
The Manganese–Bone Connection: Investigating the Role of Manganese in Bone Health
 ,
,
Abstract
:1. Introduction
2. Historical Overview: Early Discoveries on the Relationship between Mn and Bone Health
3. The Influence of Mn Metabolism on Bone Mass

4. The Role of Mn in Bone Development and Remodeling
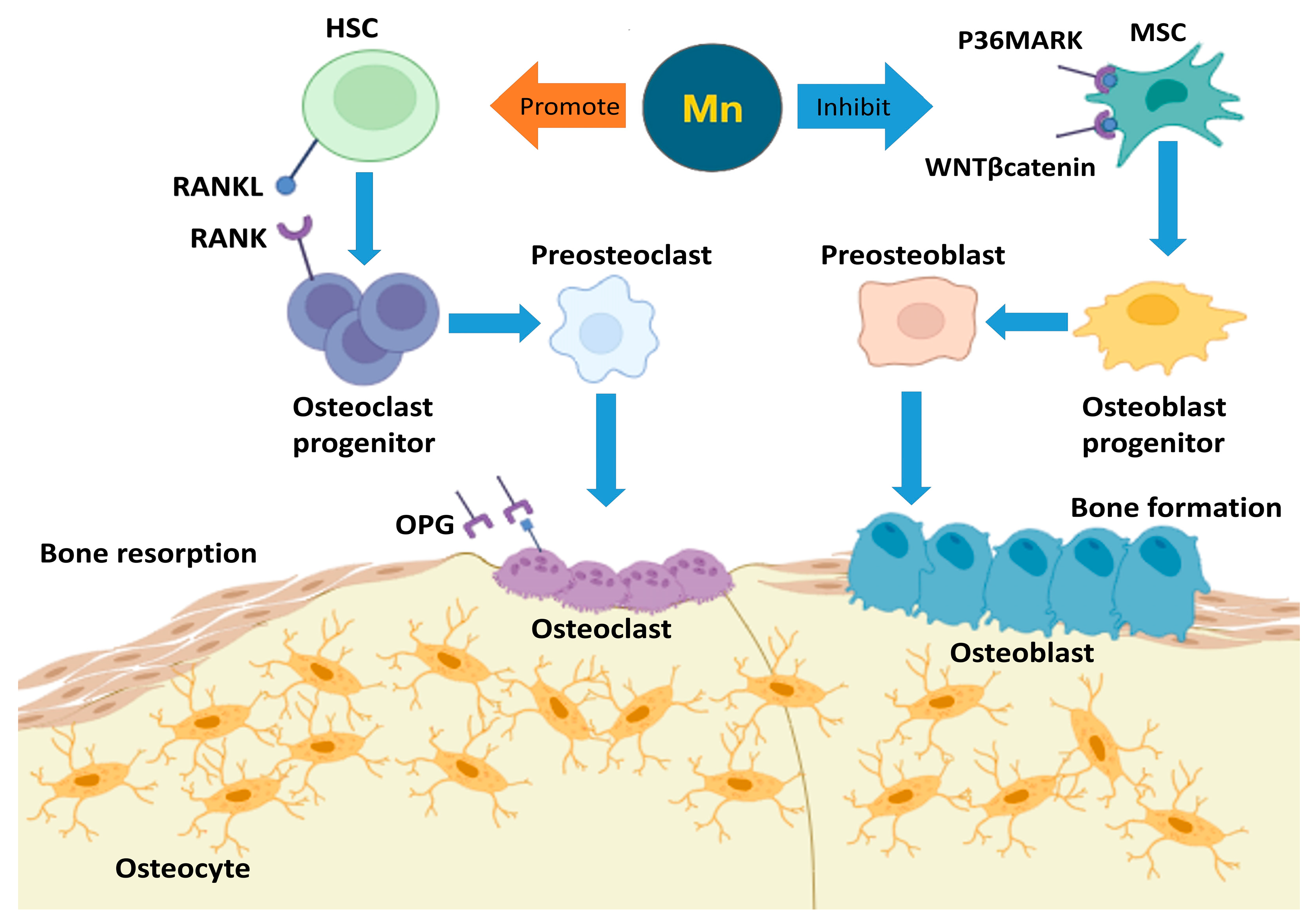
5. Mn Absorption and Distribution in the Body
6. Mn Homeostasis and Regulation in Bone Tissue
7. Mechanisms of Mn Action on Bone Cells
8. Mn Deficiency and Its Impact on Bone Health
9. Mn Toxicity and Its Effects on Bone Mass
10. Animal Studies Investigating the Relationship between Mn and Bone Density
11. Clinical Studies Assessing Mn Supplementation and Bone Health
12. Future Directions: Areas for Further Research and Investigation
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sromova, V.; Sobola, D.; Kaspar, P. A Brief Review of Bone Cell Function and Importance. Cells 2023, 12, 2576. [Google Scholar] [CrossRef] [PubMed]
- Vanitchanont, M.; Vallibhakara, S.A.; Sophonsritsuk, A.; Vallibhakara, O. Effects of Multispecies Probiotic Supplementation on Serum Bone Turnover Markers in Postmenopausal Women with Osteopenia: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2024, 16, 461. [Google Scholar] [CrossRef]
- Ciosek, Z.; Kot, K.; Kosik-Bogacka, D.; Lanocha-Arendarczyk, N.; Rotter, I. The Effects of Calcium, Magnesium, Phosphorus, Fluoride, and Lead on Bone Tissue. Biomolecules 2021, 11, 506. [Google Scholar] [CrossRef] [PubMed]
- Lopes, T.S.B.; Shi, H.; White, D.; Araujo, I.C.S.; Kim, W.K. Effects of 25-hydroxycholecalciferol on performance, gut health, and bone quality of broilers fed with reduced calcium and phosphorus diet during Eimeria challenge. Poult. Sci. 2024, 103, 103267. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.R.; Li, L.; Nie, L.X. Associations between co-exposure to heavy metals and vertebral compression fracture, as well as femoral neck bone mineral density: A cross-sectional study from NHANES data. PLoS ONE 2024, 19, e0303418. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, Y.; Long, H.T.; Ou, M.N.; Zhao, S.S. Relationship between blood manganese and bone mineral density and bone mineral content in adults: A population-based cross-sectional study. PLoS ONE 2022, 17, e0276551. [Google Scholar] [CrossRef]
- Wei, M.; Huang, Q.; Dai, Y.; Zhou, H.; Cui, Y.; Song, W.; Di, D.; Zhang, R.; Li, C.; Wang, Q.; et al. Manganese, iron, copper, and selenium co-exposure and osteoporosis risk in Chinese adults. J. Trace Elem. Med. Biol. 2022, 72, 126989. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Kishore, V.; Bose, M.; Nandi, S.K.; Roy, M. In vitro and in vivo degradability, biocompatibility and antimicrobial characteristics of Cu added iron-manganese alloy. J. Mater. Sci. Technol. 2021, 84, 159–172. [Google Scholar] [CrossRef]
- Sirri, F.; Maiorano, G.; Tavaniello, S.; Chen, J.; Petracci, M.; Meluzzi, A. Effect of different levels of dietary zinc, manganese, and copper from organic or inorganic sources on performance, bacterial chondronecrosis, intramuscular collagen characteristics, and occurrence of meat quality defects of broiler chickens. Poult. Sci. 2016, 95, 1813–1824. [Google Scholar] [CrossRef]
- Von Oettingen, W. Manganese: Its distribution, pharmacology and health hazards. Physiol. Rev. 1935, 15, 175–201. [Google Scholar] [CrossRef]
- Christie, T. Mineral Commodity Report 7—Manganese; Institute of Geological and Nuclear Sciences Ltd.: Avalon, New Zealand, 2007. [Google Scholar]
- Chukanov, N.V.; Varlamov, D.A.; Pekov, I.V.; Zubkova, N.V.; Kasatkin, A.V.; Britvin, S.N. Coupled Substitutions in Natural MnO(OH) Polymorphs: Infrared Spectroscopic Investigation. Minerals 2021, 11, 969. [Google Scholar] [CrossRef]
- Ghosh, S.K. Diversity in the Family of Manganese Oxides at the Nanoscale: From Fundamentals to Applications. ACS Omega 2020, 5, 25493–25504. [Google Scholar] [CrossRef] [PubMed]
- Kemmerer, A.; Elvehjem, C.; Hart, E. Studies on the relation of manganese to the nutrition of the mouse. J. Biol. Chem. 1931, 92, 623–630. [Google Scholar] [CrossRef]
- Wilgus, J.H.S.; Patton, A.R. Factors Affecting Manganese Utilization in the Chicken. J. Nutr. 1939, 18, 35–45. [Google Scholar] [CrossRef]
- Friedman, B.J.; Freeland-Graves, J.H.; Bales, C.W.; Behmardi, F.; Shorey-Kutschke, R.L.; Willis, R.A.; Crosby, J.B.; Trickett, P.C.; Houston, S.D. Manganese balance and clinical observations in young men fed a manganese-deficient diet. J. Nutr. 1987, 117, 133–143. [Google Scholar] [CrossRef]
- Stern, P.H. Biphasic effects of manganese on hormone-stimulated bone resorption. Endocrinology 1985, 117, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Kehoe, R.A.; Cholak, J.; Storey, R. Spectrochemical study of the normal ranges of concentration of certain trace metals in biological materials. J. Nutr. 1940, 19, 579–592. [Google Scholar] [CrossRef]
- Chen, P.; Bornhorst, J.; Aschner, M.A. Manganese Metabolism in Humans. In Postprints Der Universität Potsdam Mathematisch-Naturwissenschaftliche Reihe; Mathematisch-Naturwissenschaftliche Fakult: Zürich, Switzerland, 2019. [Google Scholar]
- Horning, K.J.; Caito, S.W.; Tipps, K.G.; Bowman, A.B.; Aschner, M. Manganese Is Essential for Neuronal Health. Annu. Rev. Nutr. 2015, 35, 71–108. [Google Scholar] [CrossRef]
