
COVID-19 and Pregnancy: A Dangerous Mix for Bone Turnover and Metabolism Biomarkers in Placenta and Colostrum

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Placenta Sampling
2.3. Multi-Elemental Analysis by Inductively Coupled Plasma-Mass Spectrometry (ICP-MS)
2.4. Colostrum Sampling
2.5. Energy and Bone Turnover Parameters
2.6. Statistical Analysis
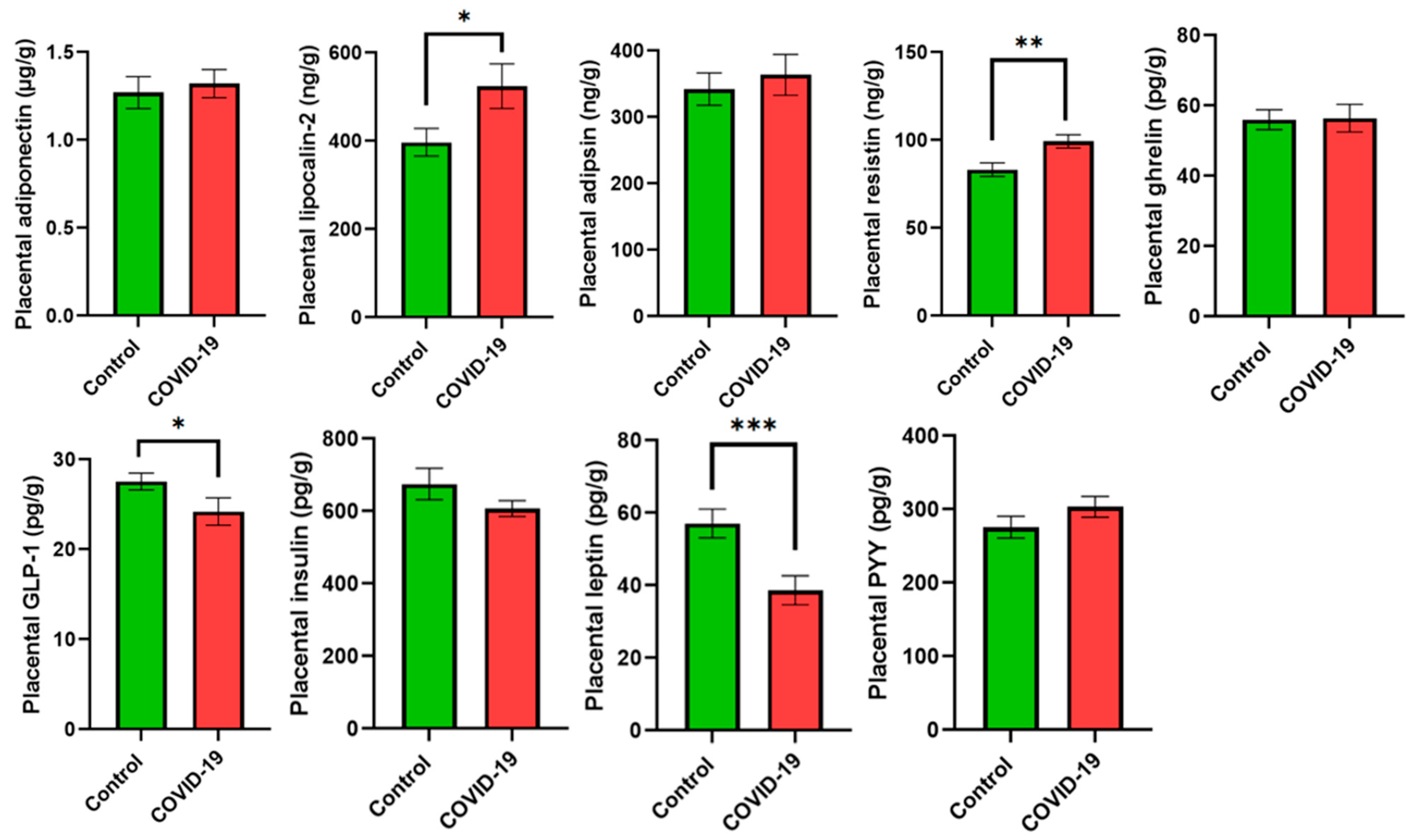
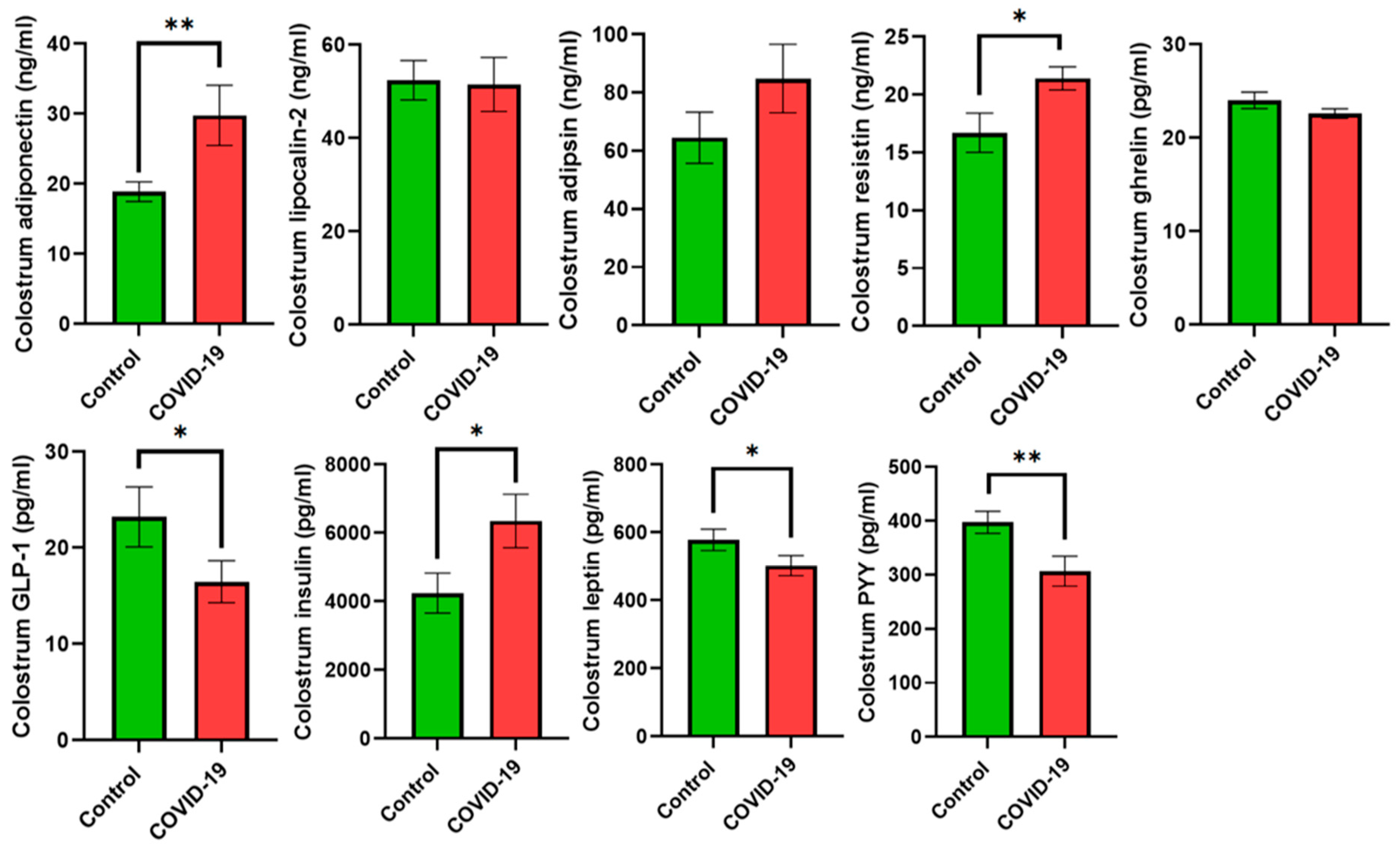
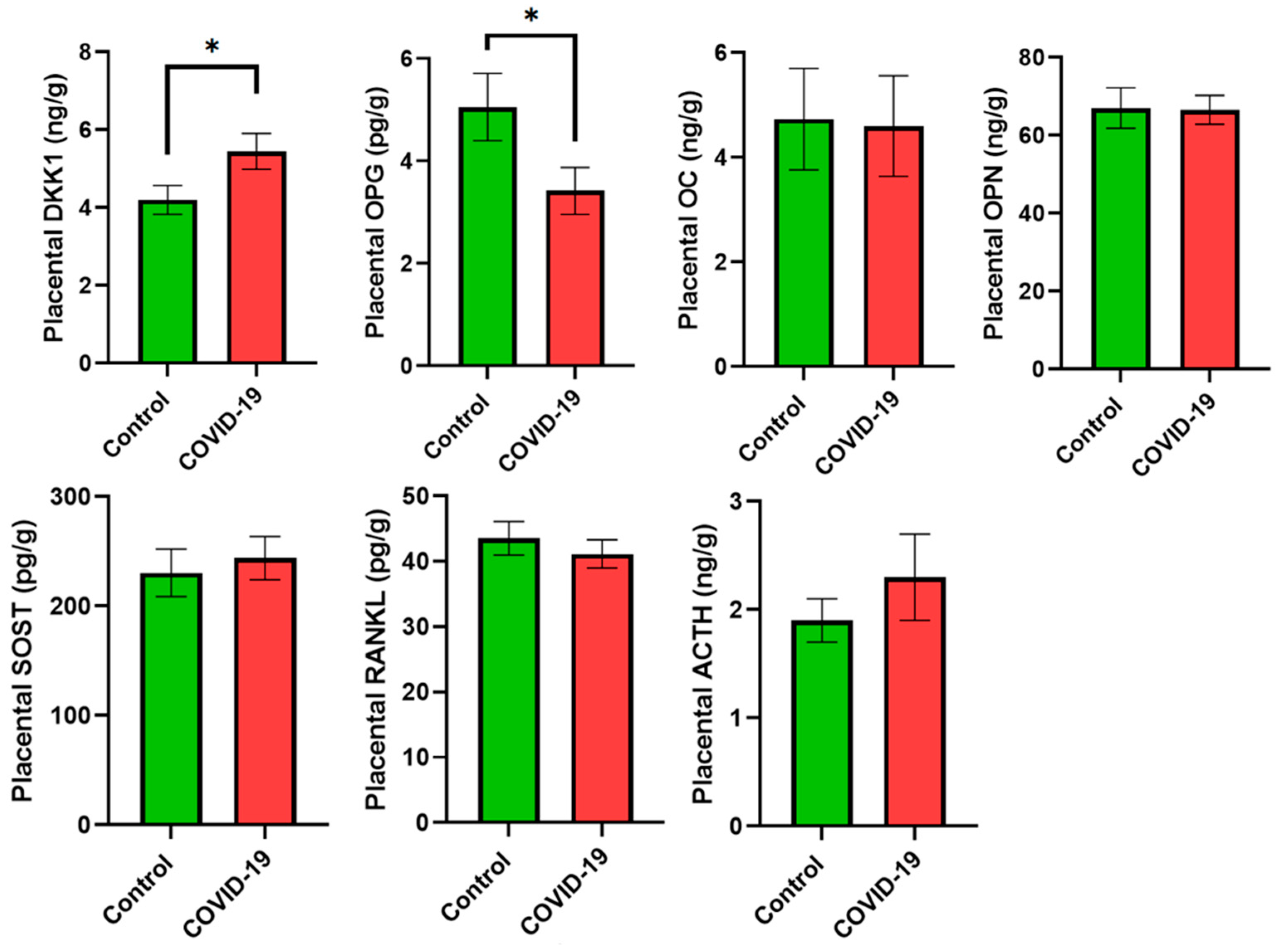
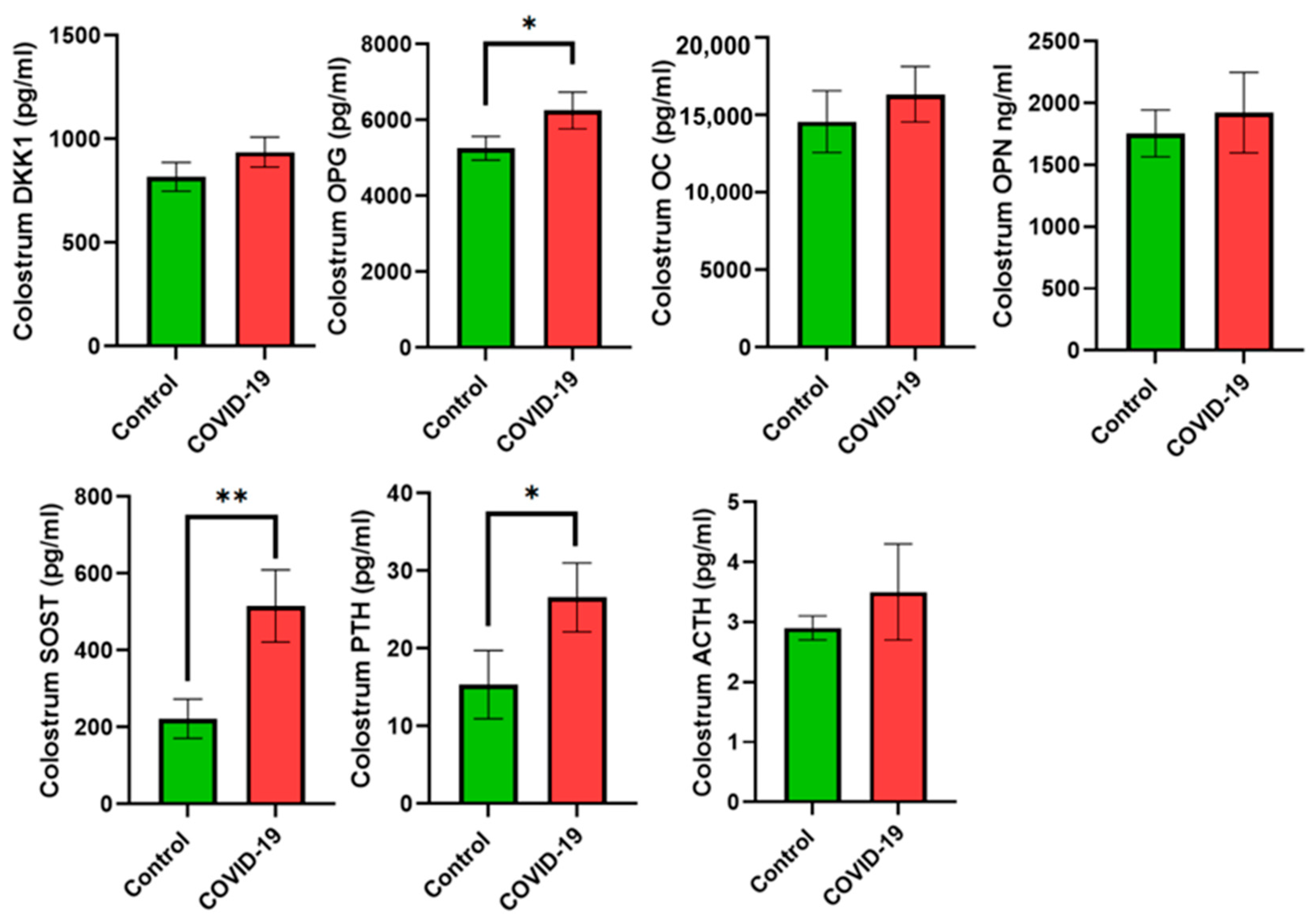
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Ander, S.E.; Diamond, M.S.; Coyne, C.B. Immune responses at the maternal-fetal interface. Sci. Immunol. 2019, 4, eaat6114. [Google Scholar] [CrossRef]
- Salles, J.P. Bone metabolism during pregnancy. Ann. Endocrinol. 2016, 77, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, P.; Quinello, C.; Silveira-Lessa, A.L.; Zago, C.A.; Carneiro-Sampaio, M. IgG placental transfer in healthy and pathological pregnancies. Clin. Dev. Immunol. 2012, 2012, 985646. [Google Scholar] [CrossRef] [PubMed]
- Delorme-Axford, E.; Sadovsky, Y.; Coyne, C.B. The Placenta as a Barrier to Viral Infections. Annu. Rev. Virol. 2014, 1, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.R.; Bakardjiev, A.I. Pathogens and the placental fortress. Curr. Opin. Microbiol. 2012, 15, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Delorme-Axford, E.; Sleigher, C.; Frey, T.K.; Trobaugh, D.W.; Klimstra, W.B.; Emert-Sedlak, L.A.; Smithgall, T.E.; Kinchington, P.R.; Vadia, S.; et al. Human trophoblasts confer resistance to viruses implicated in perinatal infection. Am. J. Obstet. Gynecol. 2015, 212, 71.e1–71.e8. [Google Scholar] [CrossRef]
