

Evolution of Minimally Invasive and Non-Invasive Preimplantation Genetic Testing: An Overview
 ,
, 
Abstract
1. Introduction
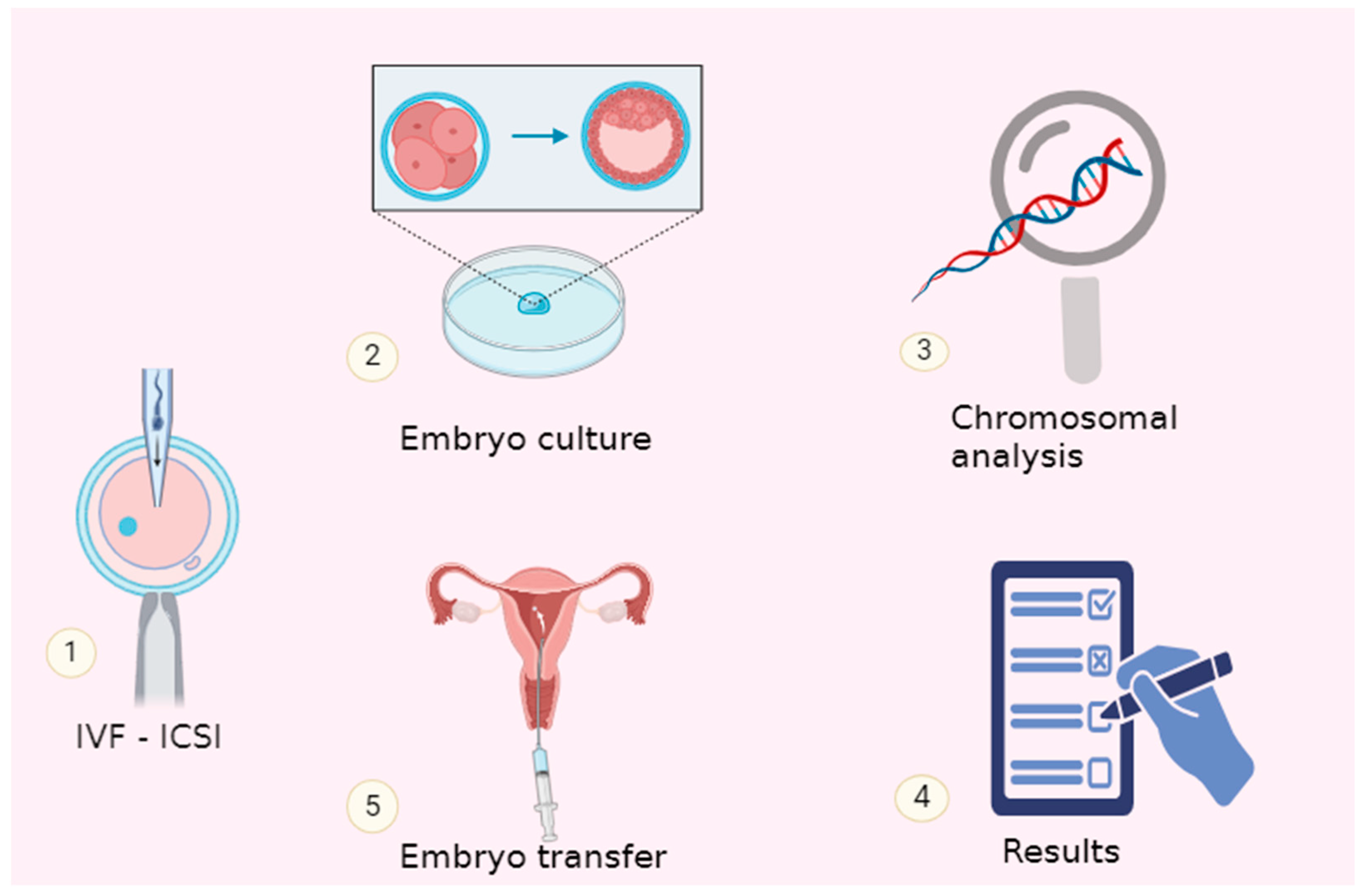
2. Schematic Evolution of PGT-A towards the Minimally Invasive and Non-Invasive Approaches
3. Standard Sources of Preimplantation Embryo Genetic Information
4. Origin of Embryonic DNA
5. Alternative Sources of Preimplantation Embryo Genetic Materialla
6. Recommendation of Trophectoderm Biopsy Replacement
7. Νon-Invasive Methods for Embryo Assessment
8. Automated Assessment
9. Benefits of Minimally Invasive and Non-Invasive PGT
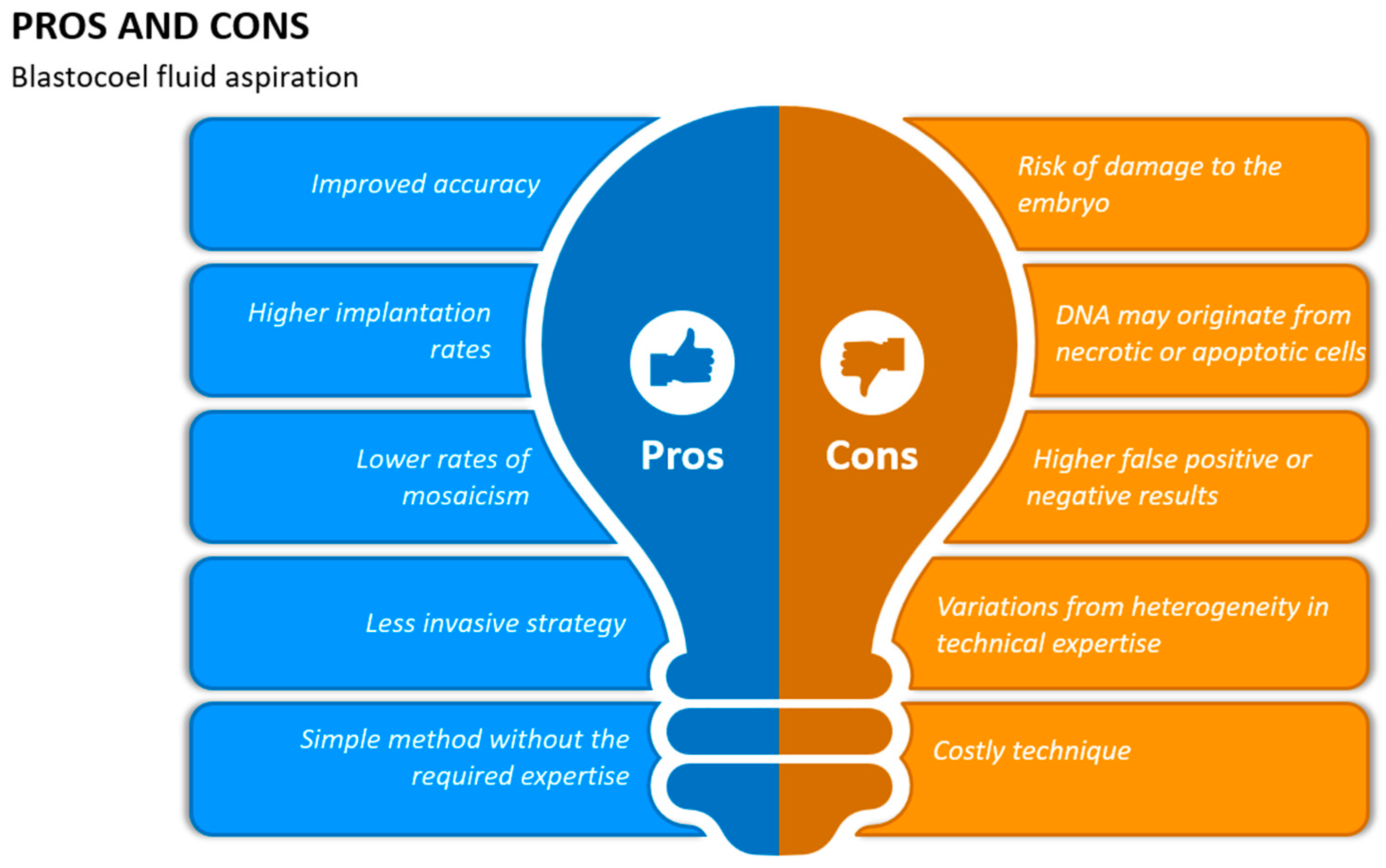
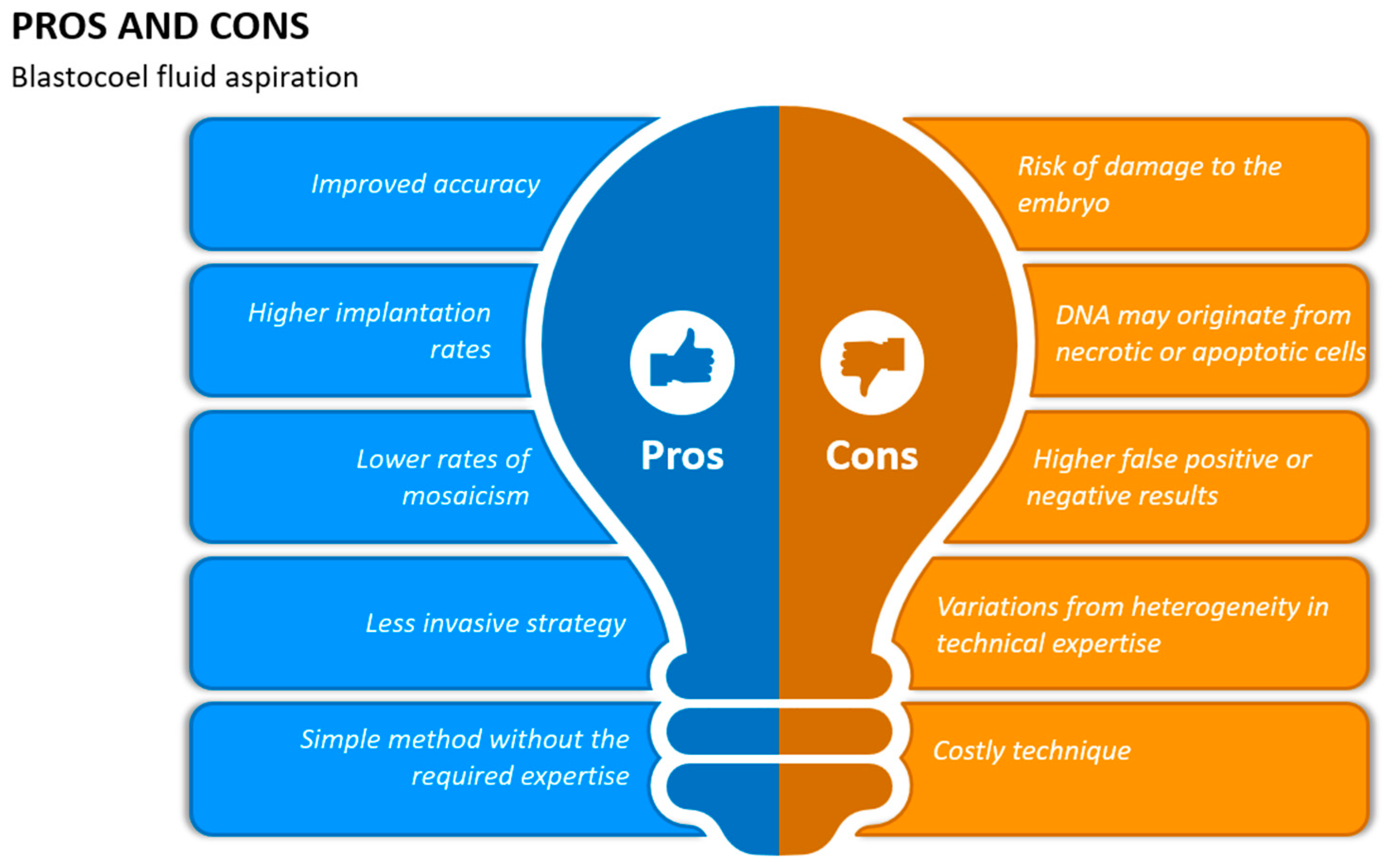
10. Blastocoel Fluid Aspiration
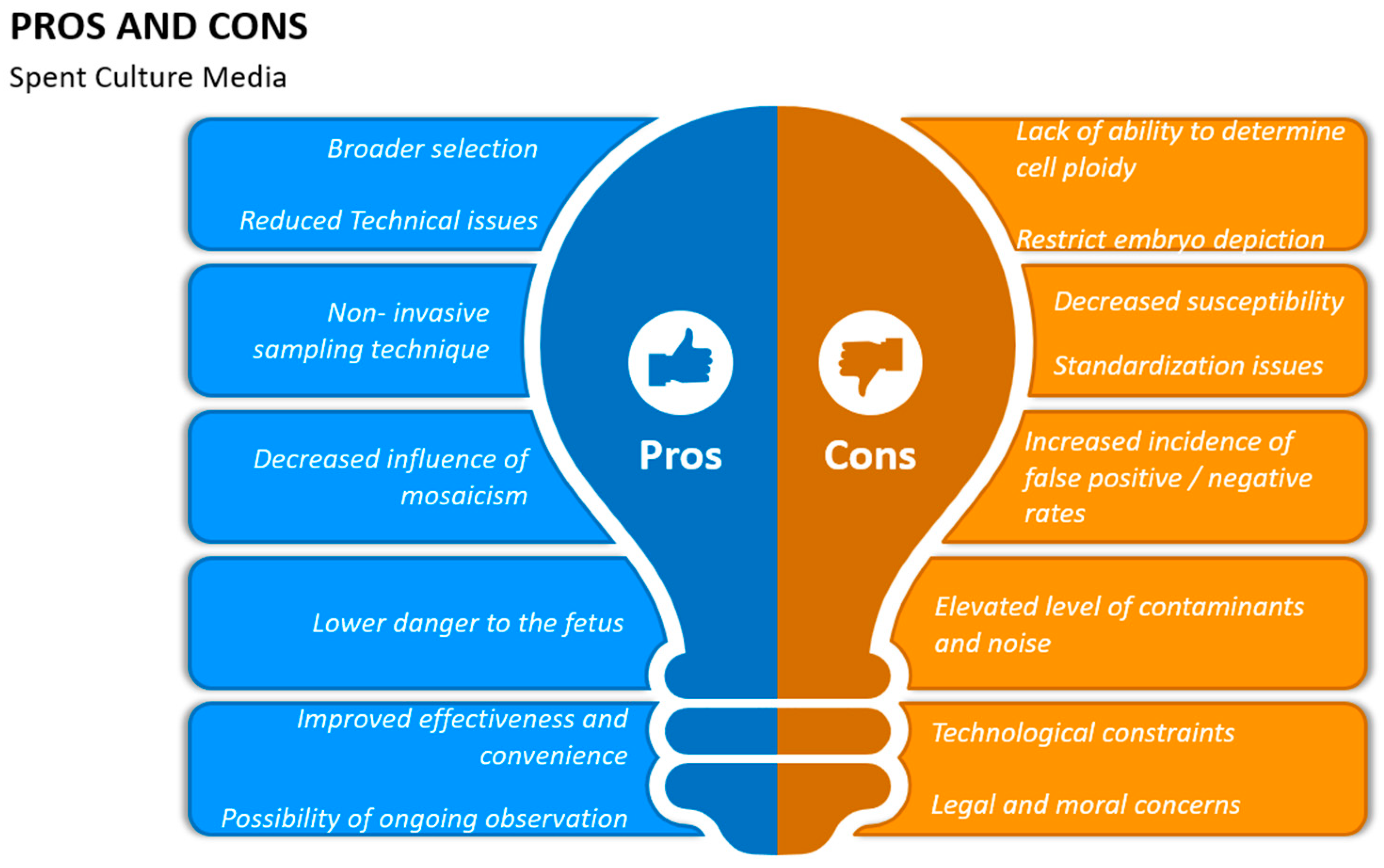
11. Spent Embryo Culture Medium (SCM)
12. Diagnostic Performance of Blastocyst Culture Media in Non-Invasive ΡGT
13. Diagnostic Performance of BF and SCM as PGT DNA Sources in Clinical Settings
14. Mosaicism
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Edwards, R.G.; Gardner, R.L. Sexing of Live Rabbit Blastocysts. Nature 1967, 214, 576–577. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, L.; Meng, L.; Liang, L.; Zhang, C. Advantages of vitrification preservation in assisted reproduction and potential influences on imprinted genes. Clin. Epigenet. 2022, 14, 141. [Google Scholar] [CrossRef]
- Wang, J.; Sauer, M.V. In vitro fertilization (IVF): A review of 3 decades of clinical innovation and technological advancement. Ther. Clin. Risk Manag. 2006, 2, 355–364. [Google Scholar] [CrossRef]
- Takeuchi, K. Pre-implantation genetic testing: Past, present, future. Reprod. Med. Biol. 2021, 20, 27–40. [Google Scholar] [CrossRef]
- Bar-El, L.; Kalma, Y.; Malcov, M.; Schwartz, T.; Raviv, S.; Cohen, T.; Amir, H.; Cohen, Y.; Reches, A.; Amit, A.; et al. Blastomere biopsy for PGD delays embryo compaction and blastulation: A time-lapse microscopic analysis. J. Assist. Reprod. Genet. 2016, 33, 1449–1457. [Google Scholar] [CrossRef]
- Chronopoulou, E.; Harper, J.C. IVF culture media: Past, present and future. Hum. Reprod. Update 2015, 21, 39–55. [Google Scholar] [CrossRef]
- Lal, A.; Roudebush, W.E.; Chosed, R.J. Embryo Biopsy Can Offer More Information Than Just Ploidy Status. Front. Cell Dev. Biol. 2020, 8, 78. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Steinberg, K.M.; Larson, D.E.; Wilson, R.K.; Mardis, E.R. The Next-Generation Sequencing Revolution and Its Impact on Genomics. Cell 2013, 155, 27–38. [Google Scholar] [CrossRef]