- Baj, J.; Flieger, W.; Barbachowska, A.; Kowalska, B.; Flieger, M.; Forma, A.; Teresinski, G.; Portincasa, P.; Buszewicz, G.; Radzikowska-Buchner, E.; et al. Consequences of Disturbing Manganese Homeostasis. Int. J. Mol. Sci. 2023, 24, 14959. [Google Scholar] [CrossRef]
- Tinkov, A.A.; Paoliello, M.M.B.; Mazilina, A.N.; Skalny, A.V.; Martins, A.C.; Voskresenskaya, O.N.; Aaseth, J.; Santamaria, A.; Notova, S.V.; Tsatsakis, A.; et al. Molecular Targets of Manganese-Induced Neurotoxicity: A Five-Year Update. Int. J. Mol. Sci. 2021, 22, 4646. [Google Scholar] [CrossRef]
- Fernandes, J.; Hao, L.; Bijli, K.M.; Chandler, J.D.; Orr, M.; Hu, X.; Jones, D.P.; Go, Y.M. From the Cover: Manganese Stimulates Mitochondrial H2O2 Production in SH-SY5Y Human Neuroblastoma Cells Over Physiologic as well as Toxicologic Range. Toxicol. Sci. 2017, 155, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Prasadh, S.; Gupta, M.; Wong, R. In vitro cytotoxicity and osteogenic potential of quaternary Mg-2Zn-1Ca/X-Mn alloys for craniofacial reconstruction. Sci. Rep. 2022, 12, 8259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wu, J.; Zhang, S.; Ni, X.; Wang, B.; Lu, K.; Zhang, P.; Xu, R. Preparation and properties of composite manganese/fluorine coatings on metallic titanium. RSC Adv. 2023, 13, 14863–14877. [Google Scholar] [CrossRef] [PubMed]
- Brotto, M.; Bonewald, L. Bone and muscle: Interactions beyond mechanical. Bone 2015, 80, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.; Mijares, D.Q.; Janal, M.N.; Aranya, A.K.; Zhang, D.S.; LeGeros, R.Z.; Zhang, Y. In vivo efficacy of calcium phosphate-based synthetic-bone-mineral on bone loss resulting from estrogen and mineral deficiencies. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 108, 1868–1878. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Du, T.; Niu, X.; Fan, Y. Biomechanics and mechanobiology of the bone matrix. Bone Res. 2022, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.S.; Cabral, J.M.S.; da Silva, C.L.; Vashishth, D. Bone Matrix Non-Collagenous Proteins in Tissue Engineering: Creating New Bone by Mimicking the Extracellular Matrix. Polymers 2021, 13, 1095. [Google Scholar] [CrossRef] [PubMed]
- Farshidfar, N.; Tanideh, N.; Emami, Z.; Aslani, F.S.; Sarafraz, N.; Khodabandeh, Z.; Zare, S.; Farshidfar, G.; Nikoofal-Sahlabadi, S.; Tayebi, L.; et al. Incorporation of curcumin into collagen- multiwalled carbon nanotubes nanocomposite scaffold: An in vitro and in vivo study. J. Mater. Res. Technol.-JmrT 2022, 21, 4558–4576. [Google Scholar] [CrossRef]
- Cumming, M.H.; Hall, B.; Hofman, K. Isolation and Characterisation of Major and Minor Collagens from Hyaline Cartilage of Hoki (Macruronus novaezelandiae). Mar. Drugs 2019, 17, 223. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Y.; He, L.; Wang, Q.; Wang, L.; Yuan, T.; Xiao, Y.; Fan, Y.; Zhang, X. Icariin conjugated hyaluronic acid/collagen hydrogel for osteochondral interface restoration. Acta Biomater. 2018, 74, 156–167. [Google Scholar] [CrossRef]
- Barrioni, B.R.; Norris, E.; Li, S.; Naruphontjirakul, P.; Jones, J.R.; Pereira, M.M. Osteogenic potential of sol-gel bioactive glasses containing manganese. J. Mater. Sci. Mater. Med. 2019, 30, 86. [Google Scholar] [CrossRef]
- Miola, M.; Brovarone, C.V.; Maina, G.; Rossi, F.; Bergandi, L.; Ghigo, D.; Saracino, S.; Maggiora, M.; Canuto, R.A.; Muzio, G.; et al. In vitro study of manganese-doped bioactive glasses for bone regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 38, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, L.I.; Bellido, T. Osteocytic signalling pathways as therapeutic targets for bone fragility. Nat. Rev. Endocrinol. 2016, 12, 593–605. [Google Scholar] [CrossRef]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xie, L.; Wang, M.; Xiong, Q.; Guo, Y.; Liang, Y.; Li, J.; Sheng, R.; Deng, P.; Wang, Y.; et al. Mettl3-mediated m(6)A RNA methylation regulates the fate of bone marrow mesenchymal stem cells and osteoporosis. Nat. Commun. 2018, 9, 4772. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, C.; Liang, W.; Kang, F.; Bai, Y.; Ma, B.; Wu, C.; Dong, S. Mn-containing bioceramics inhibit osteoclastogenesis and promote osteoporotic bone regeneration via scavenging ROS. Bioact. Mater. 2021, 6, 3839–3850. [Google Scholar] [CrossRef]
- Ma, Z.; Sun, J.; Jiang, Q.; Zhao, Y.; Jiang, H.; Sun, P.; Feng, W. Identification and analysis of mitochondria-related central genes in steroid-induced osteonecrosis of the femoral head, along with drug prediction. Front. Endocrinol. 2024, 15, 1341366. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, Y.D.; Kim, H.J.; Lee, Z.H.; Kim, H.H. SOD2 and Sirt3 Control Osteoclastogenesis by Regulating Mitochondrial ROS. J. Bone Min. Res. 2017, 32, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Zhang, L.Q.; Konermann, A.; Zhou, H.; Jin, F.; Liu, W.J. Manganese superoxide dismutase is required to maintain osteoclast differentiation and function under static force. Sci. Rep. 2015, 5, 8016. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Biological production, detection, and fate of hydrogen peroxide. Antioxid. Redox Signal. 2018, 29, 541–551. [Google Scholar] [CrossRef]