- León-Juárez, M.; Martínez-Castillo, M.; González-García, L.D.; Helguera-Repetto, A.C.; Zaga-Clavellina, V.; García-Cordero, J.; Flores-Pliego, A.; Herrera-Salazar, A.; Vázquez-Martínez, E.R.; Reyes-Muñoz, E. Cellular and molecular mechanisms of viral infection in the human placenta. Pathog. Dis. 2017, 75, ftx093. [Google Scholar] [CrossRef]
- Blumberg, D.A.; Underwood, M.A.; Hedriana, H.L.; Lakshminrusimha, S. Vertical Transmission of SARS-CoV-2: What is the Optimal Definition? Am. J. Perinatol. 2020, 37, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Azinheira Nobrega Cruz, N.; Stoll, D.; Casarini, D.E.; Bertagnolli, M. Role of ACE2 in pregnancy and potential implications for COVID-19 susceptibility. Clin. Sci. 2021, 135, 1805–1824. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef]
- Indrio, F.; Neu, J.; Pettoello-Mantovani, M.; Marchese, F.; Martini, S.; Salatto, A.; Aceti, A. Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients 2022, 14, 1405. [Google Scholar] [CrossRef] [PubMed]
- Arias-Borrego, A.; Soto Cruz, F.J.; Selma-Royo, M.; Bäuerl, C.; García Verdevio, E.; Pérez-Cano, F.J.; Lerin, C.; Velasco López, I.; Martínez-Costa, C.; Collado, M.C.; et al. Metallomic and Untargeted Metabolomic Signatures of Human Milk from SARS-CoV-2 Positive Mothers. Mol. Nutr. Food Res. 2022, 66, e2200071. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Tan, M.; Zhu, J.; Tian, Y.; Liu, H.; Luo, F.; Wang, J.; Huang, Y.; Zhang, Y.; Yang, Y.; et al. Proteomic Analysis of Human Milk Reveals Nutritional and Immune Benefits in the Colostrum from Mothers with COVID-19. Nutrients 2022, 14, 2513. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Castro, J.; Mira-Rufino, P.J.; Moreno-Fernandez, J.; Chirosa, I.; Chirosa, J.L.; Guisado, R.; Ochoa, J.J. Ubiquinol supplementation modulates energy metabolism and bone turnover during high intensity exercise. Food Funct. 2020, 11, 7523–7531. [Google Scholar] [CrossRef] [PubMed]