- De Rycke, M.; Berckmoes, V. Preimplantation Genetic Testing for Monogenic Disorders. Genes 2020, 11, 871. [Google Scholar] [CrossRef] [PubMed]
- Murtinger, M.; Schuff, M.; Wirleitner, B.; Miglar, S.; Spitzer, D. Comment on the recent PGDIS Position Statement on the Transfer of Mosaic Embryos 2021. J. Assist. Reprod. Genet. 2022, 39, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Viotti, M. Preimplantation Genetic Testing for Chromosomal Abnormalities: Aneuploidy, Mosaicism, and Structural Rearrangements. Genes 2020, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- Montag, M.; Van Der Ven, K.; Rösing, B.; Van Der Ven, H. Polar body biopsy: A viable alternative to preimplantation genetic diagnosis and screening. Reprod. Biomed. Online 2009, 18, 6–11. [Google Scholar] [CrossRef]
- Schenk, M.; Groselj-Strele, A.; Eberhard, K.; Feldmeier, E.; Kastelic, D.; Cerk, S.; Weiss, G. Impact of polar body biopsy on embryo morphokinetics—Back to the roots in preimplantation genetic testing? J. Assist. Reprod. Genet. 2018, 35, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.; Sullivan, E.A.; Li, Z.; Farquhar, C. Day 5 versus day 3 embryo biopsy for preimplantation genetic testing for monogenic/single gene defects. Cochrane Database Syst. Rev. 2018, 2018, CD013233. [Google Scholar] [CrossRef]
- Toyooka, Y. Trophoblast lineage specification in the mammalian preimplantation embryo. Reprod. Med. Biol. 2020, 19, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Burks, C.; Van Heertum, K.; Weinerman, R. The Technological Advances in Embryo Selection and Genetic Testing: A Look Back at the Evolution of Aneuploidy Screening and the Prospects of Non-Invasive PGT. Reprod. Med. 2021, 2, 26–34. [Google Scholar] [CrossRef]
- Aoyama, N.; Kato, K. Trophectoderm biopsy for preimplantation genetic test and technical tips: A review. Reprod. Med. Biol. 2020, 19, 222–231. [Google Scholar] [CrossRef]
- Scott, R.T.; Upham, K.M.; Forman, E.J.; Zhao, T.; Treff, N.R. Cleavage-stage biopsy significantly impairs human embryonic implantation potential while blastocyst biopsy does not: A randomized and paired clinical trial. Fertil. Steril. 2013, 100, 624–630. [Google Scholar] [CrossRef]
- Forman, E.J.; Hong, K.H.; Ferry, K.M.; Tao, X.; Taylor, D.; Levy, B.; Treff, N.R.; Scott, R.T. In vitro fertilization with single euploid blastocyst transfer: A randomized controlled trial. Fertil. Steril. 2013, 100, 100–107.e1. [Google Scholar] [CrossRef]