- Kanzaki, H.; Shinohara, F.; Kanako, I.; Yamaguchi, Y.; Fukaya, S.; Miyamoto, Y.; Wada, S.; Nakamura, Y. Molecular regulatory mechanisms of osteoclastogenesis through cytoprotective enzymes. Redox Biol. 2016, 8, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, J.; Wu, L.; Xu, Y.; Xiao, C.; Yang, X.; Sun, R.; Zhao, J.; Xu, J.; Liu, Q.; et al. CYT387, a JAK-Specific Inhibitor Impedes Osteoclast Activity and Oophorectomy-Induced Osteoporosis via Modulating RANKL and ROS Signaling Pathways. Front. Pharmacol. 2022, 13, 829862. [Google Scholar] [CrossRef]
- Chang, X.; Xu, S.; Zhang, H. Regulation of bone health through physical exercise: Mechanisms and types. Front. Endocrinol. 2022, 13, 1029475. [Google Scholar] [CrossRef]
- Xiong, J.; Piemontese, M.; Onal, M.; Campbell, J.; Goellner, J.J.; Dusevich, V.; Bonewald, L.; Manolagas, S.C.; O’Brien, C.A. Osteocytes, not Osteoblasts or Lining Cells, are the Main Source of the RANKL Required for Osteoclast Formation in Remodeling Bone. PLoS ONE 2015, 10, e0138189. [Google Scholar] [CrossRef]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef] [PubMed]
- Marcadet, L.; Bouredji, Z.; Argaw, A.; Frenette, J. The Roles of RANK/RANKL/OPG in Cardiac, Skeletal, and Smooth Muscles in Health and Disease. Front. Cell Dev. Biol. 2022, 10, 903657. [Google Scholar] [CrossRef]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Kishikawa, A.; Ogawa, S.; Shen, W.R.; Qi, J.; Noguchi, T.; Nara, Y.; Mizoguchi, I. TNF-alpha Directly Enhances Osteocyte RANKL Expression and Promotes Osteoclast Formation. Front. Immunol. 2019, 10, 2925. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, F.; Jiang, M.; Han, M.; Huang, B. Roles of osteoprotegerin in endocrine and metabolic disorders through receptor activator of nuclear factor kappa-B ligand/receptor activator of nuclear factor kappa-B signaling. Front. Cell Dev. Biol. 2022, 10, 1005681. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H. Discovery of the RANKL/RANK/OPG system. J. Bone Miner. Metab. 2021, 39, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Marques-Carvalho, A.; Kim, H.N.; Almeida, M. The role of reactive oxygen species in bone cell physiology and pathophysiology. Bone Rep. 2023, 19, 101664. [Google Scholar] [CrossRef]
- Arias, C.F.; Herrero, M.A.; Echeverri, L.F.; Oleaga, G.E.; Lopez, J.M. Bone remodeling: A tissue-level process emerging from cell-level molecular algorithms. PLoS ONE 2018, 13, e0204171. [Google Scholar] [CrossRef] [PubMed]
- Weivoda, M.M.; Chew, C.K.; Monroe, D.G.; Farr, J.N.; Atkinson, E.J.; Geske, J.R.; Eckhardt, B.; Thicke, B.; Ruan, M.; Tweed, A.J.; et al. Identification of osteoclast-osteoblast coupling factors in humans reveals links between bone and energy metabolism. Nat. Commun. 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Gallego, J.L.; Manchado-Morales, P.; Pivonka, P.; Martinez-Reina, J. Spatio-temporal simulations of bone remodelling using a bone cell population model based on cell availability. Front. Bioeng. Biotechnol. 2023, 11, 1060158. [Google Scholar] [CrossRef] [PubMed]
- Krasnova, O.; Neganova, I. Assembling the Puzzle Pieces. Insights for in Vitro Bone Remodeling. Stem. Cell Rev. Rep. 2023, 19, 1635–1658. [Google Scholar] [CrossRef] [PubMed]
- Lerner, U.H.; Kindstedt, E.; Lundberg, P. The critical interplay between bone resorbing and bone forming cells. J. Clin. Periodontol. 2019, 46 (Suppl. S21), 33–51. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhong, L.; Yao, L.; Wei, Y.; Gui, T.; Li, Z.; Kim, H.; Holdreith, N.; Jiang, X.; Tong, W.; et al. Bone marrow adipogenic lineage precursors promote osteoclastogenesis in bone remodeling and pathologic bone loss. J. Clin. Investig. 2021, 131, e140214. [Google Scholar] [CrossRef] [PubMed]
- da Silva Mello, A.S.; dos Santos, P.L.; Marquesi, A.; Queiroz, T.P.; Margonar, R.; de Souza Faloni, A.P. Some aspects of bone remodeling around dental implants. Rev. Clínica Periodoncia Implantol. Y Rehabil. Oral 2016, in press. [Google Scholar] [CrossRef]
- Qin, L.; He, T.; Yang, D.; Wang, Y.; Li, Z.; Yan, Q.; Zhang, P.; Chen, Z.; Lin, S.; Gao, H.; et al. Osteocyte beta1 integrin loss causes low bone mass and impairs bone mechanotransduction in mice. J. Orthop. Translat. 2022, 34, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.X.; Dong, H.; Yu, J.H.; Yan, Y.; Wu, X.G.; Wang, Y.Q.; Li, P.C.; Wei, X.C.; Chen, W.Y. Mechanosensation of osteocyte with collagen hillocks and primary cilia under pressure and electric field stimulation. Acta Mech. Sinica-Prc. 2022, 38, 621569. [Google Scholar] [CrossRef]
- Choi, J.U.A.; Kijas, A.W.; Lauko, J.; Rowan, A.E. The Mechanosensory Role of Osteocytes and Implications for Bone Health and Disease States. Front. Cell Dev. Biol. 2021, 9, 770143. [Google Scholar] [CrossRef]
- Zeng, Y.; Riquelme, M.A.; Hua, R.; Zhang, J.; Acosta, F.M.; Gu, S.; Jiang, J.X. Mechanosensitive piezo1 calcium channel activates connexin 43 hemichannels through PI3K signaling pathway in bone. Cell Biosci. 2022, 12, 191. [Google Scholar] [CrossRef]