- Kerschan-Schindl, K.; Dovjak, P.; Butylina, M.; Rainer, A.; Mayr, B.; Röggla, V.; Haslacher, H.; Weber, M.; Jordakieva, G.; Pietschmann, P. Moderate COVID-19 Disease Is Associated With Reduced Bone Turnover. J. Bone Miner. Res. 2023, 38, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cao, Y.; Zhang, H.; Wang, Z.; Man, C.H.; Yang, Y.; Chen, L.; Xu, S.; Yan, X.; Zheng, Q.; et al. COVID-19 metabolism: Mechanisms and therapeutic targets. MedComm 2022, 3, e157. [Google Scholar] [CrossRef]
- Kajarabille, N.; Hurtado, J.A.; Peña-Quintana, L.; Peña, M.; Ruiz, J.; Diaz-Castro, J.; Rodríguez-Santana, Y.; Martin-Alvarez, E.; López-Frias, M.; Soldado, O.; et al. Omega-3 LCPUFA supplement: A nutritional strategy to prevent maternal and neonatal oxidative stress. Matern. Child. Nutr. 2017, 13, e12300. [Google Scholar] [CrossRef]
- Jamieson, D.J.; Rasmussen, S.A. An update on COVID-19 and pregnancy. Am. J. Obstet. Gynecol. 2022, 226, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Al-Tamimi, A.O.; Halwani, R.; Alsaidi, H.; Kannan, M.; Ahmad, F. Lipocalin-2, S100A8/A9, and cystatin C: Potential predictive biomarkers of cardiovascular complications in COVID-19. Exp. Biol. Med. 2022, 247, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, H.; Bauer, C.; Hu, Y.; Lewis, J.R.; Xu, A.; Levinger, I.; Wang, Y. Lipocalin-2 Variants and Their Relationship with Cardio-Renal Risk Factors. Front. Endocrinol. 2021, 12, 781763. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Borregaard, N.; Kjeldsen, L.; Moses, M.A. The high molecular weight urinary matrix metalloproteinase (MMP) activity is a complex of gelatinase B/MMP-9 and neutrophil gelatinase-associated lipocalin (NGAL). Modulation of MMP-9 activity by NGAL. J. Biol. Chem. 2001, 276, 37258–37265. [Google Scholar] [CrossRef] [PubMed]
