
Effects of Hypoxia and Inflammation on Hepcidin Concentration in Non-Anaemic COVID-19 Patients

 , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Study Design
2.3. Blood Sampling
2.4. Methods
2.5. Statistical Analysis
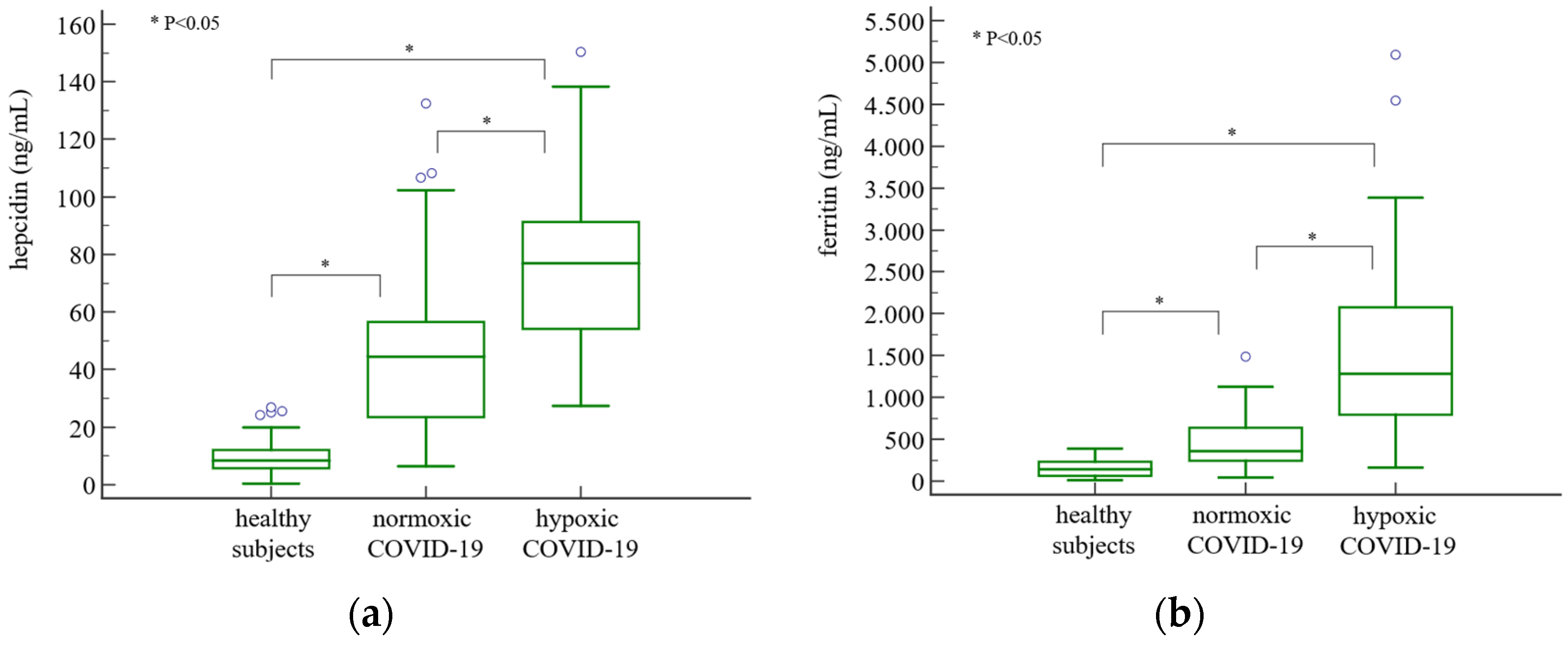
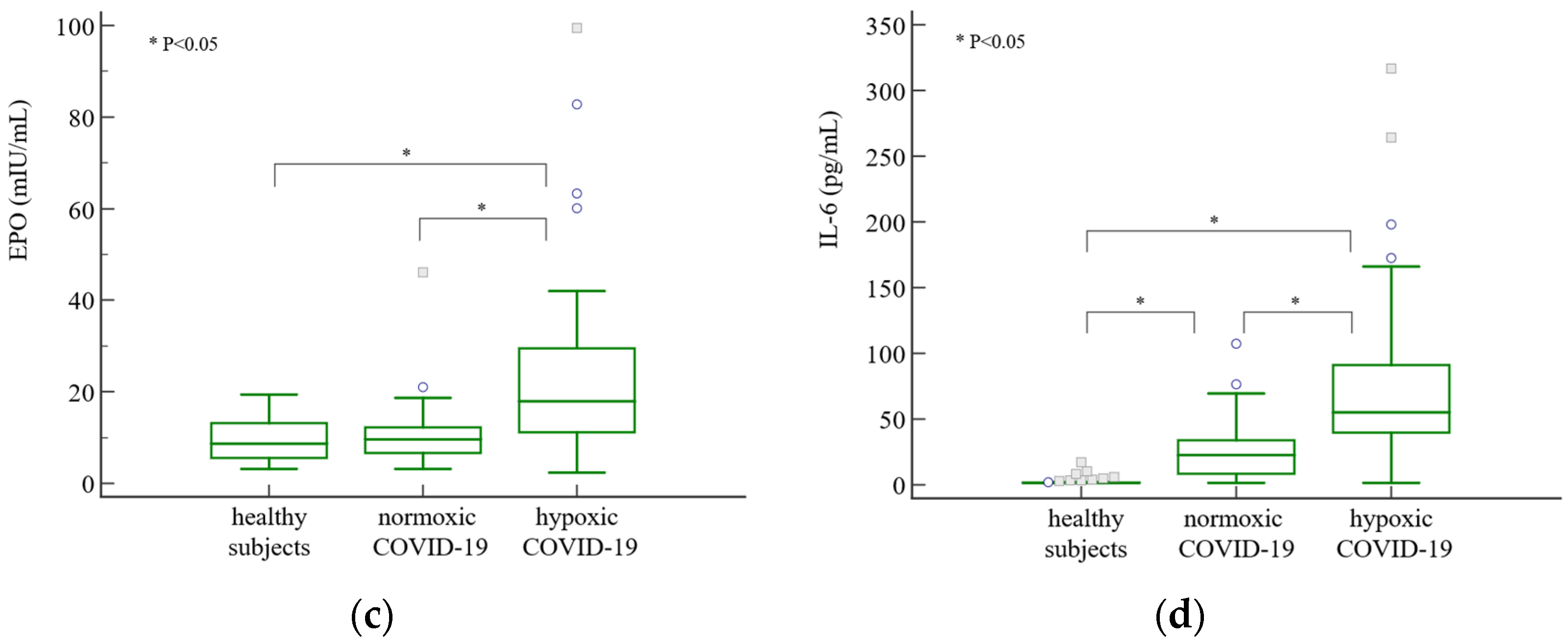
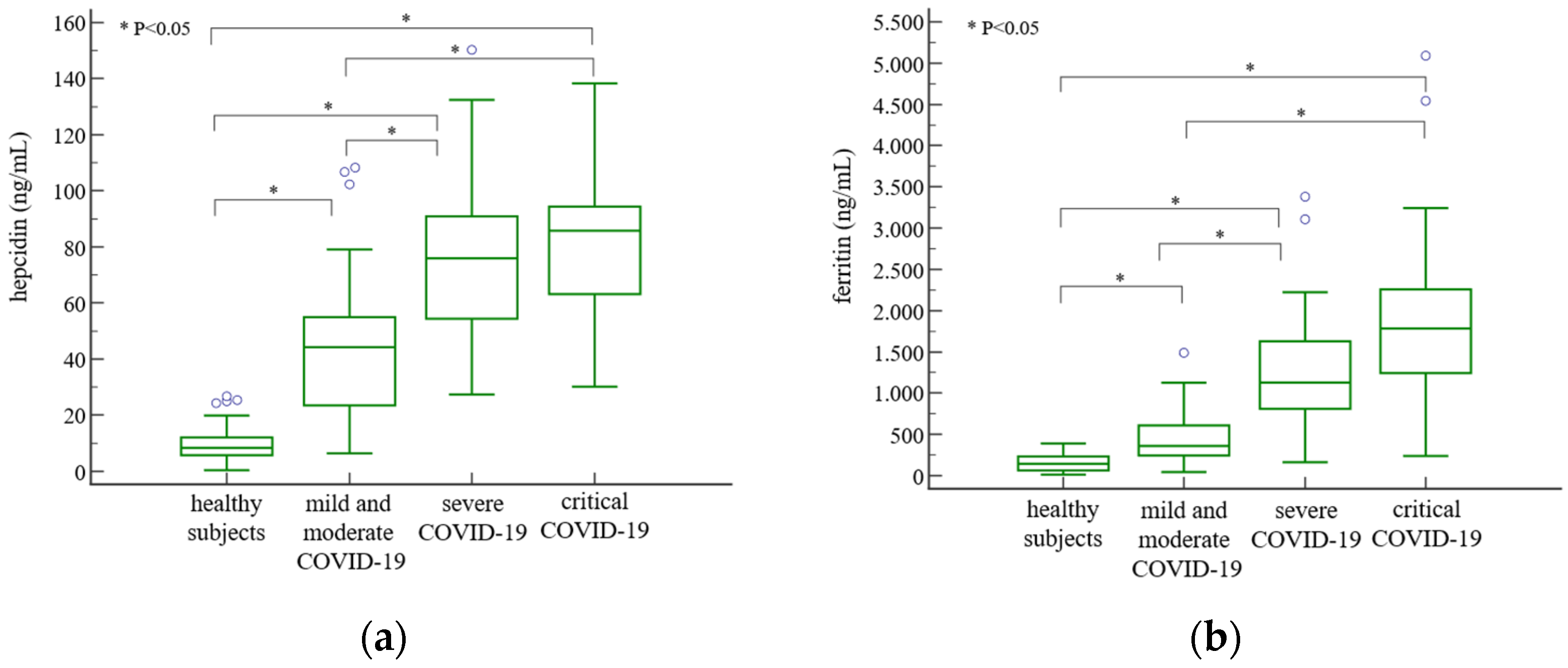
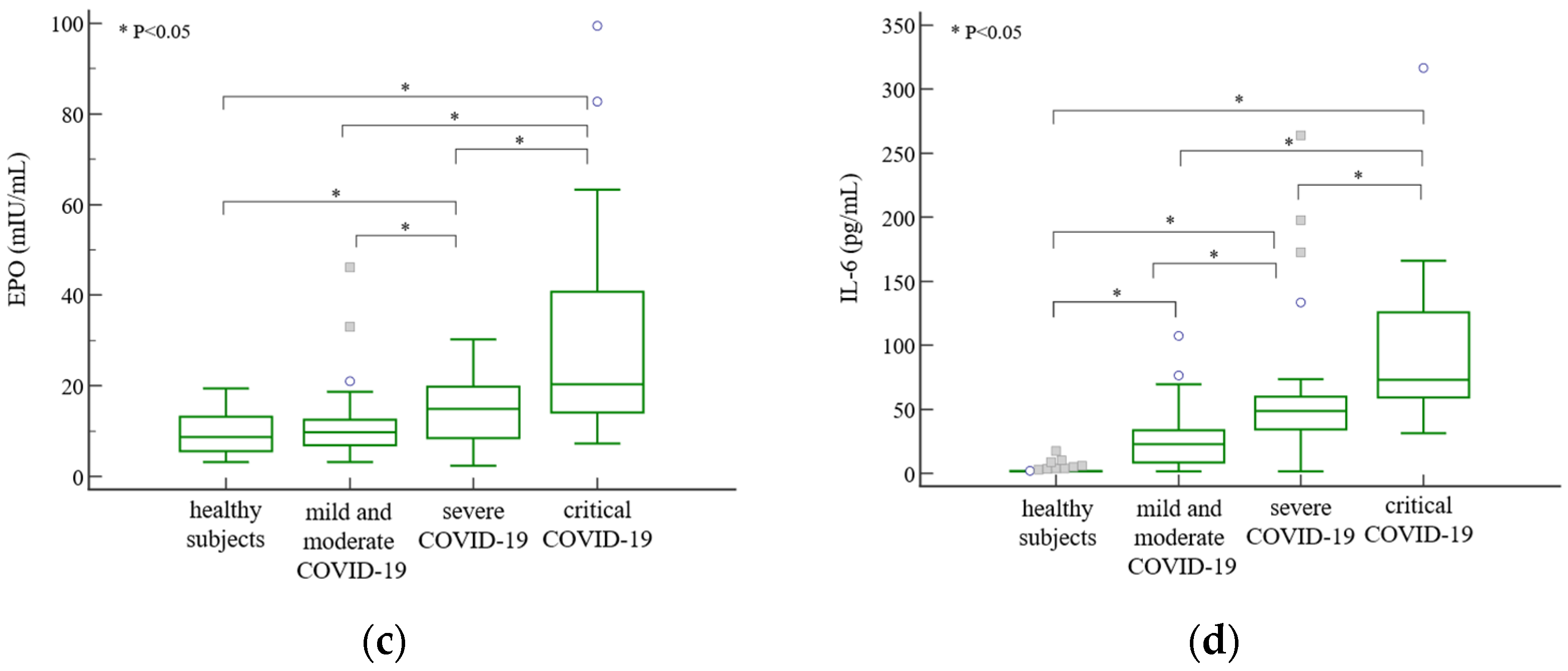
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Domenico, I.; Ward, D.M.; Langelier, C.; Vaughn, M.B.; Nemeth, E.; Sundquist, W.I.; Ganz, T.; Musci, G.; Kaplan, J. The Molecular Mechanism of Hepcidin-Mediated Ferroportin Down-Regulation. Mol. Biol. Cell 2007, 18, 2569–2578. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Nemeth, E. Hepcidin and Iron Homeostasis. Biochim. Biophys. Acta BBA Mol. Cell Res. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Vogt, A.-C.S.; Arsiwala, T.; Mohsen, M.; Vogel, M.; Manolova, V.; Bachmann, M.F. On Iron Metabolism and Its Regulation. Int. J. Mol. Sci. 2021, 22, 4591. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G. Iron and Immunity: A Double-Edged Sword. Eur. J. Clin. Investig. 2002, 32 (Suppl. S1), 70–78. [Google Scholar] [CrossRef] [PubMed]
- Schaible, U.E.; Kaufmann, S.H.E. Iron and Microbial Infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.-S.; Enns, C.A. Molecular Mechanisms of Normal Iron Homeostasis. Hematology 2009, 2009, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Sangkhae, V.; Nemeth, E. Regulation of the Iron Homeostatic Hormone Hepcidin. Adv. Nutr. 2017, 8, 126–136. [Google Scholar] [CrossRef]
- Hennigar, S.R.; McClung, J.P. Nutritional Immunity: Starving Pathogens of Trace Minerals. Am. J. Lifestyle Med. 2016, 10, 170–173. [Google Scholar] [CrossRef]
- Shah, M.D.; Sumeh, A.S.; Sheraz, M.; Kavitha, M.S.; Venmathi Maran, B.A.; Rodrigues, K.F. A Mini-Review on the Impact of COVID 19 on Vital Organs. Biomed. Pharmacother. 2021, 143, 112158. [Google Scholar] [CrossRef]
- Molina-Mora, J.A.; González, A.; Jiménez-Morgan, S.; Cordero-Laurent, E.; Brenes, H.; Soto-Garita, C.; Sequeira-Soto, J.; Duarte-Martínez, F. Clinical Profiles at the Time of Diagnosis of SARS-CoV-2 Infection in Costa Rica during the Pre-Vaccination Period Using a Machine Learning Approach. Phenomics 2022, 2, 312–322. [Google Scholar] [CrossRef]