- Wu, V.; Helder, M.N.; Bravenboer, N.; Ten Bruggenkate, C.M.; Jin, J.; Klein-Nulend, J.; Schulten, E. Bone Tissue Regeneration in the Oral and Maxillofacial Region: A Review on the Application of Stem Cells and New Strategies to Improve Vascularization. Stem. Cells Int. 2019, 2019, 6279721. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Wang, N.X.; Luo, Y.; Yu, C.Y.; Xiao, J.H. Ganoderal A effectively induces osteogenic differentiation of human amniotic mesenchymal stem cells via cross-talk between Wnt/beta-catenin and BMP/SMAD signaling pathways. Biomed. Pharmacother. 2020, 123, 109807. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yuan, X.; Arioka, M.; Bahat, D.; Sun, Q.; Chen, J.; Helms, J.A. Pro-osteogenic Effects of WNT in a Mouse Model of Bone Formation Around Femoral Implants. Calcif. Tissue Int. 2021, 108, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Ohkawara, B.; Ito, M.; Masuda, A.; Hirakawa, A.; Sakai, T.; Hiraiwa, H.; Hamada, T.; Ishiguro, N.; Ohno, K. Fluoxetine ameliorates cartilage degradation in osteoarthritis by inhibiting Wnt/beta-catenin signaling. PLoS ONE 2017, 12, e0184388. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Liu, C.F.; Zhang, F.; Dong, W.T.; Zhang, X.F.; Sang, Y.H.; Wang, J.J.; Guo, Y.G.; Liu, H.; Wang, S.H. Tunable Layered (Na,Mn)V8O20· n H2O Cathode Material for High-Performance Aqueous Zinc Ion Batteries. Adv. Sci. 2020, 7, 107937. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Yang, F.; Ling, M.; Fan, Y. Association between bone trace elements and osteoporosis in older adults: A cross-sectional study. Ther. Adv. Musculoskelet. Dis. 2022, 14, 1759720X221125984. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, J.; Zhang, D. Manganese Dioxide-Based Nanomaterials for Medical Applications. ACS Biomater. Sci. Eng. 2024, 10, 2680–2702. [Google Scholar] [CrossRef] [PubMed]
- Anish, R.J.; Mohanan, B.; Nair, A.; Radhakrishnan, K.V.; Rauf, A.A. Protective effect of Pterospermum rubiginosum bark extract on bone mineral density and bone remodelling in estrogen deficient ovariectomized Sprague-Dawley (SD) rats. 3 Biotech 2024, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- El-Ganzuri, M.A.; Ahmed, R.R.; Bastawy, E.M. Regulatory Mechanisms of Bone Development and Function. Ann. Cytol. Pathol. 2016, 1, 005–017. [Google Scholar] [CrossRef]
- Mercadante, C.J.; Prajapati, M.; Conboy, H.L.; Dash, M.E.; Herrera, C.; Pettiglio, M.A.; Cintron-Rivera, L.; Salesky, M.A.; Rao, D.B.; Bartnikas, T.B. Manganese transporter Slc30a10 controls physiological manganese excretion and toxicity. J. Clin. Investig. 2019, 129, 5442–5461. [Google Scholar] [CrossRef]
- Nakata, T.; Creasey, E.A.; Kadoki, M.; Lin, H.; Selig, M.K.; Yao, J.; Lefkovith, A.; Daly, M.J.; Graham, D.B.; Xavier, R.J. A missense variant in SLC39A8 confers risk for Crohn’s disease by disrupting manganese homeostasis and intestinal barrier integrity. Proc. Natl. Acad. Sci. USA 2020, 117, 28930–28938. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.P.; Browning, C.L.; Vaslet, C.A.; Gion, K.D.; Green, A.; Liu, M.; Kane, A.B.; Hurt, R.H. Chemical and Colloidal Dynamics of MnO(2) Nanosheets in Biological Media Relevant for Nanosafety Assessment. Small 2020, 16, e2000303. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P. Invited review: Mineral absorption mechanisms, mineral interactions that affect acid-base and antioxidant status, and diet considerations to improve mineral status. J. Dairy Sci. 2018, 101, 2763–2813. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.J.; Hein, J.; Baez, A.; Sosa, J.C.; Wessling-Resnick, M. Manganese transport and toxicity in polarized WIF-B hepatocytes. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G351–G363. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Park, J.E.; Gugnani, K.; Betharia, S.; Pino-Figueroa, A.; Kim, J. Influence of iron metabolism on manganese transport and toxicity. Metallomics 2017, 9, 1028–1046. [Google Scholar] [CrossRef] [PubMed]
- Wooten, A.L.; Aweda, T.A.; Lewis, B.C.; Gross, R.B.; Lapi, S.E. Biodistribution and PET Imaging of pharmacokinetics of manganese in mice using Manganese-52. PLoS ONE 2017, 12, e0174351. [Google Scholar] [CrossRef] [PubMed]
- Peres, T.V.; Schettinger, M.R.; Chen, P.; Carvalho, F.; Avila, D.S.; Bowman, A.B.; Aschner, M. Manganese-induced neurotoxicity: A review of its behavioral consequences and neuroprotective strategies. BMC Pharmacol. Toxicol. 2016, 17, 57. [Google Scholar] [CrossRef]
- Liu, Q.; Barker, S.; Knutson, M.D. Iron and manganese transport in mammalian systems. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118890. [Google Scholar] [CrossRef]
- Aschner, M.; Costa, L.G. Linking Environmental Exposure to Neurodevelopmental Disorders; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Wolff, N.A.; Garrick, M.D.; Zhao, L.; Garrick, L.M.; Ghio, A.J.; Thevenod, F. A role for divalent metal transporter (DMT1) in mitochondrial uptake of iron and manganese. Sci. Rep. 2018, 8, 211. [Google Scholar] [CrossRef]
- Yanatori, I.; Kishi, F. DMT1 and iron transport. Free Radic. Biol. Med. 2019, 133, 55–63. [Google Scholar] [CrossRef]
- Knutson, M.D. Non-transferrin-bound iron transporters. Free Radic. Biol. Med. 2019, 133, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Fu, X.; Wang, X.; Wu, Q.; Lu, Y.; Shi, J.; Klaunig, J.E.; Zhou, S. Autophagy plays a protective role in Mn-induced toxicity in PC12 cells. Toxicology 2018, 394, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Berry, S.P.; Wilson, E.A.; Zhang, C.H.; Shekhar, M.; Singharoy, A.; Gaudet, R. High-resolution structures with bound Mn(2+) and Cd(2+) map the metal import pathway in an Nramp transporter. Elife 2023, 12, e84006. [Google Scholar] [CrossRef]