- Sunkara, S.K.; LaMarca, A.; Polyzos, N.P.; Seed, P.T.; Khalaf, Y. Live birth and perinatal outcomes following stimulated and unstimulated IVF: Analysis of over two decades of a nationwide data. Hum. Reprod. 2016, 31, 2261–2267. [Google Scholar] [CrossRef]
- Capalbo, A.; Rienzi, L. Mosaicism between trophectoderm and inner cell mass. Fertil. Steril. 2017, 107, 1098–1106. [Google Scholar] [CrossRef]
- Lee, C.I.; Cheng, E.H.; Lee, M.S.; Lin, P.Y.; Chen, Y.C.; Chen, C.H.; Huang, L.-S.; Huang, C.-C.; Lee, T.-H. Healthy live births from transfer of low-mosaicism embryos after preimplantation genetic testing for aneuploidy. J. Assist. Reprod. Genet. 2020, 37, 2305–2313. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, Q.; Cinnioglu, C.; Kayali, R.; Lathi, R.B.; Behr, B. The impact of culture conditions on blastocyst formation and aneuploidy rates: A comparison between single-step and sequential media in a large academic practice. J. Assist. Reprod. Genet. 2020, 37, 161–169. [Google Scholar] [CrossRef]
- Capalbo, A.; Romanelli, V.; Patassini, C.; Poli, M.; Girardi, L.; Giancani, A.; Stoppa, M.; Cimadomo, D.; Ubaldi, F.M.; Rienzi, L. Diagnostic efficacy of blastocoel fluid and spent media as sources of DNA for preimplantation genetic testing in standard clinical conditions. Fertil. Steril. 2018, 110, 870–879.e5. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular Stress Responses: Cell Survival and Cell Death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef]
- Pisko, J.; Špirková, A.; Čikoš, Š.; Olexiková, L.; Kovaříková, V.; Šefčíková, Z.; Fabian, D. Apoptotic cells in mouse blastocysts are eliminated by neighbouring blastomeres. Sci. Rep. 2021, 11, 9228. [Google Scholar] [CrossRef]
- Bolton, H.; Graham, S.J.L.; Van Der Aa, N.; Kumar, P.; Theunis, K.; Fernandez Gallardo, E.; Voet, T.; Zernicka-Goetz, M. Mouse model of chromosome mosaicism reveals lineage-specific depletion of aneuploid cells and normal developmental potential. Nat. Commun. 2016, 7, 11165. [Google Scholar] [CrossRef]
- Fabian, M.A.; Biggs, W.H.; Treiber, D.K.; Atteridge, C.E.; Azimioara, M.D.; Benedetti, M.G.; Carter, T.A.; Ciceri, P.; Edeen, P.T.; Floyd, M.; et al. A small molecule–kinase interaction map for clinical kinase inhibitors. Nat. Biotechnol. 2005, 23, 329–336. [Google Scholar] [CrossRef]
- Matassov, D.; Kagan, T.; Leblanc, J.; Sikorska, M.; Zakeri, Z. Measurement of Apoptosis by DNA Fragmentation. In Apoptosis Methods and Protocols; Humana Press: Totowa, NJ, USA, 2004; pp. 1–18. Available online: http://link.springer.com/10.1385/1-59259-812-9:001 (accessed on 15 January 2024).