- Xie, J.; Covassin, N.; Fan, Z.; Singh, P.; Gao, W.; Li, G.; Kara, T.; Somers, V.K. Association between Hypoxemia and Mortality in Patients with COVID-19. Mayo Clin. Proc. 2020, 95, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Gustine, J.N.; Jones, D. Immunopathology of Hyperinflammation in COVID-19. Am. J. Pathol. 2021, 191, 4–17. [Google Scholar] [CrossRef]
- Armitage, A.E.; Stacey, A.R.; Giannoulatou, E.; Marshall, E.; Sturges, P.; Chatha, K.; Smith, N.M.G.; Huang, X.; Xu, X.; Pasricha, S.-R.; et al. Distinct Patterns of Hepcidin and Iron Regulation during HIV-1, HBV, and HCV Infections. Proc. Natl. Acad. Sci. USA 2014, 111, 12187–12192. [Google Scholar] [CrossRef]
- Chakurkar, V.; Rajapurkar, M.; Lele, S.; Mukhopadhyay, B.; Lobo, V.; Injarapu, R.; Sheikh, M.; Dholu, B.; Ghosh, A.; Jha, V. Increased Serum Catalytic Iron May Mediate Tissue Injury and Death in Patients with COVID-19. Sci. Rep. 2021, 11, 19618. [Google Scholar] [CrossRef] [PubMed]
- Hippchen, T.; Altamura, S.; Muckenthaler, M.U.; Merle, U. Hypoferremia Is Associated with Increased Hospitalization and Oxygen Demand in COVID-19 Patients. HemaSphere 2020, 4, e492. [Google Scholar] [CrossRef] [PubMed]
- Sonnweber, T.; Boehm, A.; Sahanic, S.; Pizzini, A.; Aichner, M.; Sonnweber, B.; Kurz, K.; Koppelstätter, S.; Haschka, D.; Petzer, V.; et al. Persisting Alterations of Iron Homeostasis in COVID-19 Are Associated with Non-Resolving Lung Pathologies and Poor Patients’ Performance: A Prospective Observational Cohort Study. Respir. Res. 2020, 21, 276. [Google Scholar] [CrossRef]
- Sukhomlin, T. Hepcidin Is a Friend Rather than a Foe in COVID19-Induced Complications. Acta Biomed. Atenei Parm. 2020, 91, e2020138. [Google Scholar] [CrossRef]
- Suriawinata, E.; Mehta, K.J. Iron and Iron-Related Proteins in COVID-19. Clin. Exp. Med. 2022, 23, 969–991. [Google Scholar] [CrossRef]
- Nemeth, E.; Rivera, S.; Gabayan, V.; Keller, C.; Taudorf, S.; Pedersen, B.K.; Ganz, T. IL-6 Mediates Hypoferremia of Inflammation by Inducing the Synthesis of the Iron Regulatory Hormone Hepcidin. J. Clin. Investig. 2004, 113, 1271–1276. [Google Scholar] [CrossRef]
- Huang, H.; Constante, M.; Layoun, A.; Santos, M.M. Contribution of STAT3 and SMAD4 Pathways to the Regulation of Hepcidin by Opposing Stimuli. Blood 2009, 113, 3593–3599. [Google Scholar] [CrossRef] [PubMed]