- Shawki, A.; Anthony, S.R.; Nose, Y.; Engevik, M.A.; Niespodzany, E.J.; Barrientos, T.; Ohrvik, H.; Worrell, R.T.; Thiele, D.J.; Mackenzie, B. Intestinal DMT1 is critical for iron absorption in the mouse but is not required for the absorption of copper or manganese. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G635–G647. [Google Scholar] [CrossRef]
- Fujishiro, H.; Kambe, T. Manganese transport in mammals by zinc transporter family proteins, ZNT and ZIP. J. Pharmacol. Sci. 2022, 148, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Winslow, J.W.W.; Limesand, K.H.; Zhao, N. The Functions of ZIP8, ZIP14, and ZnT10 in the Regulation of Systemic Manganese Homeostasis. Int. J. Mol. Sci. 2020, 21, 3304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, Y.H.; Gao, K.F.; Sui, D.X.; Yu, P.X.; Su, M.; Wei, G.W.; Hu, J. Structural insights into the elevator-type transport mechanism of a bacterial ZIP metal transporter. Nat. Commun. 2023, 14, 385. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B.; Kim, M.H.; Kim, J.; Colon-Perez, L.M.; Banan, G.; Mareci, T.H.; Febo, M.; Cousins, R.J. Metal Transporter Zip14 (Slc39a14) Deletion in Mice Increases Manganese Deposition and Produces Neurotoxic Signatures and Diminished Motor Activity. J. Neurosci. 2017, 37, 5996–6006. [Google Scholar] [CrossRef]
- Nishito, Y.; Tsuji, N.; Fujishiro, H.; Takeda, T.A.; Yamazaki, T.; Teranishi, F.; Okazaki, F.; Matsunaga, A.; Tuschl, K.; Rao, R.; et al. Direct Comparison of Manganese Detoxification/Efflux Proteins and Molecular Characterization of ZnT10 Protein as a Manganese Transporter. J. Biol. Chem. 2016, 291, 14773–14787. [Google Scholar] [CrossRef]
- Zogzas, C.E.; Aschner, M.; Mukhopadhyay, S. Structural Elements in the Transmembrane and Cytoplasmic Domains of the Metal Transporter SLC30A10 Are Required for Its Manganese Efflux Activity. J. Biol. Chem. 2016, 291, 15940–15957. [Google Scholar] [CrossRef]
- Hutchens, S.; Liu, C.; Jursa, T.; Shawlot, W.; Chaffee, B.K.; Yin, W.; Gore, A.C.; Aschner, M.; Smith, D.R.; Mukhopadhyay, S. Deficiency in the manganese efflux transporter SLC30A10 induces severe hypothyroidism in mice. J. Biol. Chem. 2017, 292, 9760–9773. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Kočvara, M. Elemental content in the tissues of the song thrush Turdus philomelos I. Accumulation of macro-and microminerals in internal organs and tissues. Oecologia Mont. 2023, 32, 9–24. [Google Scholar]
- Prajapati, M.; Conboy, H.L.; Hojyo, S.; Fukada, T.; Budnik, B.; Bartnikas, T.B. Biliary excretion of excess iron in mice requires hepatocyte iron import by Slc39a14. J. Biol. Chem. 2021, 297, 100835. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Tsukamoto, M.; Saito, M.; Hojyo, S.; Fukada, T.; Takami, M.; Furuichi, T. Disruption of the mouse Slc39a14 gene encoding zinc transporter ZIP 14 is associated with decreased bone mass, likely caused by enhanced bone resorption. FEBS Open Bio. 2018, 8, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.C.; Jee, Y.H.; Garrison, P.; Iben, J.R.; Yue, S.; Ad, M.; Nguyen, Q.; Kikani, B.; Wakabayashi, Y.; Baron, J. Differential aging of growth plate cartilage underlies differences in bone length and thus helps determine skeletal proportions. PLoS Biol. 2018, 16, e2005263. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Zhang, Y.T.; Cao, C.Y.; Liu, J.M.; Deng, Y.; Zhang, Z.Q.; Wang, C. TaNRAMP3 is essential for manganese transport in. Stress Biol. 2023, 3, 41. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Yan, G.; Guan, M. Zinc Homeostasis in Bone: Zinc Transporters and Bone Diseases. Int. J. Mol. Sci. 2020, 21, 1236. [Google Scholar] [CrossRef] [PubMed]
- Felber, D.M.; Wu, Y.; Zhao, N. Regulation of the Metal Transporters ZIP14 and ZnT10 by Manganese Intake in Mice. Nutrients 2019, 11, 2099. [Google Scholar] [CrossRef]
- Takagishi, T.; Hara, T.; Fukada, T. Recent Advances in the Role of SLC39A/ZIP Zinc Transporters In Vivo. Int. J. Mol. Sci. 2017, 18, 2708. [Google Scholar] [CrossRef]
- Kong, N.; Zhao, Q.; Liu, C.; Li, J.; Liu, Z.; Gao, L.; Wang, L.; Song, L. The involvement of zinc transporters in the zinc accumulation in the Pacific oyster Crassostrea gigas. Gene 2020, 750, 144759. [Google Scholar] [CrossRef] [PubMed]
- Wiuf, A.; Steffen, J.H.; Becares, E.R.; Gronberg, C.; Mahato, D.R.; Rasmussen, S.G.F.; Andersson, M.; Croll, T.; Gotfryd, K.; Gourdon, P. The two-domain elevator-type mechanism of zinc-transporting ZIP proteins. Sci. Adv. 2022, 8, eabn4331. [Google Scholar] [CrossRef]
- Tang, H.; Li, C.; Zhang, Y.; Zheng, H.; Cheng, Y.; Zhu, J.; Chen, X.; Zhu, Z.; Piao, J.G.; Li, F. Targeted Manganese doped silica nano GSH-cleaner for treatment of Liver Cancer by destroying the intracellular redox homeostasis. Theranostics 2020, 10, 9865–9887. [Google Scholar] [CrossRef]
- Lin, W.; Vann, D.R.; Doulias, P.T.; Wang, T.; Landesberg, G.; Li, X.; Ricciotti, E.; Scalia, R.; He, M.; Hand, N.J.; et al. Hepatic metal ion transporter ZIP8 regulates manganese homeostasis and manganese-dependent enzyme activity. J. Clin. Investig. 2017, 127, 2407–2417. [Google Scholar] [CrossRef] [PubMed]