- Rajnics, P.; Kellner, Á.; Karádi, É.; Moizs, M.; Bödör, C.; Király, P.A.; Marosvári, D.; Andrikovics, H.; Egyed, M. Increased Lipocalin 2 level may have important role in thrombotic events in patients with polycythemia vera and essential thrombocythemia. Leuk. Res. 2016, 48, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Fernandez, J.; Ochoa, J.J.; De Paco Matallana, C.; Caño, A.; Martín-Alvarez, E.; Sanchez-Romero, J.; Toledano, J.M.; Puche-Juarez, M.; Prados, S.; Ruiz-Duran, S.; et al. COVID-19 during Gestation: Maternal Implications of Evoked Oxidative Stress and Iron Metabolism Impairment. Antioxidants 2022, 11, 184. [Google Scholar] [CrossRef]
- Birnhuber, A.; Fließer, E.; Gorkiewicz, G.; Zacharias, M.; Seeliger, B.; David, S.; Welte, T.; Schmidt, J.; Olschewski, H.; Wygrecka, M.; et al. Between inflammation and thrombosis: Endothelial cells in COVID-19. Eur. Respir. J. 2021, 58, 2100377. [Google Scholar] [CrossRef]
- Ebihara, T.; Matsumoto, H.; Matsubara, T.; Togami, Y.; Nakao, S.; Matsuura, H.; Onishi, S.; Kojima, T.; Sugihara, F.; Okuzaki, D.; et al. Resistin Associated With Cytokines and Endothelial Cell Adhesion Molecules Is Related to Worse Outcome in COVID-19. Front. Immunol. 2022, 13, 830061. [Google Scholar] [CrossRef] [PubMed]
- Leisman, D.E.; Ronner, L.; Pinotti, R.; Taylor, M.D.; Sinha, P.; Calfee, C.S.; Hirayama, A.V.; Mastroiani, F.; Turtle, C.J.; Harhay, M.O.; et al. Cytokine elevation in severe and critical COVID-19: A rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med. 2020, 8, 1233–1244. [Google Scholar] [CrossRef]
- Jiang, S.; Teague, A.M.; Tryggestad, J.B.; Lyons, T.J.; Chernausek, S.D. Fetal circulating human resistin increases in diabetes during pregnancy and impairs placental mitochondrial biogenesis. Mol. Med. 2020, 26, 76. [Google Scholar] [CrossRef]