- Nai, A.; Lorè, N.I.; Pagani, A.; De Lorenzo, R.; Di Modica, S.; Saliu, F.; Cirillo, D.M.; Rovere-Querini, P.; Manfredi, A.A.; Silvestri, L. Hepcidin Levels Predict COVID-19 Severity and Mortality in a Cohort of Hospitalized Italian Patients. Am. J. Hematol. 2021, 96, E32–E35. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Chen, Y.; Ji, Y.; He, X.; Xue, D. Increased Serum Levels of Hepcidin and Ferritin Are Associated with Severity of COVID-19. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e926178-1–e926178-6. [Google Scholar] [CrossRef] [PubMed]
- Al-Janabi, G.; Al-Fahham, A.; Alsaedi, A.N.N.; Al-Amery, A.Y.K. Correlation between hepcidin and procalcitonin and their diagnostic role in patients with COVID-19. Wiad. Lek. 2023, 76, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, N.; Bayraktar, M.; Ozturk, A.; Ibrahim, B. Evaluation of the Relationship Between Aquaporin-1, Hepcidin, Zinc, Copper, and İron Levels and Oxidative Stress in the Serum of Critically Ill Patients with COVID-19. Biol. Trace Elem. Res. 2022, 200, 5013–5021. [Google Scholar] [CrossRef] [PubMed]
- Yağcı, S.; Serin, E.; Acicbe, Ö.; Zeren, M.İ.; Odabaşı, M.S. The Relationship between Serum Erythropoietin, Hepcidin, and Haptoglobin Levels with Disease Severity and Other Biochemical Values in Patients with COVID-19. Int. J. Lab. Hematol. 2021, 43, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Ciotti, M.; Nuccetelli, M.; Pieri, M.; Petrangeli, C.M.; Giovannelli, A.; Cosio, T.; Rosa, L.; Valenti, P.; Leonardis, F.; Legramante, J.M.; et al. Evaluation of Hepcidin Level in COVID-19 Patients Admitted to the Intensive Care Unit. Diagnostics 2022, 12, 2665. [Google Scholar] [CrossRef]
- Piperno, A.; Galimberti, S.; Mariani, R.; Pelucchi, S.; Ravasi, G.; Lombardi, C.; Bilo, G.; Revera, M.; Giuliano, A.; Faini, A.; et al. Modulation of Hepcidin Production during Hypoxia-Induced Erythropoiesis in Humans In Vivo: Data from the HIGHCARE Project. Blood 2011, 117, 2953–2959. [Google Scholar] [CrossRef]
- Goetze, O.; Schmitt, J.; Spliethoff, K.; Theurl, I.; Weiss, G.; Swinkels, D.W.; Tjalsma, H.; Maggiorini, M.; Krayenbühl, P.; Rau, M.; et al. Adaptation of Iron Transport and Metabolism to Acute High-Altitude Hypoxia in Mountaineers. Hepatology 2013, 58, 2153–2162. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron Balance and the Role of Hepcidin in Chronic Kidney Disease. Semin. Nephrol. 2016, 36, 87–93. [Google Scholar] [CrossRef]
- Kowdley, K.V.; Gochanour, E.M.; Sundaram, V.; Shah, R.A.; Handa, P. Hepcidin Signaling in Health and Disease: Ironing Out the Details. Hepatol. Commun. 2021, 5, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Coffey, R.; Ganz, T. Erythroferrone: An Erythroid Regulator of Hepcidin and Iron Metabolism. HemaSphere 2018, 2, e35. [Google Scholar] [CrossRef] [PubMed]