- Coffey, R.; Knutson, M.D. The plasma membrane metal-ion transporter ZIP14 contributes to nontransferrin-bound iron uptake by human beta-cells. Am. J. Physiol. Cell Physiol. 2017, 312, C169–C175. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Liu, Z. SLC39A8 gene encoding a metal ion transporter: Discovery and bench to bedside. Hum. Genom. 2019, 13, 51. [Google Scholar] [CrossRef]
- Kim, G.; Elnabawi, O.; Shin, D.; Pae, E.K. Transient Intermittent Hypoxia Exposure Disrupts Neonatal Bone Strength. Front. Pediatr. 2016, 4, 15. [Google Scholar] [CrossRef]
- Bin, B.H.; Bhin, J.; Seo, J.; Kim, S.Y.; Lee, E.; Park, K.; Choi, D.H.; Takagishi, T.; Hara, T.; Hwang, D.; et al. Requirement of Zinc Transporter SLC39A7/ZIP7 for Dermal Development to Fine-Tune Endoplasmic Reticulum Function by Regulating Protein Disulfide Isomerase. J. Investig. Dermatol. 2017, 137, 1682–1691. [Google Scholar] [CrossRef]
- Fang, L.; Watkinson, M. Subcellular localised small molecule fluorescent probes to image mobile Zn(2). Chem. Sci. 2020, 11, 11366–11379. [Google Scholar] [CrossRef]
- Riley, L.G.; Cowley, M.J.; Gayevskiy, V.; Roscioli, T.; Thorburn, D.R.; Prelog, K.; Bahlo, M.; Sue, C.M.; Balasubramaniam, S.; Christodoulou, J. A SLC39A8 variant causes manganese deficiency, and glycosylation and mitochondrial disorders. J. Inherit. Metab. Dis. 2017, 40, 261–269. [Google Scholar] [CrossRef]
- Geng, X.; Liu, L.; Banes-Berceli, A.; Yang, Z.; Kang, P.; Shen, J.; Tsai, K.J.; Liu, Z. Role of ZIP8 in regulating cell morphology and NF-kappaB/Snail2 signaling. Metallomics 2018, 10, 953–964. [Google Scholar] [CrossRef]
- Conley, T.E.; Richardson, C.; Pacheco, J.; Dave, N.; Jursa, T.; Guazzetti, S.; Lucchini, R.G.; Fendorf, S.; Ritchie, R.O.; Smith, D.R. Bone manganese is a sensitive biomarker of ongoing elevated manganese exposure, but does not accumulate across the lifespan. Environ. Res. 2022, 204, 112355. [Google Scholar] [CrossRef]
- Zeglam, A.; Abugrara, A.; Kabuka, M. Autosomal-recessive iron deficiency anemia, dystonia and hypermanganesemia caused by new variant mutation of the manganese transporter gene SLC39A14. Acta Neurol. Belg. 2019, 119, 379–384. [Google Scholar] [CrossRef]
- Morgan, S.E.; Schroten, H.; Ishikawa, H.; Zhao, N. Localization of ZIP14 and ZIP8 in HIBCPP Cells. Brain Sci. 2020, 10, 534. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, C.Q.; Wang, X.Y.; Yuan, Z.C.; Sun, B.B.; EL-Newehy, M.; Abdulhameed, M.M.; Fang, B.; Mo, X.M. Aspirin-Loaded Anti-Inflammatory ZnO-SiO2 Aerogel Scaffolds for Bone Regeneration. ACS Appl. Mater. Interfaces 2024, 16, 17092–17108. [Google Scholar] [CrossRef]
- Hendrickx, G.; Borra, V.M.; Steenackers, E.; Yorgan, T.A.; Hermans, C.; Boudin, E.; Waterval, J.J.; Jansen, I.D.C.; Aydemir, T.B.; Kamerling, N.; et al. Conditional mouse models support the role of SLC39A14 (ZIP14) in Hyperostosis Cranialis Interna and in bone homeostasis. PLoS Genet. 2018, 14, e1007321. [Google Scholar] [CrossRef]
- Das, B.K.; Wang, L.; Fujiwara, T.; Zhou, J.; Aykin-Burns, N.; Krager, K.J.; Lan, R.; Mackintosh, S.G.; Edmondson, R.; Jennings, M.L.; et al. Transferrin receptor 1-mediated iron uptake regulates bone mass in mice via osteoclast mitochondria and cytoskeleton. Elife 2022, 11, e73539. [Google Scholar] [CrossRef]
- Torres, P.M.C.; Vieira, S.I.; Cerqueira, A.R.; Pina, S.; Silva, O.A.B.D.; Abrantes, J.C.C.; Ferreira, J.M.F. Effects of Mn-doping on the structure and biological properties of β-tricalcium phosphate. J. Inorg. Biochem. 2014, 136, 57–66. [Google Scholar] [CrossRef]
- Wu, T.; Shi, H.; Liang, Y.; Lu, T.; Lin, Z.; Ye, J. Improving osteogenesis of calcium phosphate bone cement by incorporating with manganese doped beta-tricalcium phosphate. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 109, 110481. [Google Scholar] [CrossRef]
- Hreha, J.; Wey, A.; Cunningham, C.; Krell, E.S.; Brietbart, E.A.; Paglia, D.N.; Montemurro, N.J.; Nguyen, D.A.; Lee, Y.J.; Komlos, D.; et al. Local manganese chloride treatment accelerates fracture healing in a rat model. J. Orthop. Res. 2015, 33, 122–130. [Google Scholar] [CrossRef]
- Zhao, Q.M.; Sun, Y.Y.; Wu, C.S.; Yang, J.; Bao, G.F.; Cui, Z.M. Enhanced osteogenic activity and antibacterial ability of manganese-titanium dioxide microporous coating on titanium surfaces. Nanotoxicology 2020, 14, 289–309. [Google Scholar] [CrossRef]
- Bourebaba, L.; Michalak, I.; Baouche, M.; Kucharczyk, K.; Fal, A.M.; Marycz, K. Cladophora glomerata enriched by biosorption with Mn(II) ions alleviates lipopolysaccharide-induced osteomyelitis-like model in MC3T3-E1, and 4B12 osteoclastogenesis. J. Cell Mol. Med. 2020, 24, 7282–7300. [Google Scholar] [CrossRef]
- Chen, Q.X.; Li, J.Y.; Han, F.; Meng, Q.C.; Wang, H.; Qiang, W.; Li, Z.X.; Li, F.F.; Xie, E.; Qin, X.Y.; et al. A Multifunctional Composite Hydrogel That Rescues the ROS Microenvironment and Guides the Immune Response for Repair of Osteoporotic Bone Defects. Adv. Funct. Mater. 2022, 32, 2201067. [Google Scholar] [CrossRef]
