 2005, 195, 16–26. [Google Scholar] [CrossRef]
- Volkman, R.; Offen, D. Concise Review: Mesenchymal Stem Cells in Neurodegenerative Diseases. Stem Cells 2017, 35, 1867–1880. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, M.A.; Panet, H.; Barhum, Y.; Melamed, E.; Offen, D. Increased survival and migration of engrafted mesenchymal bone marrow stem cells in 6-hydroxydopamine-lesioned rodents. Neurosci. Lett. 2006, 395, 124–128. [Google Scholar] [CrossRef]
- Nakamura, K.; Mieda, T.; Suto, N.; Matsuura, S.; Hirai, H. Mesenchymal stem cells as a potential therapeutic tool for spinocerebellar ataxia. Cerebellum 2015, 14, 165–170. [Google Scholar] [CrossRef]
- Chopp, M.; Li, Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002, 1, 92–100. [Google Scholar] [CrossRef]
- Oliveira Miranda, C.; Marcelo, A.; Silva, T.P.; Barata, J.; Vasconcelos-Ferreira, A.; Pereira, D.; Nobrega, C.; Duarte, S.; Barros, I.; Alves, J.; et al. Repeated Mesenchymal Stromal Cell Treatment Sustainably Alleviates Machado-Joseph Disease. Mol. Ther. 2018, 26, 2131–2151. [Google Scholar] [CrossRef] [Green Version]
- Tatsuoka, Y.; Kato, Y.; Imura, H. Effect of DN-1417, a synthetic thyrotropin-releasing hormone analogue, on [3H]GABA binding in the cerebellum of ataxic rats. Neurosci. Lett. 1985, 53, 63–68. [Google Scholar] [CrossRef]
- Lopez, J.A.; Agarwal, R.P. Acute cerebellar toxicity after high-dose cytarabine associated with CNS accumulation of its metabolite, uracil arabinoside. Cancer Treat. Rep. 1984, 68, 1309–1310. [Google Scholar]
- Mitoma, H.; Manto, M.; Shaikh, A.G. Mechanisms of Ethanol-Induced Cerebellar Ataxia: Underpinnings of Neuronal Death in the Cerebellum. Int. J. Environ. Res. Public Health 2021, 18, 8678. [Google Scholar] [CrossRef]
- Cendelin, J. From mice to men: Lessons from mutant ataxic mice. Cerebellum Ataxias 2014, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Hoxha, E.; Balbo, I.; Miniaci, M.C.; Tempia, F. Purkinje Cell Signaling Deficits in Animal Models of Ataxia. Front. Synaptic Neurosci. 2018, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Xia, G.; McFarland, K.N.; Wang, K.; Sarkar, P.S.; Yachnis, A.T.; Ashizawa, T. Purkinje cell loss is the major brain pathology of spinocerebellar ataxia type 10. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1409–1411. [Google Scholar] [CrossRef] [Green Version]
- Manto, M. Toxic agents causing cerebellar ataxias. Handb. Clin. Neurol. 2012, 103, 201–213. [Google Scholar] [CrossRef]
- Evert, B.O.; Vogt, I.R.; Kindermann, C.; Ozimek, L.; de Vos, R.A.; Brunt, E.R.; Schmitt, I.; Klockgether, T.; Wullner, U. Inflammatory genes are upregulated in expanded ataxin-3-expressing cell lines and spinocerebellar ataxia type 3 brains. J. Neurosci. 2001, 21, 5389–5396. [Google Scholar] [CrossRef] [Green Version]
- Cvetanovic, M.; Ingram, M.; Orr, H.; Opal, P. Early activation of microglia and astrocytes in mouse models of spinocerebellar ataxia type 1. Neuroscience 2015, 289, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Zemel’ko, V.I.; Kozhukharova, I.V.; Kovaleva, Z.V.; Domnina, A.P.; Pugovkina, N.A.; Fridlianskaia, I.I.; Puzanov, M.V.; Anisimov, S.V.; Grinchuk, T.M.; Nikol’skii, N.N. [BDNF secretion in human mesenchymal stem cells isolated from bone marrow, endometrium and adipose tissue]. Tsitologiia 2014, 56, 204–211. [Google Scholar]