- Living Guidance for Clinical Management of COVID-19. Available online: https://www.who.int/publications-detail-redirect/WHO-2019-nCoV-clinical-2021-2 (accessed on 29 August 2023).
- Agustí, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E.; Vestbo, J.; Lomas, D.A.; Calverley, P.M.A.; Wouters, E.; et al. Persistent Systemic Inflammation Is Associated with Poor Clinical Outcomes in COPD: A Novel Phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef] [PubMed]
- Hortová-Kohoutková, M.; Skotáková, M.; Onyango, I.G.; Slezáková, M.; Panovský, R.; Opatřil, L.; Slanina, P.; De Zuani, M.; Mrkva, O.; Andrejčinová, I.; et al. Hepcidin and Ferritin Levels as Markers of Immune Cell Activation during Septic Shock, Severe COVID-19 and Sterile Inflammation. Front. Immunol. 2023, 14, 1110540. [Google Scholar] [CrossRef] [PubMed]
- Hegelund, M.H.; Glenthøj, A.; Ryrsø, C.K.; Ritz, C.; Dungu, A.M.; Sejdic, A.; List, K.C.K.; Krogh-Madsen, R.; Lindegaard, B.; Kurtzhals, J.A.L.; et al. Biomarkers for Iron Metabolism among Patients Hospitalized with Community-Acquired Pneumonia Caused by Infection with SARS-CoV-2, Bacteria, and Influenza. Acta Pathol. Microbiol. Immunol. Scand. 2022, 130, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Elbostany, E.A.; Elghoroury, E.A.; Thabet, E.H.; Rashad, A.A.; Rasheed, E.A.; El-Saeed, G.S.M.; Abdelhalim, D.A.; Abdelfattah, S.N.; Salama, I.I.; Salama, N. Oxidative Stress and Hepcidin Expression in Pediatric Sickle Cell Anemia with Iron Overload. Hematol. Oncol. Stem Cell Ther. 2023, 16, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Luque-Ramírez, M.; Álvarez-Blasco, F.; Alpañés, M.; Escobar-Morreale, H.F. Role of Decreased Circulating Hepcidin Concentrations in the Iron Excess of Women with the Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2011, 96, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Heeney, M.M.; Guo, D.; De Falco, L.; Campagna, D.R.; Olbina, G.; Kao, P.P.-C.; Schmitz-Abe, K.; Rahimov, F.; Gutschow, P.; Westerman, K.; et al. Normalizing Hepcidin Predicts TMPRSS6 Mutation Status in Patients with Chronic Iron Deficiency. Blood 2018, 132, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Korenaga, M.; Nishina, S.; Korenaga, K.; Tomiyama, Y.; Yoshioka, N.; Hara, Y.; Sasaki, Y.; Shimonaka, Y.; Hino, K. Branched-Chain Amino Acids Reduce Hepatic Iron Accumulation and Oxidative Stress in Hepatitis C Virus Polyprotein-Expressing Mice. Liver Int. 2015, 35, 1303–1314. [Google Scholar] [CrossRef]