- Jutant, E.M.; Tu, L.; Humbert, M.; Guignabert, C.; Huertas, A. The Thousand Faces of Leptin in the Lung. Chest 2021, 159, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, J.P.; Huang, H.; Wang, Z.H.; Cheng, R.; Cai, W.B. Leptin promotes fetal lung maturity and upregulates SP-A expression in pulmonary alveoli type-II epithelial cells involving TTF-1 activation. PLoS ONE 2013, 8, e69297. [Google Scholar] [CrossRef] [PubMed]
- Arteaga-Solis, E.; Zee, T.; Emala, C.W.; Vinson, C.; Wess, J.; Karsenty, G. Inhibition of leptin regulation of parasympathetic signaling as a cause of extreme body weight-associated asthma. Cell Metab. 2013, 17, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Ferrante, G.; Di Vincenzo, S.; Pace, E.; La Grutta, S. Leptin in the Respiratory Tract: Is There a Role in SARS-CoV-2 Infection? Front. Physiol. 2021, 12, 776963. [Google Scholar] [CrossRef] [PubMed]
- Abudalo, R.A.; Alqudah, A.M.; Roarty, C.; Athamneh, R.Y.; Grieve, D.J. Oxidative stress and inflammation in COVID-19: Potential application OF GLP-1 receptor agonists. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 6459–6471. [Google Scholar] [PubMed]
- Vikram, A.; Tripathi, D.N.; Kumar, A.; Singh, S. Oxidative stress and inflammation in diabetic complications. Int. J. Endocrinol. 2014, 2014, 679754. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.J.; Patterson, M.; Ghatei, M.A.; Bloom, S.R.; Wilson, C.A. Ghrelin and peptide YY (PYY) profiles in gastrointestinal tissues and the circulation of the rat during pregnancy and lactation. Peptides 2009, 30, 2213–2220. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, E.A.; Fondjo, L.A.; Owiredu, W.; Czika, A.; Nelson, W.; Lamptey, J.; Wang, Y.X.; Ding, Y.B. The role of adiponectin in placentation and preeclampsia. Cell Biochem. Funct. 