- Pepa, G.D.; Brandi, M.L. Microelements for bone boost: The last but not the least. Clin. Cases Miner. Bone Metab. 2016, 13, 181–185. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.Y.; Wang, Z.J.; Liu, R.; Liu, S.Q.; Wang, L. Effects of manganese deficiency on chondrocyte development in tibia growth plate of Arbor Acres chicks. J. Bone Miner. Metab. 2015, 33, 23–29. [Google Scholar] [CrossRef]
- Liu, R.; Jin, C.; Wang, Z.; Wang, Z.; Wang, J.; Wang, L. Effects of manganese deficiency on the microstructure of proximal tibia and OPG/RANKL gene expression in chicks. Vet. Res. Commun. 2015, 39, 31–37. [Google Scholar] [CrossRef]
- Wang, C.Y.; Xia, W.H.; Wang, L.; Wang, Z.Y. Manganese deficiency induces avian tibial dyschondroplasia by inhibiting chondrocyte proliferation and differentiation. Res. Vet. Sci. 2021, 140, 164–170. [Google Scholar] [CrossRef]
- Zhaojun, W.; Lin, W.; Zhenyong, W.; Jian, W.; Ran, L. Effects of manganese deficiency on serum hormones and biochemical markers of bone metabolism in chicks. J. Bone Miner. Metab. 2013, 31, 285–292. [Google Scholar] [CrossRef]
- Lu, L.; Jin, C.; Dong, P.F.; Wang, Z.Y. HIF-1alpha upregulation exerts the antagonistic effect against angiogenesis inhibition in manganese deficiency-induced tibial dyschondroplasia of broiler chicks. Vet. Res. Commun. 2022, 46, 1023–1032. [Google Scholar] [CrossRef]
- O’Neal, S.L.; Hong, L.; Fu, S.; Jiang, W.; Jones, A.; Nie, L.H.; Zheng, W. Manganese accumulation in bone following chronic exposure in rats: Steady-state concentration and half-life in bone. Toxicol. Lett. 2014, 229, 93–100. [Google Scholar] [CrossRef]
- Rolle-McFarland, D.; Liu, Y.; Zhou, J.; Mostafaei, F.; Zhou, Y.; Li, Y.; Fan, Q.; Zheng, W.; Nie, L.H.; Wells, E.M. Development of a Cumulative Exposure Index (CEI) for Manganese and Comparison with Bone Manganese and Other Biomarkers of Manganese Exposure. Int. J. Environ. Res. Public Health 2018, 15, 1341. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ge, X.; Liu, Z.; Huang, L.; Zhou, Y.; Liu, P.; Qin, L.; Lin, S.; Liu, C.; Hou, Q.; et al. Association between long-term occupational manganese exposure and bone quality among retired workers. Environ. Sci. Pollut. Res. Int. 2020, 27, 482–489. [Google Scholar] [CrossRef]
- Liu, J.; Tang, Y.; Chen, Y.; Zhang, X.; Xia, Y.; Geng, B. Association between blood manganese and bone mineral density in US adolescents. Environ. Sci. Pollut. Res. Int. 2023, 30, 29743–29754. [Google Scholar] [CrossRef]
- Salami, A. Manganese Chloride Attenuates Osteoporosis in Rats with Experimental Ulcerative Colitis. Arch. Basic Appl. Med. 2019, 7, 21–34. [Google Scholar]
- Yang, X.; Xiong, S.; Zhou, J.; Zhang, Y.; He, H.; Chen, P.; Li, C.; Wang, Q.; Shao, Z.; Wang, L. Coating of manganese functional polyetheretherketone implants for osseous interface integration. Front. Bioeng. Biotech. 2023, 11, 1182187. [Google Scholar] [CrossRef]
- Austin, C.; Richardson, C.; Smith, D.; Arora, M. Tooth manganese as a biomarker of exposure and body burden in rats. Environ. Res. 2017, 155, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ding, T.; Xue, Y.; Sun, J. Osteoinduction and long-term osseointegration promoted by combined effects of nitrogen and manganese elements in high nitrogen nickel-free stainless steel. J. Mater. Chem. B 2016, 4, 801–812. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.; Li, S.; Hou, Y.; Zhang, H. The effects of Mn(2+) on the proliferation, osteogenic differentiation and adipogenic differentiation of primary mouse bone marrow stromal cells. Biol. Trace Elem. Res. 2013, 151, 415–423. [Google Scholar] [CrossRef]
- Chen, X.; Yang, G.; Zhang, B.; Li, F.; Liu, L.; Li, F. Effects of manganese-supplemented diets on growth performance, blood biochemistry, nitrogen metabolism and skeletal development of rex rabbits. J. Trace Elem. Med. Biol. 2020, 61, 126543. [Google Scholar] [CrossRef]
- Santos, L.F.J.; Goncalves, A.M.; Neira, L.M.; Nakagi, V.S.; Macari, M.; Laurentiz, A.C.; Pizauro, J.M. Effects of Supplementation of Zinc, Manganese, or Copper and Different Phytase Levels in Serum and Bone Acid and Alkaline Phosphatases of Broiler Chicks. Braz. J. Poult. Sci 2023, 25, eRBCA-2022-1722. [Google Scholar] [CrossRef]
- Wang, Y.; Gou, Z.; Lin, X.; Fan, Q.; Ye, J.; Jiang, S. Optimal Level of Supplemental Manganese for Yellow-Feathered Broilers during the Growth Phase. Animals 2021, 11, 1389. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.H.; Tang, L.; Wang, Z.Y.; Wang, L. Effects of Inorganic and Organic Manganese Supplementation on Growth Performance, Tibia Development, and Oxidative Stress in Broiler Chickens. Biol. Trace Elem. Res. 2022, 200, 4453–4464. [Google Scholar] [CrossRef] [PubMed]
- Viegas, M.N.; Salgado, M.A.; Aguiar, C.; Almeida, A.; Gavaia, P.; Dias, J. Effect of Dietary Manganese and Zinc Levels on Growth and Bone Status of Senegalese Sole (Solea senegalensis) Post-Larvae. Biol. Trace Elem. Res. 2021, 199, 2012–2021. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Zhang, N.N.; Gong, Y.J.; Zhou, M.Y.; Zhan, H.Q.; Zou, X.T. Effects of dietary Mn-methionine supplementation on the egg quality of laying hens. Poult. Sci. 2018, 97, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Venglovska, K.; Grešáková, Ľ.; Placha, I.; Ryzner, M.; Čobanová, K. Effects of feed supplementation with manganese from its different sources on performance and egg parameters of laying hens Original Paper. Czech J. Anim. Sci. 2014, 59, 147–155. [Google Scholar] [CrossRef]