- Evanchuk, J.L.; Kozyrskyj, A.; Hanas, N.; Goruk, S.; Vaghef-Mehrabani, E.; Archundia-Herrera, C.M.; O’Brien, K.O.; Letourneau, N.L.; Giesbrecht, G.F.; Bell, R.C.; et al. Maternal Iron Status Is Dynamic Throughout Pregnancy and Might Predict Birth Outcomes in a Sex Dependent Manner: Results from the Alberta Pregnancy Outcomes and Nutrition (APrON) Cohort Study. J. Nutr. 2023, 153, 2585–2597. [Google Scholar] [CrossRef]
- Wei, S.; Liu, W.; Qi, Y.; Guo, Y.; Zhang, S.; Wang, L.; Zhuang, T.; Zhang, N.; Liu, S. Disordered Serum Erythroferrone and Hepcidin Levels as Indicators of the Spontaneous Abortion Occurrence during Early Pregnancy in Humans. Br. J. Haematol. 2021, 192, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, S. COVID-19 and Iron Dysregulation: Distant Sequence Similarity between Hepcidin and the Novel Coronavirus Spike Glycoprotein. Biol. Direct 2020, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Maira, D.; Duca, L.; Busti, F.; Consonni, D.; Salvatici, M.; Vianello, A.; Milani, A.; Guzzardella, A.; Di Pierro, E.; Aliberti, S.; et al. The Role of Hypoxia and Inflammation in the Regulation of Iron Metabolism and Erythropoiesis in COVID-19: The IRONCOVID Study. Am. J. Hematol. 2022, 97, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.-R.; McHugh, K.; Drakesmith, H. Regulation of Hepcidin by Erythropoiesis: The Story So Far. Annu. Rev. Nutr. 2016, 36, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Buttarello, M. Laboratory Diagnosis of Anemia: Are the Old and New Red Cell Parameters Useful in Classification and Treatment, How? Int. J. Lab. Hematol. 2016, 38 (Suppl. S1), 123–132. [Google Scholar] [CrossRef] [PubMed]
- Ashby, D.R.; Gale, D.P.; Busbridge, M.; Murphy, K.G.; Duncan, N.D.; Cairns, T.D.; Taube, D.H.; Bloom, S.R.; Tam, F.W.K.; Chapman, R.; et al. Erythropoietin Administration in Humans Causes a Marked and Prolonged Reduction in Circulating Hepcidin. Haematologica 2010, 95, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.; Lopez, M.A.; Gabayan, V.; Ganz, T.; Rivera, S. Suppression of Hepcidin during Anemia Requires Erythropoietic Activity. Blood 2006, 108, 3730–3735. [Google Scholar] [CrossRef]