2020, 38, 106–117. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.S.; Rulli, N.E.; Taylor, A.; Sheng, K.C.; Herrero, L.J.; Herring, B.L.; Lidbury, B.A.; Li, R.W.; Walsh, N.C.; et al. Arthritogenic alphaviral infection perturbs osteoblast function and triggers pathologic bone loss. Proc. Natl. Acad. Sci. USA 2014, 111, 6040–6045. [Google Scholar] [CrossRef]
- Harijith, A.; Ebenezer, D.L.; Natarajan, V. Reactive oxygen species at the crossroads of inflammasome and inflammation. Front. Physiol. 2014, 5, 352. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Koide, M.; Yamashita, T.; Nakamura, K.; Yasuda, H.; Udagawa, N.; Kobayashi, Y. Evidence for the major contribution of remodeling-based bone formation in sclerostin-deficient mice. Bone 2022, 160, 116401. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.; Levine, M.A. Hypocalcemia in the critically ill patient. J. Intensive Care Med. 2013, 28, 166–177. [Google Scholar] [CrossRef]
- He, L.; Ding, Y.; Zhang, Q.; Che, X.; He, Y.; Shen, H.; Wang, H.; Li, Z.; Zhao, L.; Geng, J.; et al. Expression of elevated levels of pro-inflammatory cytokines in SARS-CoV-infected ACE2+ cells in SARS patients: Relation to the acute lung injury and pathogenesis of SARS. J. Pathol. 2006, 210, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Gong, C.; Ma, S. Evaluation of COVID-19 based on ACE2 expression in normal and cancer patients. Open Med. 2020, 15, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Di Filippo, L.; Formenti, A.M.; Rovere-Querini, P.; Carlucci, M.; Conte, C.; Ciceri, F.; Zangrillo, A.; Giustina, A. Hypocalcemia is highly prevalent and predicts hospitalization in patients with COVID-19. Endocrine 2020, 68, 475–478. [Google Scholar] [CrossRef]