- Sun, Y.; Geng, S.; Yuan, T.; Liu, Y.; Zhang, Y.; Di, Y.; Li, J.; Zhang, L. Effects of Manganese Hydroxychloride on Growth Performance, Antioxidant Capacity, Tibia Parameters and Manganese Deposition of Broilers. Animals 2021, 11, 3470. [Google Scholar] [CrossRef]
- Cappelli, J.; Garcia, A.; Ceacero, F.; Gomez, S.; Luna, S.; Gallego, L.; Gambin, P.; Landete-Castillejos, T. Manganese Supplementation in Deer under Balanced Diet Increases Impact Energy and Contents in Minerals of Antler Bone Tissue. PLoS ONE 2015, 10, e0132738. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mn Compound Type | Bone Cell Types | Mechanisms of Action | Outcomes | References |
|---|---|---|---|---|
| Mn compound | Osteoblasts | Proliferation | Evaluated cell viability and type I collagen secretion | [122] |
| Mn compound | mBMSCs | Differentiation | Effective for osteogenic differentiation | [123] |
| MnCl2 | Osteoblasts | Osteogenesis | Accelerated osteogenesis, increased angiogenesis | [124] |
| Mn containing BG | hMSCs | Differentiation | High expression of osteogenic markers (ALP, collagen type I, osteopontin, osteocalcin) | [33] |
| Mn-containing coatings by micro-arc oxidation and PIIID | Extracellular matrix | Collagen secretion and mineralization | Improve bone differentiation | [125] |
| MnTBAP | BMMs | Downregulate osteoclast marker genes | Inhibit induction of NFATc1 | [40] |
| Mn2+ ions released from Mn-TCP bioceramics | Osteoclasts | Inhibit osteoclast formation | Decreased osteoclasts, accelerated bone defect regeneration by activating Nrf2 and scavenging ROS | [38] |
| Mn(II)-enriched C. glomerata methanolic extract | LPS-induced MC3T3-E1 cells | mRNA expression of osteoclast-related genes | Attenuate signaling pathways, reduce aberrant osteoclast differentiation | [126] |
| Mn2+ in composite hydrogels | Osteoclasts | Reduce ROS, inhibit NF-κB signaling | Suppress osteoclastogenesis, inhibit osteoclast formation | [127] |
| Mn Level (mg/kg) | Experimental Subjects | Tissue | Effects of Deficiency | References |
|---|---|---|---|---|
| Low diet | Rats | Bone | Inhibits cartilage formation and induces osteopenia | [128] |
| 40 mg/kg diet | Chickens | Bone | Inhibits chondrocyte proliferation and stimulates chondrocyte apoptosis | [129] |
| 8.7 mg/kg | Chickens | Bone | Leads to metaphyseal osteoporosis due to decreased OPG/RANKL mRNA expression | [130] |
| 22 mg/kg diet | Chickens | Tibia | Affects chondrocyte proliferation and differentiation in the tibial growth plate | [131] |
| 8.7 mg/kg | Chickens | Serum markers of bone | Causes disorders in bone regulatory hormones and enzymes of bone metabolism in serum | [132] |
| 22 mg/kg | Chickens | Tibial growth plate | HIF-1α up-regulation and autophagy activation protect against Mn deficiency-induced angiogenesis inhibition | [133] |
| Types of Supplementations | Species | Outcomes | Target Tissue | Mechanisms | References |
|---|---|---|---|---|---|
| Oral | Rex rabbits | Improved growth performance | Skeletal development | Significant effects on bone strength | [143] |
| Oral | Cobb broilers | Increased growth plate activity, accelerated calcification | Bone tissue | Increased growth plate activity, accelerated calcification | [144] |
| Oral | Yellow feather broilers | Increased bone density | Tibia characteristics | Increased tibia diameter | [145] |
| Organic Mn supplementation | Broilers | Improved leg development and absorption efficiency | Tibial bone | Increased growth performance, tibial bone parameters, oxidative stress indicators | [146] |
| Oral | Marine fish larvae | Increased bone Mn deposition | Bone | Reduced severity of vertebral defects | [147] |
| Oral, Mn-methionine supplementation | Laying hens | Increased eggshell ultrastructure | Eggshell | Affected egg quality | [148] |
| Oral, Mn supplementation from its inorganic and organic sources | Laying hens | Improved eggshell quality | Eggshell | Beneficial impact on eggshell quality | [149] |
| Dietary supplementation with Mn hydroxychloride | Arbor Acres broilers | Increased tibia length, strength, and density index | Tibial bone | Improved antioxidant capacity, bone quality, Mn deposition | [150] |
| Mn injection | Deer | Increased impact energy and mineral content in antler bone tissue | Bone tissue | Altered mineral composition, improved structure and mechanical properties | [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taskozhina, G.; Batyrova, G.; Umarova, G.; Issanguzhina, Z.; Kereyeva, N. The Manganese–Bone Connection: Investigating the Role of Manganese in Bone Health. J. Clin. Med. 2024, 13, 4679. https://doi.org/10.3390/jcm13164679
Taskozhina G, Batyrova G, Umarova G, Issanguzhina Z, Kereyeva N. The Manganese–Bone Connection: Investigating the Role of Manganese in Bone Health. Journal of Clinical Medicine. 2024; 13(16):4679. https://doi.org/10.3390/jcm13164679
Chicago/Turabian StyleTaskozhina, Gulaim, Gulnara Batyrova, Gulmira Umarova, Zhamilya Issanguzhina, and Nurgul Kereyeva. 2024. "The Manganese–Bone Connection: Investigating the Role of Manganese in Bone Health" Journal of Clinical Medicine 13, no. 16: 4679. https://doi.org/10.3390/jcm13164679