- Ahmed, S.; Ahmed, Z.A.; Siddiqui, I.; Rashid, N.H.; Mansoor, M.; Jafri, L. Evaluation of Serum Ferritin for Prediction of Severity and Mortality in COVID-19—A Cross Sectional Study. Ann. Med. Surg. 2021, 63, 102163. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Subjects (n = 47) | Total COVID-19 Patients (n = 96) | Normoxic COVID-19 Patients (n = 49) | Hypoxic COVID-19 Patients (n = 47) | P1 | P2 | P3 | |
|---|---|---|---|---|---|---|---|
| age (years) | 58 (39–70) | 54 (36–73) | 53 (36–73) | 55 (37–72) | 0.19 | 0.17 | 0.18 |
| sex male, n female, n | 31 16 | 64 32 | 31 18 | 33 14 | 1.00 | 0.77 | 0.52 |
| SpO2 (%) | / | 96 (88–98) | 97 (97–98) | 87 (83–89) | / | / | <0.001 |
| creatinine (µmol/L) | 77 ± 12 | 79 ± 14 | 80 ± 15 | 78 ± 14 | 0.29 | 0.46 | 0.48 |
| eGFR (mL/min/1.73 m2) | 91 (80–99) | 87 (77–101) | 86 (78–99) | 88 (77–102) | 0.52 | 0.70 | 0.61 |
| symptoms duration on admission (days), n | / | 8 ± 3 | 8 ± 3 | 9 ± 3 | / | / | 0.001 |
| Clinical Manifestations | Normoxic COVID-19 Patients (n = 49) | Hypoxic COVID-19 Patients (n = 47) |
|---|---|---|
| fever before admission | 47 (96) | 46 (98) |
| fever on admission | 28 (57) | 35 (74) |
| cough | 35 (71) | 36 (77) |
| sore throat | 9 (18) | 8 (17) |
| dyspnea | 0 (0) | 35 (74) |
| fatigue | 19 (39) | 32 (68) |
| myalgia | 18 (37) | 17 (36) |
| pneumonia on X-ray imaging | 16 (33) | 47 (100) |
| Parameter | Healthy Subjects (n = 47) | Total COVID-19 Patients (n = 96) | Normoxic COVID-19 Patients (n = 49) | Hypoxic COVID-19 Patients (n = 47) | P1 | P2 |
|---|---|---|---|---|---|---|
| haemoglobin (g/dL) | 15.0 (13.7–15.4) | 14.5 (13.5–15.4) | 14.4 (13.5–15.6) | 14.6 (13.6–15.4) | 0.48 | 0.75 |
| iron (µmol/L) | 17.0 (13.3–21.0) | 6.0 (5.0–8.5) | 6.0 (5.0–8.3) a | 5.0 (4.3–8.8) a | <0.001 | <0.001 |
| UIBC (µmol/L) | 42 (35–49) | 39 (32–45) | 44 (37–47) | 33 (29–40) a,b | 0.06 | <0.001 |
| TIBC (µmol/L) | 59 (54–68) | 46 (40–51) | 50 (45–54) a | 42 (37–46) a,b | <0.001 | <0.001 |
| TSAT (%) | 28 (23–36) | 13 (10–18) | 12 (10–16) a | 15 (10–20) a | <0.001 | <0.001 |
| sTfR (mg/L) | 1.09 (1.00–1.25) | 1.26 (1.07–1.45) | 1.30 (1.12–1.49) a | 1.23 (1.02–1.41) a | 0.002 | 0.002 |
| RET-He (pg) | 33.0 (32.4–33.7) | 30.1 (28.6–31.5) | 30.2 (28.8–31.0) a | 30.1 (28.4–31.9) a | <0.001 | <0.001 |
| RTC (‰) | 13.20 (11.70–15.58) | 5.25 (4.40–6.85) | 5.00 (4.18–6.98) a | 5.30 (4.43–6.80) a | <0.001 | <0.001 |
| RTC (×109/L) | 67.60 (56.05–82.05) | 26.10 (20.90–34.80) | 25.60 (19.45–35.05) a | 26.20 (21.80–34.63) a | <0.001 | <0.001 |
| IRF (%) | 8.9 (7.5–10.3) | 4.6 (3.4–7.9) | 3.6 (3.0–4.9) a | 6.2 (4.1–10.7) a,b | <0.001 | <0.001 |