- Zhang, Y.; Li, N.; Wang, L.; Sun, H.; Ma, M.; Wang, H.; Xu, X.; Zhang, W.; Liu, Y.; Cram, D.S.; et al. Molecular analysis of DNA in blastocoele fluid using next-generation sequencing. J. Assist. Reprod. Genet. 2016, 33, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Thowfeequ, S.; Srinivas, S. Embryonic and extraembryonic tissues during mammalian development: Shifting boundaries in time and space. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210255. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.; Handyside, A.H.; Winston, R.M.L. The human blastocyst: Cell number, death and allocation during late preimplantation development in vitro. Development 1989, 107, 597–604. [Google Scholar] [CrossRef]
- Brison, D.R. Apoptosis in mammalian preimplantation embryos: Regulation by survival factors. Hum. Fertil. 2000, 3, 36–47. [Google Scholar] [CrossRef]
- Kakourou, G.; Mamas, T.; Vrettou, C.; Traeger-Synodinos, J. An Update on Non-invasive Approaches for Genetic Testing of the Preimplantation Embryo. Curr. Genom. 2022, 23, 337–352. [Google Scholar] [CrossRef]
- Magli, M.C.; Pomante, A.; Cafueri, G.; Valerio, M.; Crippa, A.; Ferraretti, A.P.; Gianaroli, L. Preimplantation genetic testing: Polar bodies, blastomeres, trophectoderm cells, or blastocoelic fluid? Fertil. Steril. 2016, 105, 676–683.e5. [Google Scholar] [CrossRef]
- Rule, K.; Chosed, R.J.; Arthur Chang, T.; David Wininger, J.; Roudebush, W.E. Relationship between blastocoel cell-free DNA and day-5 blastocyst morphology. J. Assist. Reprod. Genet. 2018, 35, 1497–1501. [Google Scholar] [CrossRef]
- Magli, M.C.; Albanese, C.; Crippa, A.; Tabanelli, C.; Ferraretti, A.P.; Gianaroli, L. Deoxyribonucleic acid detection in blastocoelic fluid: A new predictor of embryo ploidy and viable pregnancy. Fertil. Steril. 2019, 111, 77–85. [Google Scholar] [CrossRef]
- Shi, W.; Zhao, Z.; Xue, X.; Li, Q.; Yao, Y.; Wang, D.; Wang, J.; Lu, S.; Shi, J. Ploidy Testing of Blastocoel Fluid for Screening May Be Technically Challenging and More Invasive Than That of Spent Cell Culture Media. Front. Physiol. 2022, 13, 794210. [Google Scholar] [CrossRef]
- Poli, M.; Ori, A.; Child, T.; Jaroudi, S.; Spath, K.; Beck, M.; Wells, D. Characterization and quantification of proteins secreted by single human embryos prior to implantation. EMBO Mol. Med. 2015, 7, 1465–1479. [Google Scholar] [CrossRef]
- Battaglia, R.; Palini, S.; Vento, M.E.; La Ferlita, A.; Lo Faro, M.J.; Caroppo, E.; Borzì, P.; Falzone, L.; Barbagallo, D.; Ragusa, M.; et al. Identification of extracellular vesicles and characterization of miRNA expression profiles in human blastocoel fluid. Sci. Rep. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.; Bolumar, D.; Amadoz, A.; Jimenez-Almazán, J.; Valbuena, D.; Vilella, F.; Moreno, I. Identification and Characterization of Extracellular Vesicles and Its DNA Cargo Secreted During Murine Embryo Development. Genes 2020, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Tomic, M.; Vrtacnik Bokal, E.; Stimpfel, M. Non-Invasive Preimplantation Genetic Testing for Aneuploidy and the Mystery of Genetic Material: A Review Article. Int. J. Mol. Sci. 2022, 23, 3568. [Google Scholar] [CrossRef] [PubMed]
- Michailov, Y.; Friedler, S.; Saar-Ryss, B. Methods to improve frozen-thawed blastocyst transfer outcomes- the IVF laboratory perspective. J. IVF-Worldw. 2023, 1, 1–15. [Google Scholar] [CrossRef]
- Sialakouma, A.; Karakasiliotis, I.; Ntala, V.; Nikolettos, N.; Asimakopoulos, B. Embryonic Cell-free DNA in Spent Culture Medium: A Non-invasive Tool for Aneuploidy Screening of the Corresponding Embryos. In Vivo 2021, 35, 3449–3457. [Google Scholar] [CrossRef] [PubMed]
- Brouillet, S.; Martinez, G.; Coutton, C.; Hamamah, S. Is cell-free DNA in spent embryo culture medium an alternative to embryo biopsy for preimplantation genetic testing? A systematic review. Reprod. Biomed. Online 2020, 40, 779–796. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Dheedene, A.; Christodoulou, C.; Taelman, J.; Dhaenens, L.; Van Nieuwerburgh, F.; Deforce, D.; Van den Abbeel, E.; De Sutter, P.; Menten, B.; et al. Chromosomal mosaicism in human blastocysts: The ultimate challenge of preimplantation genetic testing? Hum. Reprod. 2018, 33, 1342–1354. [Google Scholar] [CrossRef] [PubMed]
- Victor, A.R.; Griffin, D.K.; Brake, A.J.; Tyndall, J.C.; Murphy, A.E.; Lepkowsky, L.T.; Lal, A.; Zouves, C.G.; Barnes, F.L.; McCoy, R.C.; et al. Assessment of aneuploidy concordance between clinical trophectoderm biopsy and blastocyst. Hum. Reprod. 2019, 34, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, B.; El Khatib, I.; Liñán, A.; Bayram, A.; Arnanz, A.; Chopra, R.; De Munck, N.; Fatemi, H.M. The clinicians’ dilemma with mosaicism—An insight from inner cell mass biopsies. Hum. Reprod. 2019, 34, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.H.; Hsieh, J.Y.; Lee, M.J.; Lai, H.H.; Hsieh, C.L.; Wang, H.L.; Chang, Y.-J.; Chen, S.-U. Concordance between different trophectoderm biopsy sites and the inner cell mass of chromosomal composition measured with a next-generation sequencing platform. MHR Basic Sci. Reprod. Med. 2018, 24, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Dhaenens, L.; Taelman, J.; Dheedene, A.; Bialecka, M.; De Sutter, P.; Lopes, S.M.C.d.S.; Menten, B.; Heindryckx, B. Extended in vitro culture of human embryos demonstrates the complex nature of diagnosing chromosomal mosaicism from a single trophectoderm biopsy. Hum. Reprod. 2019, 34, 758–769. [Google Scholar] [CrossRef]