- Sun, S.; Zhang, Q.; Li, M.; Gao, P.; Huang, K.; Beejadhursing, R.; Jiang, W.; Lei, T.; Zhu, M.; Shu, K. GDNF Promotes Survival and Therapeutic Efficacy of Human Adipose-Derived Mesenchymal Stem Cells in a Mouse Model of Parkinson’s Disease. Cell Transplant. 2020, 29, 963689720908512. [Google Scholar] [CrossRef] [Green Version]
- Mount, H.T.; Dean, D.O.; Alberch, J.; Dreyfus, C.F.; Black, I.B. Glial cell line-derived neurotrophic factor promotes the survival and morphologic differentiation of Purkinje cells. Proc. Natl. Acad. Sci. USA 1995, 92, 9092–9096. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, P.M.; Borghesani, P.R.; Levy, R.L.; Pomeroy, S.L.; Segal, R.A. Abnormal cerebellar development and foliation in BDNF-/- mice reveals a role for neurotrophins in CNS patterning. Neuron 1997, 19, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.E.; Mason, C.A. Granule neuron regulation of Purkinje cell development: Striking a balance between neurotrophin and glutamate signaling. J. Neurosci. 1998, 18, 3563–3573. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.R.; Chen, C.; Schwartz, P.M.; Segal, R.A. Brain-derived neurotrophic factor modulates cerebellar plasticity and synaptic ultrastructure. J. Neurosci. 2002, 22, 1316–1327. [Google Scholar] [CrossRef]
- Tolbert, D.L.; Clark, B.R. GDNF and IGF-I trophic factors delay hereditary Purkinje cell degeneration and the progression of gait ataxia. Exp. Neurol. 2003, 183, 205–219. [Google Scholar] [CrossRef]
- Martins, L.F.; Costa, R.O.; Pedro, J.R.; Aguiar, P.; Serra, S.C.; Teixeira, F.G.; Sousa, N.; Salgado, A.J.; Almeida, R.D. Mesenchymal stem cells secretome-induced axonal outgrowth is mediated by BDNF. Sci. Rep. 2017, 7, 4153. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, Y.; Nakamura, S.; Matsuo, A.; Akiguchi, I. Glial cell line-derived neurotrophic factor-like immunoreactivity in the cerebella of normal subjects and patients with multiple system atrophy. Acta Neuropathol. 2000, 100, 2300. [Google Scholar] [CrossRef]
- Aikawa, T.; Mogushi, K.; Iijima-Tsutsui, K.; Ishikawa, K.; Sakurai, M.; Tanaka, H.; Mizusawa, H.; Watase, K. Loss of MyD88 alters neuroinflammatory response and attenuates early Purkinje cell loss in a spinocerebellar ataxia type 6 mouse model. Hum. Mol. Genet. 2015, 24, 4780–4791. [Google Scholar] [CrossRef] [Green Version]
- Muzes, G.; Sipos, F. Mesenchymal Stem Cell-Derived Secretome: A Potential Therapeutic Option for Autoimmune and Immune-Mediated Inflammatory Diseases. Cells 2022, 11, 2300. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, N.; Sharma, C.; Jung, U.J.; Kim, S.; Nam, Y.; Kim, K.-S.; Suk, K.; Lee, H.-W.; Kim, S.R. Mesenchymal Stem Cell Transplantation Ameliorates Ara-C-Induced Motor Deficits in a Mouse Model of Cerebellar Ataxia. J. Clin. Med. 2023, 12, 1756. https://doi.org/10.3390/jcm12051756
Park N, Sharma C, Jung UJ, Kim S, Nam Y, Kim K-S, Suk K, Lee H-W, Kim SR. Mesenchymal Stem Cell Transplantation Ameliorates Ara-C-Induced Motor Deficits in a Mouse Model of Cerebellar Ataxia. Journal of Clinical Medicine. 2023; 12(5):1756. https://doi.org/10.3390/jcm12051756
Chicago/Turabian StylePark, Narae, Chanchal Sharma, Un Ju Jung, Sehwan Kim, Youngpyo Nam, Kyung-Suk Kim, Kyoungho Suk, Ho-Won Lee, and Sang Ryong Kim. 2023. "Mesenchymal Stem Cell Transplantation Ameliorates Ara-C-Induced Motor Deficits in a Mouse Model of Cerebellar Ataxia" Journal of Clinical Medicine 12, no. 5: 1756. https://doi.org/10.3390/jcm12051756