| CRP (mg/L) | 1.2 (0.7–2.3) | 37.1 (12.7–100.0) | 13.1 (5.2–33.2) a | 85.5 (39.7–120.7) a,b | <0.001 | <0.001 |
| ferritin/hepcidin | 15.17 (9.84–25.09) | 13.55 (7.06–20.91) | 7.72 (6.24–13.98) a | 19.02 (12.98–29.64) b | 0.10 | <0.001 |
| hepcidin/iron | 0.49 (0.34–0.84) | 10.24 (5.04–14.05) | 7.37 (3.44–12.00) a | 13.04 (8.06–16.04) a,b | <0.001 | <0.001 |
| hepcidin /CRP | 7.14 (2.70–12.16) | 1.67 (0.77–3.25) | 2.58 (1.39–6.42) a | 0.93 (0.53–1.80) a,b | <0.001 | <0.001 |
| hepcidin/IL-6 | 4.54 (2.50–7.02) | 1.57 (1.00–2.45 | 1.94 (1.42–3.69) a | 1.33 (0.61–1.92) a,b | <0.001 | <0.001 |
| hepcidin/EPO | 1.13 (0.57–2.14) | 4.09 (2.32–7.60) | 4.23 (2.36–7.81) a | 4.08 (2.26–7.37) a | <0.001 | <0.001 |
| Parameter | Healthy Subjects (n = 47) | Mild and Moderate COVID-19 (n = 49) | Severe COVID-19 (n = 28) | Critical COVID-19 (n = 19) | p |
|---|---|---|---|---|---|
| haemoglobin (g/dL) | 15.0 (13.7–15.4) | 14.3 (13.4–15.6) | 14.5 (13.8–15.4) | 14.7 (13.6–15.2) | 0.87 |
| iron (µmol/L) | 17.0 (13.3–21.0) | 6.0 (5.0–8.0) a | 6.0 (5.0–9.0) a | 5.0 (4.3–8.8) a | <0.001 |
| UIBC (µmol/L) | 42 (35–49) | 44 (36–47) | 34 (29–39) a,b | 33 (29–41) a,b | <0.001 |
| TIBC (µmol/L) | 59 (54–68) | 49 (44–53) a | 43 (38–47) a,b | 41 (37–47) a,b | <0.001 |
| TSAT (%) | 28 (23–36) | 12 (10–15) a | 15 (11–19) a | 16 (10–21) a | <0.001 |
| sTfR (mg/L) | 1.09 (1.00–1.25) | 1.29 (11.12–1.48) a | 1.23 (1.06–1.45) a | 1.23 (1.01–1.40) | 0.010 |
| RET-He (pg) | 33.0 (32.4–33.7) | 30.2 (28.6–31.0) a | 30.1 (28.5–31.8) a | 30.1 (28.7–31.6) a | <0.001 |
| RTC (‰) | 13.20 (11.70–15.58) | 5.20 (4.18–6.98) a | 5.45 (4.50–6.80) a | 4.90 (4.40–6.63) a | <0.001 |
| RTC (×109/L) | 67.60 (56.05–82.05) | 25.60 (19.45–35.58) a | 26.75 (22.05–35.05) a | 26.10 (20.90–33.38) a | <0.001 |
| IRF (%) | 8.9 (7.5–10.3) | 3.6 (3.1–5.4) a | 6.5 (4.4.–11.0) a,b | 4.9 (3.5–10.0) a,b | <0.001 |
| CRP (mg/L) | 1.2 (0.7–2.3) | 14.3 (5.2–35.2) a | 84.0 (38.1–120.7) a,b | 86.7 (42.1–129.9) a,b | <0.001 |
| ferritin/hepcidin | 15.17 (9.84–25.09) | 7.72 (6.26–14.19) a | 16.78 (11.94–24.23) b | 23.16 (13.83–33.23) b | <0.001 |
| hepcidin/iron | 0.49 (0.34–0.84) | 7.37 (3.44–12.00) a | 12.40 (8.08–15.50) a,b | 13.04 (9.86–16.92) a,b | <0.001 |
| hepcidin /CRP | 7.14 (2.70–12.16) | 2.49 (1.34–6.42) a | 1.03 (0.61–1.76) a,b | 0.87 (0.43–1.98) a,b | <0.001 |
| hepcidin/IL-6 | 4.54 (2.50–7.02) | 1.85 (1.33–3.27) a | 1.47 (0.75–2.07) a,b | 1.39 (0.59–1.64) a,b | <0.001 |