- Girardi, L.; Figliuzzi, M.; Poli, M.; Serdarogullari, M.; Patassini, C.; Caroselli, S.; Pergher, I.; Cogo, F.; Coban, O.; Boynukalin, F.K.; et al. The use of copy number loads to designate mosaicism in blastocyst stage PGT-A cycles: Fewer is better. Hum. Reprod. 2023, 38, 982–991. [Google Scholar] [CrossRef]
- Cimadomo, D.; Capalbo, A.; Ubaldi, F.M.; Scarica, C.; Palagiano, A.; Canipari, R.; Rienzi, L. The Impact of Biopsy on Human Embryo Developmental Potential during Preimplantation Genetic Diagnosis. BioMed Res. Int. 2016, 2016, 7193075. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, A.; Ubaldi, F.M.; Rienzi, L.; Scott, R.; Treff, N. Detecting mosaicism in trophectoderm biopsies: Current challenges and future possibilities. Hum. Reprod. 2017, 32, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Sharma, R. Non invasive assessment of human oocytes and embryos in assisted reproduction: Review on present practices and future trends. Med. J. Armed. Forces India 2022, 78, 7–16. [Google Scholar] [CrossRef]
- Armstrong, S.; Bhide, P.; Jordan, V.; Pacey, A.; Farquhar, C. Time-lapse systems for embryo incubation and assessment in assisted reproduction. Cochrane Database Syst. Rev. 2018, 2018, CD011320. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhang, S.; Gu, Y.; Gong, F.; Kong, L.; Lu, G.; Lin, G.; Liang, B.; Hu, L. Non-invasive Metabolomic Profiling of Embryo Culture Medium Using Raman Spectroscopy With Deep Learning Model Predicts the Blastocyst Development Potential of Embryos. Front. Physiol. 2021, 12, 777259. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.; Guerra-Carvalho, B.; Crisóstomo, L.; Rocha, A.; Barros, A.; Alves, M.G.; Oliveira, P.F. Metabolomics Integration in Assisted Reproductive Technologies for Enhanced Embryo Selection beyond Morphokinetic Analysis. Int. J. Mol. Sci. 2023, 25, 491. [Google Scholar] [CrossRef] [PubMed]
- Raudonis, V.; Paulauskaite-Taraseviciene, A.; Sutiene, K.; Jonaitis, D. Towards the automation of early-stage human embryo development detection. Biomed. Eng. OnLine 2019, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Paulson, R.J.; Reichman, D.E.; Zaninovic, N.; Goodman, L.R.; Racowsky, C. Time-lapse imaging: Clearly useful to both laboratory personnel and patient outcomes versus just because we can doesn’t mean we should. Fertil. Steril. 2018, 109, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Sun, H.X. Blastocyst transfer ameliorates live birth rate compared with cleavage-stage embryos transfer in fresh in vitro fertilization or intracytoplasmic sperm injection cycles: Reviews and meta-analysis. Yonsei Med. J. 2014, 55, 815–825. [Google Scholar] [CrossRef]
- Wong, C.C.; Loewke, K.E.; Bossert, N.L.; Behr, B.; De Jonge, C.J.; Baer, T.M.; Reijo Pera, R.A. Non-invasive imaging of human embryos before embryonic genome activation predicts development to the blastocyst stage. Nat. Biotechnol. 2010, 28, 1115–1121. [Google Scholar] [CrossRef]
- Lundervold, A.S.; Lundervold, A. An overview of deep learning in medical imaging focusing on MRI. Z. Med. Phys. 2019, 29, 102–127. [Google Scholar] [CrossRef]
- Khosravi, P.; Kazemi, E.; Zhan, Q.; Malmsten, J.E.; Toschi, M.; Zisimopoulos, P.; Sigaras, A.; Lavery, S.; Cooper, L.A.D.; Hickman, C.; et al. Deep learning enables robust assessment and selection of human blastocysts after in vitro fertilization. NPJ Digit. Med. 2019, 2, 21. [Google Scholar] [CrossRef]
- Malmsten, J.; Zaninovic, N.; Zhan, Q.; Toschi, M.; Rosenwaks, Z.; Shan, J. Automatic prediction of embryo cell stages using artificial intelligence convolutional neural network. Fertil. Steril. 2018, 110, e360. [Google Scholar] [CrossRef]
- Motiei, M.; Vaculikova, K.; Cela, A.; Tvrdonova, K.; Khalili, R.; Rumpik, D.; Rumpikova, T.; Glatz, Z.; Saha, T. Non-Invasive Human Embryo Metabolic Assessment as a Developmental Criterion. J. Clin. Med. 2020, 9, 4094. [Google Scholar] [CrossRef]
- Bartolacci, A.; Tondo, F.; Alteri, A.; Solano Narduche, L.; De Girolamo, S.; D’Alessandro, G.; Rabellotti, E.; Papaleo, E.; Pagliardini, L. The Task Matters: A Comprehensive Review and Proposed Literature Score of the Effects of Chemical and Physical Parameters on Embryo Developmental Competence. Life 2023, 13, 2161. [Google Scholar] [CrossRef]
- Lundin, K.; Park, H. Time-lapse technology for embryo culture and selection. Upsala J. Med. Sci. 2020, 125, 77–84. [Google Scholar] [CrossRef]
- Villena Gonzales, W.; Mobashsher, A.; Abbosh, A. The Progress of Glucose Monitoring—A Review of Invasive to Minimally and Non-Invasive Techniques, Devices and Sensors. Sensors 2019, 19, 800. [Google Scholar] [CrossRef]
- Li, P.; Song, Z.; Yao, Y.; Huang, T.; Mao, R.; Huang, J.; Ma, Y.; Dong, X.; Huang, W.; Huang, J.; et al. Preimplantation Genetic Screening with Spent Culture Medium/Blastocoel Fluid for in Vitro Fertilization. Sci. Rep. 2018, 8, 9275. [Google Scholar] [CrossRef]
- Wu, A.K.; Elliott, P.; Katz, P.P.; Smith, J.F. Time costs of fertility care: The hidden hardship of building a family. Fertil. Steril. 2013, 99, 2025–2030. [Google Scholar] [CrossRef]
- Wilton, L.; Thornhill, A.; Traeger-Synodinos, J.; Sermon, K.D.; Harper, J.C. The causes of misdiagnosis and adverse outcomes in PGD. Hum. Reprod. 2009, 24, 1221–1228. [Google Scholar] [CrossRef]
- Bouba, I.; Hatzi, E.; Ladias, P.; Sakaloglou, P.; Kostoulas, C.; Georgiou, I. Biological and Clinical Significance of Mosaicism in Human Preimplantation Embryos. J. Dev. Biol. 2021, 9, 18. [Google Scholar] [CrossRef]
