Identification and Expression of Neurotrophin-6 in the Brain of Nothobranchius furzeri: One More Piece in Neurotrophin Research

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Experimental Section
2.1. Protocols
2.2. Animals and Tissue Preparation
2.3. RNA Isolation and cDNA Synthesis
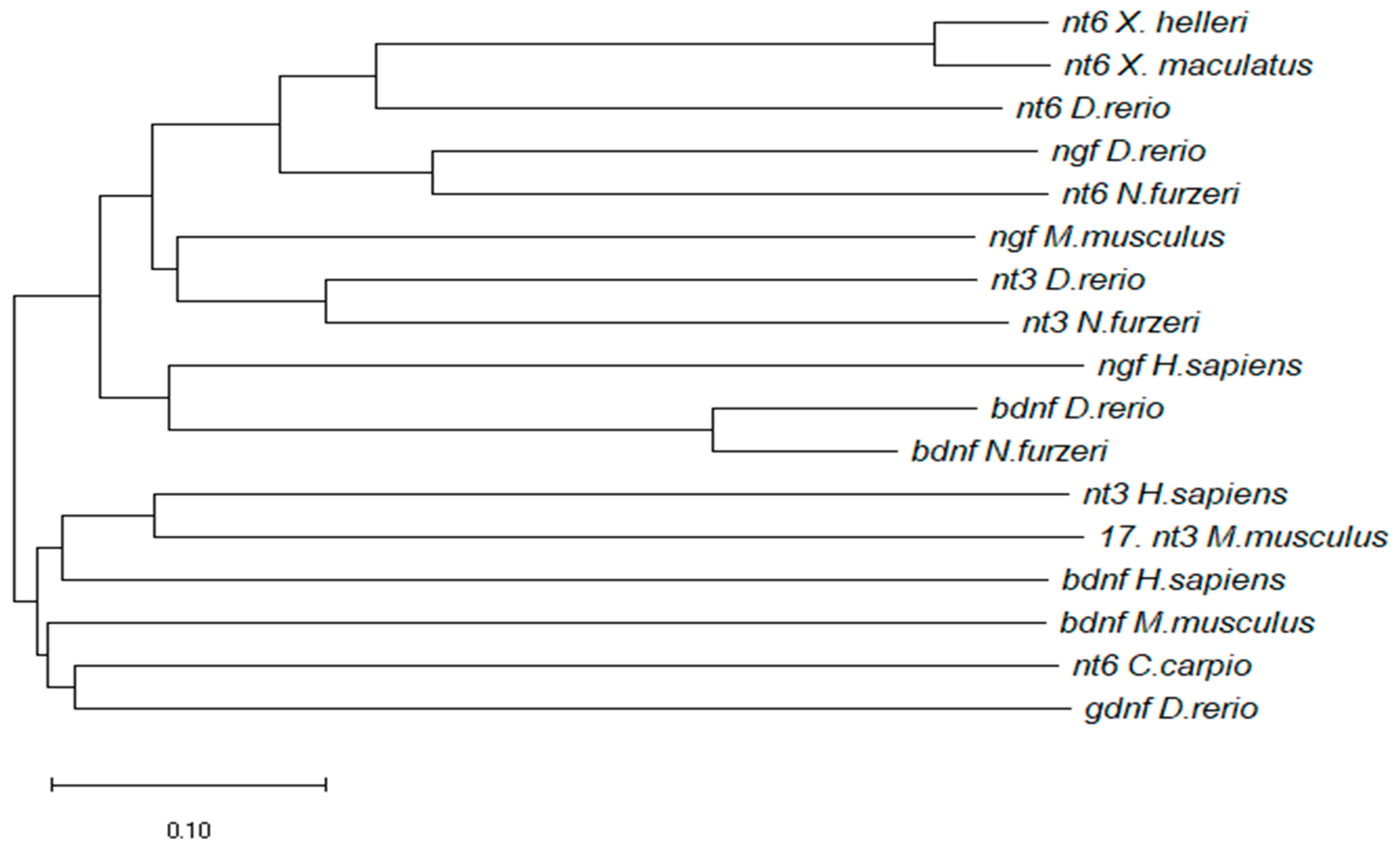
2.4. Phylogenetics Analysis
2.5. Quantitative Real Time-PCR
2.6. Statistical Analysis
2.7. Probe Design
2.8. Riboprobe Synthesis
2.9. LNA Probe Synthesis
2.10. Fluorescence In Situ Hybridization
2.11. Combined In Situ Hybridization and Immunohistochemistry
2.12. Microscopy
3. Results
3.1. Molecular Characterization of NfuNT-6
3.2. Expression Studies
3.2.1. NfuNT-6 mRNA Expression in Young versus Old Animals
3.2.2. Neuroanatomical Expression of NT-6 mRNA
3.2.3. NT-6 mRNA Expression in Mature Neurons
3.2.4. Neuroanatomical Distribution of NfuNT-6 mRNA in the Adult N. furzeri
Forebrain
Midbrain
Hindbrain
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cellerino, A.; Valenzano, D.R.; Reichard, M. From the bush to the bench: The annual Nothobranchius fishes as a new model system in biology. Biol. Rev. Camb. Philos. Soc. 2016, 91, 511–533. [Google Scholar] [CrossRef]
- D’Angelo, L.; Lossi, L.; Merighi, A.; de Girolamo, P. Anatomical features for the adequate choice of experimental animal models in biomedicine: I. Fishes. Ann. Anat. 2016, 205, 75–84. [Google Scholar] [CrossRef]
- Kim, Y.; Nam, H.G.; Valenzano, D.R. The short-lived African turquoise killifish: An emerging experimental model for ageing. Dis. Model Mech. 2016, 2, 115–129. [Google Scholar] [CrossRef]
- Genade, T.; Benedetti, M.; Terzibasi, E.; Roncaglia, P.; Valenzano, D.R.; Cattaneo, A.; Cellerino, A. Annual fishes of the genus Nothobranchius as a model system for aging research. Anat. Soc. 2005, 4, 223–233. [Google Scholar] [CrossRef]
- Blazek, R.; Polacik, M.; Reichard, M. Rapid growth, early maturation and short generation time in African annual fishes. EvoDevo 2013, 4, 24. [Google Scholar] [CrossRef]
- Valenzano, D.R.; Terzibasi, E.; Genade, T.; Cattaneo, A.; Domenici, L.; Cellerino, A. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short lived vertebrate. Curr. Biol 2006, 16, 296–300. [Google Scholar] [CrossRef]