- Lafond, J.; Goyer-O’Reilly, I.; Laramée, M.; Simoneau, L. Hormonal regulation and implication of cell signaling in calcium transfer by placenta. Endocrine 2001, 14, 285–294. [Google Scholar] [CrossRef]
- Kamath, S.G.; Kelley, L.K.; Friedman, A.F.; Smith, C.H. Transport and binding in calcium uptake by microvillous membrane of human placenta. Am. J. Physiol.-Cell Physiol. 1992, 262, C789–C794. [Google Scholar] [CrossRef]
- Ke, H.Z.; Richards, W.G.; Li, X.; Ominsky, M.S. Sclerostin and Dickkopf-1 as therapeutic targets in bone diseases. Endocr. Rev. 2012, 33, 747–783. [Google Scholar] [CrossRef]
- Ueland, T.; Otterdal, K.; Lekva, T.; Halvorsen, B.; Gabrielsen, A.; Sandberg, W.J.; Paulsson-Berne, G.; Pedersen, T.M.; Folkersen, L.; Gullestad, L.; et al. Dickkopf-1 enhances inflammatory interaction between platelets and endothelial cells and shows increased expression in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1228–1234. [Google Scholar] [CrossRef]
- Astrup, E.; Lekva, T.; Davì, G.; Otterdal, K.; Santilli, F.; Oie, E.; Halvorsen, B.; Damås, J.K.; Raoult, D.; Vitale, G.; et al. A complex interaction between Rickettsia conorii and Dickkopf-1--potential role in immune evasion mechanisms in endothelial cells. PLoS ONE 2012, 7, e43638. [Google Scholar] [CrossRef]
- Pinzone, J.J.; Hall, B.M.; Thudi, N.K.; Vonau, M.; Qiang, Y.W.; Rosol, T.J.; Shaughnessy, J.D., Jr. The role of Dickkopf-1 in bone development, homeostasis, and disease. Blood 2009, 113, 517–525. [Google Scholar] [CrossRef]
- Christodoulides, C.; Laudes, M.; Cawthorn, W.P.; Schinner, S.; Soos, M.; O’Rahilly, S.; Sethi, J.K.; Vidal-Puig, A. The Wnt antagonist Dickkopf-1 and its receptors are coordinately regulated during early human adipogenesis. J. Cell Sci. 2006, 119, 2613–2620. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | COVID-19 | |