- Gardner, D.K.; Wale, P.L.; Collins, R.; Lane, M. Glucose consumption of single post-compaction human embryos is predictive of embryo sex and live birth outcome. Hum. Reprod. 2011, 26, 1981–1986. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Larman, M.G.; Thouas, G.A. Sex-related physiology of the preimplantation embryo. Mol. Hum. Reprod. 2010, 16, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.J.; Natale, D.R.; Barcroft, L.C. Molecular regulation of blastocyst formation. Anim. Reprod. Sci. 2004, 82–83, 583–592. [Google Scholar] [CrossRef]
- Perloe, M.; Welch, C.; Morton, P.; Venier, W.; Wells, D.; Palini, S. Validation of blastocoele fluid aspiration for preimplantation genetic screening using array comparative genomic hybridization (aCGH). Fertil. Steril. 2013, 100, S208. [Google Scholar] [CrossRef]
- Gianaroli, L.; Magli, M.C.; Pomante, A.; Crivello, A.M.; Cafueri, G.; Valerio, M.; Ferraretti, A.P. Blastocentesis: A source of DNA for preimplantation genetic testing. Results from a pilot study. Fertil. Steril. 2014, 102, 1692–1699.e6. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.; Jaroudi, S.; Sarasa, J.; Spath, K.; Child, T.; Wells, D. The blastocoel fluid as a source of DNA for preimplantation genetic diagnosis and screening. Fertil. Steril. 2013, 100, S37. [Google Scholar] [CrossRef]
- Fang, F.; Li, Z.; Yu, J.; Long, Y.; Zhao, Q.; Ding, X.; Wu, L.; Shao, S.; Zhang, L.; Xiang, W. MicroRNAs secreted by human embryos could be potential biomarkers for clinical outcomes of assisted reproductive technology. J. Adv. Res. 2021, 31, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Stigliani, S.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA content in embryo culture medium is significantly associated with human embryo fragmentation. Hum. Reprod. 2013, 28, 2652–2660. [Google Scholar] [CrossRef]
- Lledo, B.; Morales, R.; Antonio Ortiz, J.; Bernabeu, A.; Bernabeu, R. Noninvasive preimplantation genetic testing using the embryo spent culture medium: An update. Curr. Opin. Obstet. Gynecol. 2023, 35, 294–299. [Google Scholar] [CrossRef]
- Ma, Y.; Gu, M.; Chen, L.; Shen, H.; Pan, Y.; Pang, Y.; Miao, S.; Tong, R.; Huang, H.; Zhu, Y.; et al. Recent advances in critical nodes of embryo engineering technology. Theranostics 2021, 11, 7391–7424. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.; Spanos, S.; Becker, D.; Iannelli, P.; Winston, R.M.; Stark, J. From cell death to embryo arrest: Mathematical models of human preimplantation embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Cecchele, A.; Cermisoni, G.C.; Giacomini, E.; Pinna, M.; Vigano, P. Cellular and Molecular Nature of Fragmentation of Human Embryos. Int. J. Mol. Sci. 2022, 23, 1349. [Google Scholar] [CrossRef] [PubMed]
- Stigliani, S.; Persico, L.; Lagazio, C.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA in Day 3 embryo culture medium is a novel, non-invasive biomarker of blastocyst potential and implantation outcome. MHR Basic Sci. Reprod. Med. 2014, 20, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.R.; McGillivray, B.C.; Wicker, S.M.; Peek, J.C.; Shelling, A.N.; Stone, P.; Chamley, L.W.; Cree, L.M. Characterizing nuclear and mitochondrial DNA in spent embryo culture media: Genetic contamination identified. Fertil. Steril. 2017, 107, 220–228.e5. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.N.; Monteiro, P.B. Non-invasive preimplantation genetic testing: A literature review. JBRA Assist. Reprod. 2022, 26, 554–558. [Google Scholar]
- Sepúlveda, S.; Garcia, J.; Arriaga, E.; Diaz, J.; Noriega-Portella, L.; Noriega-Hoces, L. In vitro development and pregnancy outcomes for human embryos cultured in either a single medium or in a sequential media system. Fertil. Steril. 2009, 91, 1765–1770. [Google Scholar] [CrossRef] [PubMed]
- Minasi, M.G.; Fabozzi, G.; Casciani, V.; Lobascio, A.M.; Colasante, A.; Scarselli, F.; Greco, E. Improved blastocyst formation with reduced culture volume: Comparison of three different culture conditions on 1128 sibling human zygotes. J. Assist. Reprod. Genet. 2015, 32, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jia, L.; Li, T.; Guo, Y.; He, S.; Zhang, Z.; Su, W.; Zhang, S.; Fang, C. Diagnostic efficiency of blastocyst culture medium in noninvasive preimplantation genetic testing. F&S Rep. 2021, 2, 88–94. [Google Scholar]
- Popovic, M.; Dhaenens, L.; Boel, A.; Menten, B.; Heindryckx, B. Chromosomal mosaicism in human blastocysts: The ultimate diagnostic dilemma. Hum. Reprod. Update 2020, 26, 313–334. [Google Scholar] [CrossRef]
- Esfandiari, N.; Bunnell, M.E.; Casper, R.F. Human embryo mosaicism: Did we drop the ball on chromosomal testing? J. Assist. Reprod. Genet. 2016, 33, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Cram, D.S.; Leigh, D.; Handyside, A.; Rechitsky, L.; Xu, K.; Harton, G.; Grifo, J.; Rubio, C.; Fragouli, E.; Kahraman, S.; et al. PGDIS Position Statement on the Transfer of Mosaic Embryos 2019. Reprod. Biomed. Online 2019, 39, e1–e4. [Google Scholar] [CrossRef] [PubMed]
- Munné, S.; Wells, D. Detection of mosaicism at blastocyst stage with the use of high-resolution next-generation sequencing. Fertil. Steril. 2017, 107, 1085–1091. [Google Scholar] [CrossRef]
- Gajecka, M. Unrevealed mosaicism in the next-generation sequencing era. Mol. Genet. Genom. 2016, 291, 513–530. [Google Scholar] [CrossRef]
- Neal, S.A.; Franasiak, J.M.; Forman, E.J.; Werner, M.D.; Morin, S.J.; Tao, X.; Treff, N.R.; Scott, R.T. High relative deoxyribonucleic acid content of trophectoderm biopsy adversely affects pregnancy outcomes. Fertil. Steril. 2017, 107, 731–736.e1. [Google Scholar] [CrossRef] [PubMed]
- Palini, S.; Galluzzi, L.; De Stefani, S.; Bianchi, M.; Wells, D.; Magnani, M.; Bulletti, C. Genomic DNA in human blastocoele fluid. Reprod. Biomed. Online 2013, 26, 603–610. [Google Scholar] [CrossRef]