| hepcidin/EPO | 1.13 (0.57–2.14) | 4.07 (2.13–6.89) a | 5.40 (3.21–7.86) a | 3.29 (1.51–6.42) a,c | <0.001 |
| Odds for Critical COVID-19 | |||
|---|---|---|---|
| Parameter | OR | 95% CI | p |
| haemoglobin (g/L) | 1.01 | 0.97–1.05 | 0.73 |
| hepcidin (ng/mL) | 1.03 | 1.01–1.05 | 0.004 * |
| iron (µmol/L) | 0.96 | 0.83–1.11 | 0.58 |
| UIBC (µmol/L) | 0.92 | 0.86–0.99 | 0.028 * |
| TIBC (µmol/L) | 0.91 | 0.85–0.98 | 0.013 * |
| TSAT (%) | 1.01 | 0.95–1.08 | 0.69 |
| ferritin (ng/mL) | 1.00 | 1.00–1.00 | <0.001 * |
| sTfR (mg/L) | 0.93 | 0.16–5.50 | 0.94 |
| RET-He (pg) | 1.03 | 0.80–1.33 | 0.82 |
| RTC (‰) | 1.02 | 0.87–1.21 | 0.78 |
| RTC (×109/L) | 1.01 | 0.98–1.04 | 0.54 |
| IRF (%) | 1.07 | 0.97–1.18 | 0.16 |
| EPO (mIU/mL) | 1.10 | 1.04–1.16 | <0.001 * |
| CRP (mg/L) | 1.01 | 1.00–1.03 | 0.005 * |
| IL-6 (pg/mL) | 1.02 | 1.01–1.03 | 0.001 * |
| ferritin/hepcidin | 1.08 | 1.03–1.13 | 0.001 * |
| hepcidin/iron | 1.07 | 1.00–1.15 | 0.037 * |
| hepcidin /CRP | 0.80 | 0.61–1.06 | 0.12 |
| hepcidin/IL-6 | 0.63 | 0.37–1.08 | 0.10 |
| hepcidin/EPO | 0.94 | 0.83–1.07 | 0.34 |
| Odds for Critical COVID-19 | |||
|---|---|---|---|
| Parameter | OR | 95% CI | p |
| EPO | 1.10 | 1.04–1.16 | <0.001 |
| ferritin/hepcidin | 1.08 | 1.02–1.14 | 0.004 |
| The analysis gave results with 88% of correctly classified cases and AUC of 0.838 (0.749–0.905). | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugo, K.; Tandara, L.; Juricic, G.; Pavicic Ivelja, M.; Rumora, L. Effects of Hypoxia and Inflammation on Hepcidin Concentration in Non-Anaemic COVID-19 Patients. J. Clin. Med. 2024, 13, 3201. https://doi.org/10.3390/jcm13113201
Gugo K, Tandara L, Juricic G, Pavicic Ivelja M, Rumora L. Effects of Hypoxia and Inflammation on Hepcidin Concentration in Non-Anaemic COVID-19 Patients. Journal of Clinical Medicine. 2024; 13(11):3201. https://doi.org/10.3390/jcm13113201
Chicago/Turabian StyleGugo, Katarina, Leida Tandara, Gordana Juricic, Mirela Pavicic Ivelja, and Lada Rumora. 2024. "Effects of Hypoxia and Inflammation on Hepcidin Concentration in Non-Anaemic COVID-19 Patients" Journal of Clinical Medicine 13, no. 11: 3201. https://doi.org/10.3390/jcm13113201
APA StyleGugo, K., Tandara, L., Juricic, G., Pavicic Ivelja, M., & Rumora, L. (2024). Effects of Hypoxia and Inflammation on Hepcidin Concentration in Non-Anaemic COVID-19 Patients. Journal of Clinical Medicine, 13(11), 3201. https://doi.org/10.3390/jcm13113201