|---|---|---|
| Calcium (mg/g DM) | 2.01 ± 0.26 | 3.02 ± 0.51 * |
| Phosphorus (mg/g DM) | 9.34 ± 0.42 | 11.78 ± 0.63 ** |
| Magnesium (mg/g DM) | 0.32 ± 0.07 | 0.38 ± 0.11 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Castro, J.; Toledano, J.M.; Sanchez-Romero, J.; Aguilar, A.C.; Martín-Alvarez, E.; Puche-Juarez, M.; Moreno-Fernandez, J.; Pinar-Gonzalez, M.; Prados, S.; Carrillo, M.P.; et al. COVID-19 and Pregnancy: A Dangerous Mix for Bone Turnover and Metabolism Biomarkers in Placenta and Colostrum. J. Clin. Med. 2024, 13, 2124. https://doi.org/10.3390/jcm13072124
Diaz-Castro J, Toledano JM, Sanchez-Romero J, Aguilar AC, Martín-Alvarez E, Puche-Juarez M, Moreno-Fernandez J, Pinar-Gonzalez M, Prados S, Carrillo MP, et al. COVID-19 and Pregnancy: A Dangerous Mix for Bone Turnover and Metabolism Biomarkers in Placenta and Colostrum. Journal of Clinical Medicine. 2024; 13(7):2124. https://doi.org/10.3390/jcm13072124
Chicago/Turabian StyleDiaz-Castro, Javier, Juan M. Toledano, Javier Sanchez-Romero, Africa Caño Aguilar, Estefanía Martín-Alvarez, Maria Puche-Juarez, Jorge Moreno-Fernandez, Maria Pinar-Gonzalez, Sonia Prados, María Paz Carrillo, and et al. 2024. "COVID-19 and Pregnancy: A Dangerous Mix for Bone Turnover and Metabolism Biomarkers in Placenta and Colostrum" Journal of Clinical Medicine 13, no. 7: 2124. https://doi.org/10.3390/jcm13072124
APA StyleDiaz-Castro, J., Toledano, J. M., Sanchez-Romero, J., Aguilar, A. C., Martín-Alvarez, E., Puche-Juarez, M., Moreno-Fernandez, J., Pinar-Gonzalez, M., Prados, S., Carrillo, M. P., Ruiz-Duran, S., De Paco Matallana, C., & Ochoa, J. J. (2024). COVID-19 and Pregnancy: A Dangerous Mix for Bone Turnover and Metabolism Biomarkers in Placenta and Colostrum. Journal of Clinical Medicine, 13(7), 2124. https://doi.org/10.3390/jcm13072124