- Galluzzi, L.; Palini, S.; Stefani, S.D.; Andreoni, F.; Primiterra, M.; Diotallevi, A.; Bulletti, C.; Magnani, M. Extracellular embryo genomic DNA and its potential for genotyping applications. Future Sci. OA 2015, 1, fso.15.62. [Google Scholar] [CrossRef] [PubMed]
- Vera-Rodriguez, M.; Diez-Juan, A.; Jimenez-Almazan, J.; Martinez, S.; Navarro, R.; Peinado, V.; Mercader, A.; Meseguer, M.; Blesa, D.; Moreno, I.; et al. Origin and composition of cell-free DNA in spent medium from human embryo culture during preimplantation development. Hum. Reprod. 2018, 33, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Rienzi, L.; Navarro-Sánchez, L.; Cimadomo, D.; García-Pascual, C.M.; Albricci, L.; Soscia, D.; Valbuena, D.; Capalbo, A.; Ubaldi, F.; et al. Embryonic cell-free DNA versus trophectoderm biopsy for aneuploidy testing: Concordance rate and clinical implications. Fertil. Steril. 2019, 112, 510–519. [Google Scholar] [CrossRef]
- Huang, L.; Bogale, B.; Tang, Y.; Lu, S.; Xie, X.S.; Racowsky, C. Noninvasive preimplantation genetic testing for aneuploidy in spent medium may be more reliable than trophectoderm biopsy. Proc. Natl. Acad. Sci. USA 2019, 116, 14105–14112. [Google Scholar] [CrossRef]
- Tobler, K.J.; Zhao, Y.; Ross, R.; Benner, A.T.; Xu, X.; Du, L.; Broman, K.; Thrift, K.; Brezina, P.R.; Kearns, W.G. Blastocoel fluid from differentiated blastocysts harbors embryonic genomic material capable of a whole-genome deoxyribonucleic acid amplification and comprehensive chromosome microarray analysis. Fertil. Steril. 2015, 104, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Shi, B.; Sagnelli, M.; Yang, D.; Yao, Y.; Li, W.; Shao, L.; Lu, S.; Li, D.; Wang, X. Minimally invasive preimplantation genetic testing using blastocyst culture medium. Hum. Reprod. 2019, 34, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Q.S.Y.; Zhang, Y.X.; Chung, J.P.W.; Lui, W.T.; Kwok, Y.K.Y.; Gui, B.; Kong, G.W.S.; Cao, Y.; Li, T.C.; Choy, K.W. A prospective study of non-invasive preimplantation genetic testing for aneuploidies (NiPGT-A) using next-generation sequencing (NGS) on spent culture media (SCM). J. Assist. Reprod. Genet. 2019, 36, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Fang, R.; Chen, L.; Chen, D.; Xiao, J.P.; Yang, W.; Wang, H.; Song, X.; Ma, T.; Bo, S.; et al. Noninvasive chromosome screening of human embryos by genome sequencing of embryo culture medium for in vitro fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, 11907–11912. [Google Scholar] [CrossRef] [PubMed]
- Patrizio, P.; Shoham, G.; Shoham, Z.; Leong, M.; Barad, D.H.; Gleicher, N. Worldwide live births following the transfer of chromosomally “Abnormal” embryos after PGT/A: Results of a worldwide web-based survey. J. Assist. Reprod. Genet. 2019, 36, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Vera-Rodriguez, M.; Rubio, C. Assessing the true incidence of mosaicism in preimplantation embryos. Fertil. Steril. 2017, 107, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Chen, Z.; Yin, M.; Deng, Y.; Liang, Y.; Wang, W.; Yao, Y.; Sun, L. Re-analysis of whole blastocysts after trophectoderm biopsy indicated chromosome aneuploidy. Hum. Genom. 2020, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Leaver, M.; Wells, D. Non-invasive preimplantation genetic testing (niPGT): The next revolution in reproductive genetics? Hum. Reprod. Update 2020, 26, 16–42. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, N.M.; Maxwell, S.M.; Besser, A.G.; Grifo, J.A. Diagnosis and clinical management of embryonic mosaicism. Fertil. Steril. 2017, 107, 6–11. [Google Scholar] [CrossRef]
- Maxwell, S.M.; Colls, P.; Hodes-Wertz, B.; McCulloh, D.H.; McCaffrey, C.; Wells, D.; Munné, S.; Grifo, J.A. Why do euploid embryos miscarry? A case-control study comparing the rate of aneuploidy within presumed euploid embryos that resulted in miscarriage or live birth using next-generation sequencing. Fertil. Steril. 2016, 106, 1414–1419.e5. [Google Scholar] [CrossRef]
- Wells, D.; Kaur, K.; Grifo, J.; Glassner, M.; Taylor, J.C.; Fragouli, E.; Munne, S. Clinical utilisation of a rapid low-pass whole genome sequencing technique for the diagnosis of aneuploidy in human embryos prior to implantation. J. Med. Genet. 2014, 51, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Greco, E.; Minasi, M.G.; Fiorentino, F. Healthy Babies after Intrauterine Transfer of Mosaic Aneuploid Blastocysts. N. Engl. J. Med. 2015, 373, 2089–2090. [Google Scholar] [CrossRef] [PubMed]
- Spinella, F.; Fiorentino, F.; Biricik, A.; Bono, S.; Ruberti, A.; Cotroneo, E.; Baldi, M.; Cursio, E.; Minasi, M.G.; Greco, E. Extent of chromosomal mosaicism influences the clinical outcome of in vitro fertilization treatments. Fertil. Steril. 2018, 109, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zore, T.; Kroener, L.L.; Wang, C.; Liu, L.; Buyalos, R.; Hubert, G.; Shamonki, M. Transfer of embryos with segmental mosaicism is associated with a significant reduction in live-birth rate. Fertil. Steril. 2019, 111, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Alfarawati, S.; Spath, K.; Babariya, D.; Tarozzi, N.; Borini, A.; Wells, D. Analysis of implantation and ongoing pregnancy rates following the transfer of mosaic diploid–aneuploid blastocysts. Hum. Genet. 2017, 136, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.R.; Arrach, N.; Rhodes-Long, K.; Ahmady, A.; Ingles, S.; Chung, K.; Bendikson, K.A.; Paulson, R.J.; McGinnis, L.K. Pushing the limits of detection: Investigation of cell-free DNA for aneuploidy screening in embryos. Fertil. Steril. 2018, 110, 467–475.e2. [Google Scholar] [CrossRef] [PubMed]
- Darwish, E.; Magdi, Y. Artificial shrinkage of blastocoel using a laser pulse prior to vitrification improves clinical outcome. J. Assist. Reprod. Genet. 2016, 33, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Shamonki, M.I.; Jin, H.; Haimowitz, Z.; Liu, L. Proof of concept: Preimplantation genetic screening without embryo biopsy through analysis of cell-free DNA in spent embryo culture media. Fertil. Steril. 2016, 106, 1312–1318. [Google Scholar] [CrossRef]
- Wu, H.; Ding, C.; Shen, X.; Wang, J.; Li, R.; Cai, B.; Xu, Y.; Zhong, Y.; Zhou, C. Medium-Based Noninvasive Preimplantation Genetic Diagnosis for Human α-Thalassemias-SEA. Medicine 2015, 94, e669. [Google Scholar] [CrossRef]
- Giacomini, E.; Vago, R.; Sanchez, A.M.; Podini, P.; Zarovni, N.; Murdica, V.; Rizzo, R.; Bortolotti, D.; Candiani, M.; Viganò, P. Secretome of in vitro cultured human embryos contains extracellular vesicles that are uptaken by the maternal side. Sci. Rep. 2017, 7, 5210. [Google Scholar] [CrossRef]
- Bankoglu, E.E.; Stipp, F.; Gerber, J.; Seyfried, F.; Heidland, A.; Bahner, U.; Stopper, H. Effect of cryopreservation on DNA damage and DNA repair activity in human blood samples in the comet assay. Arch. Toxicol. 2021, 95, 1831–1841. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.D.; Scott, K.; Bohrer, C.; Gabriele, D.; Tao, X.; Hong, K.H.; Taylor, D.; Treff, N.; Scott, R. The accuracy of blastocoel fluid comprehensive chromosomal screening (CCS) is dependent on amplification yield and sequencing depth when using nextgen sequencing. Fertil. Steril. 2014, 102, e308–e309. [Google Scholar] [CrossRef]
- Anvari, M.; Gharib, A.; Abolhasani, M.; Azari-Yaam, A.; Gharalari, F.; Safavi, M.; Zare-Mirzaie, A.; Vasei, M. Pre-analytical Practices in the Molecular Diagnostic Tests, A Concise Review. Iran. J. Pathol. 2021, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, J.; Osei-Owusu, I.A.; Avigdor, B.E.; Tupler, R.; Pevsner, J. Mosaicism in Human Health and Disease. Annu. Rev. Genet. 2020, 54, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Mourad, A.; Antaki, R.; Bissonnette, F.; Al Baini, O.; Saadeh, B.; Jamal, W. Evidence-based clinical prioritization of embryos with mosaic results: A systematic review and meta-analysis. J. Assist. Reprod. Genet. 2021, 38, 2849–2860. [Google Scholar] [CrossRef] [PubMed]
- Lledó, B.; Morales, R.; Ortiz, J.A.; Blanca, H.; Ten, J.; Llácer, J.; Bernabeu, R. Implantation potential of mosaic embryos. Syst. Biol. Reprod. Med. 2017, 63, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Tšuiko, O.; Zhigalina, D.I.; Jatsenko, T.; Skryabin, N.A.; Kanbekova, O.R.; Artyukhova, V.G.; Svetlakov, A.V.; Teearu, K.; Trošin, A.; Salumets, A.; et al. Karyotype of the blastocoel fluid demonstrates low concordance with both trophectoderm and inner cell mass. Fertil. Steril. 2018, 109, 1127–1134.e1. [Google Scholar] [CrossRef] [PubMed]
- McCoy, R.C. Mosaicism in Preimplantation Human Embryos: When Chromosomal Abnormalities Are the Norm. Trends Genet. 2017, 33, 448–463. [Google Scholar] [CrossRef]
- Li, X.; Hao, Y.; Elshewy, N.; Zhu, X.; Zhang, Z.; Zhou, P. The mechanisms and clinical application of mosaicism in preimplantation embryos. J. Assist. Reprod. Genet. 2020, 37, 497–508. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
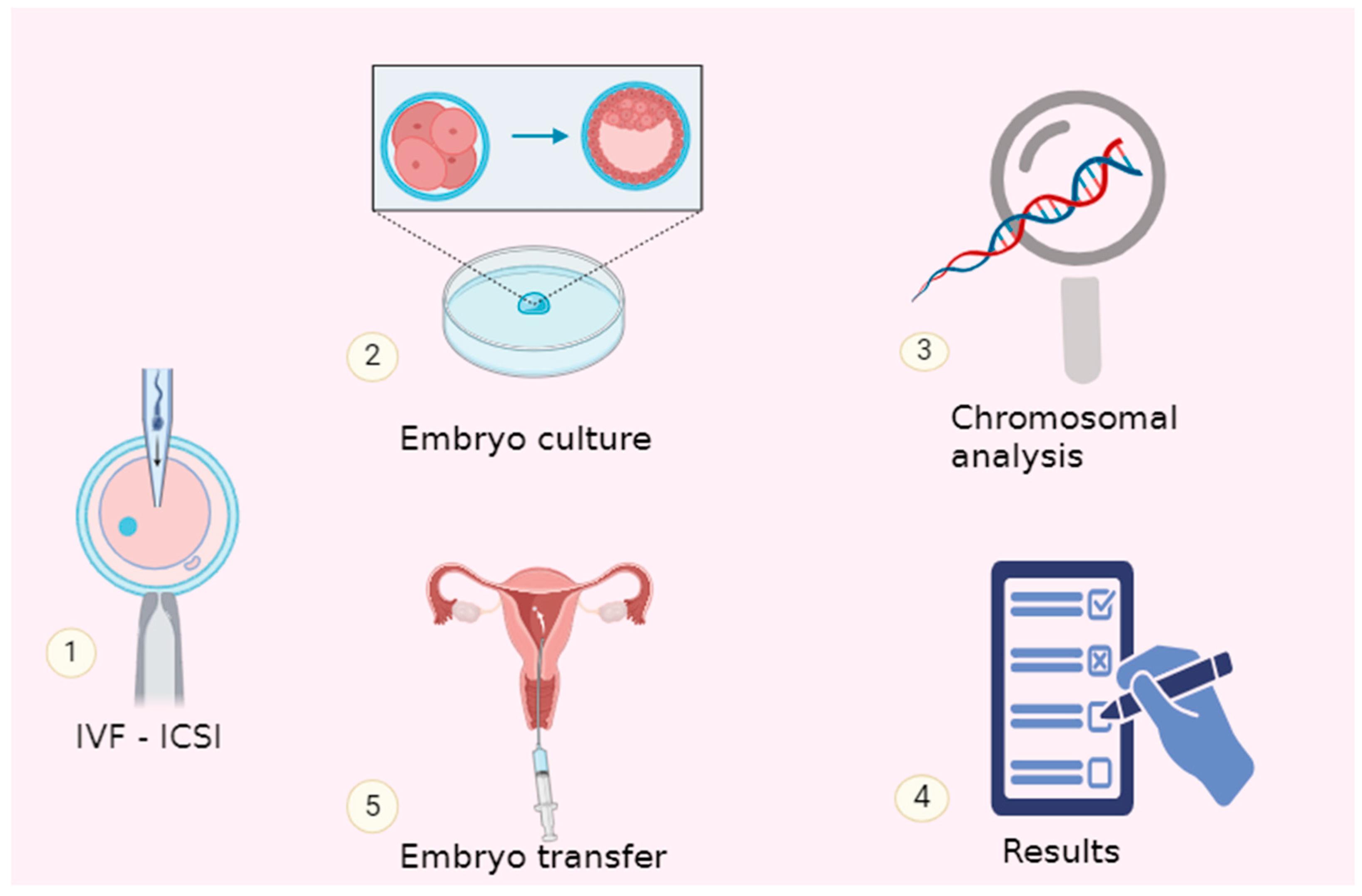{kind=link}
| Invasive PGT | |||
|---|---|---|---|
| FISH | aCGH | NGS | |
|
Green: chromosomal region in excess Red: chromosomal region in defect |
| |
| Minimally invasive PGT | Non-invasive PGT | ||
| Blastocoel fluid (BF) | Spent Culture Media (SCM) | ||
|
| ||
| Cell-Free DNA SCM | Cell-Free DNA BF |
|---|---|
| The ploidy of the embryo was unaffected by cfDNA. | Cells separate into TE and ICM on the 4th day of blastocyst development. Blastocoel forms withing the blastocyst during cavitation. |
| An estimated 86–94% of instances involve contamination with maternal genetic material. | Through blastocentesis → BF aspirated. About 0.01 μL of BF can be isolated. |
| The seed embryo’s embryonic DNA content ranged from 0% to 100%. Not every embryo’s embryonic genome may be evenly represented in the seed embryo. | Only 34.8% of BF samples were able to produce a signal suitable for embryo karyotyping. This outcome might have been caused by the fact that fresh blastocysts were used for the blastocentesis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustakli, E.; Zikopoulos, A.; Skentou, C.; Bouba, I.; Dafopoulos, K.; Georgiou, I. Evolution of Minimally Invasive and Non-Invasive Preimplantation Genetic Testing: An Overview. J. Clin. Med. 2024, 13, 2160. https://doi.org/10.3390/jcm13082160
Moustakli E, Zikopoulos A, Skentou C, Bouba I, Dafopoulos K, Georgiou I. Evolution of Minimally Invasive and Non-Invasive Preimplantation Genetic Testing: An Overview. Journal of Clinical Medicine. 2024; 13(8):2160. https://doi.org/10.3390/jcm13082160
Chicago/Turabian StyleMoustakli, Efthalia, Athanasios Zikopoulos, Charikleia Skentou, Ioanna Bouba, Konstantinos Dafopoulos, and Ioannis Georgiou. 2024. "Evolution of Minimally Invasive and Non-Invasive Preimplantation Genetic Testing: An Overview" Journal of Clinical Medicine 13, no. 8: 2160. https://doi.org/10.3390/jcm13082160
APA StyleMoustakli, E., Zikopoulos, A., Skentou, C., Bouba, I., Dafopoulos, K., & Georgiou, I. (2024). Evolution of Minimally Invasive and Non-Invasive Preimplantation Genetic Testing: An Overview. Journal of Clinical Medicine, 13(8), 2160. https://doi.org/10.3390/jcm13082160