- Di Cicco, E.; Terzibasi Tozzini, E.; Rossi, G.; Cellerino, A. The short-lived annual fish Nothobranchius furzeri shows a typical teleost aging process reinforced by high incidence of age-dependent neoplasias. Exp. Gerontol. 2011, 46, 249–256. [Google Scholar] [CrossRef]
- Hartmann, N.; Reichwald, K.; Wittig, I.; Dröse, S.; Schmeisser, S.; Lück, C.; Hahn, C.; Graf, M.; Gausmann, U.; Terzibasi, E.; et al. Mitochondrial DNA copy number and function decrease with age in the short-lived fish Nothobranchius furzeri. Anat. Soc. 2011, 10, 824–831. [Google Scholar]
- Terzibasi Tozzini, E.; Baumgart, E.; Battistoni, G.; Cellerino, A. Adult neurogenesis in the short-lived teleost Nothobranchius furzeri: localization of neurogenic niches, molecular characterization and effects of aging. Anat. Soc. 2012, 11, 241–251. [Google Scholar]
- Valenzano, D.R.; Benayoun, B.A.; Singh, P.P.; Zhang, E.; Etter, P.D.; Hu, C.K.; Clément-Ziza, M.; Wilemesen, D.; Cui, R.; Harel, I.; et al. The African Turquoise Killifish Genome Provides Insights into Evolution and Genetic Architecture of Lifespan. Cell Press 2015, 163, 1539–1554. [Google Scholar] [CrossRef]
- Wang, W.X.; Wilfred, B.R.; Madathil, S.K.; Tang, G.; Hu, Y.; Dimajuga, J.; Stromberg, A.J.; Huang, Q.; Saatman, K.E.; Nelson, P.T. miR-107 Regulates Granulin/Progranulin with Implications for Traumatic Brain Injury and Neurodegenerative Disease. Am. J. Pathol. 2010, 177, 334–345. [Google Scholar] [CrossRef]
- Cohen, S.; Levi-Montalcini, R.; Hamburger, V. A nerve growth-stimulating factor isolated from sarcom as 37 and 180. Proc. Natl. Acad. Sci. USA 1954, 40, 1014–1018. [Google Scholar] [CrossRef]
- Barde, Y.A.; Edgar, D.; Thoenen, H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982, 1, 549–553. [Google Scholar] [CrossRef]
- Leibrock, J.; Lottspeich, F.; Hohn, A.; Hofer, M.; Hengerer, B.; Masiakowski, P.; Thoenen, H.; Barde, Y.A. Molecular cloning and expression of brain-derived neurotrophic factor. Nature 1989, 341, 149–152. [Google Scholar] [CrossRef]
- Lewin, G.R.; Barde, Y.A. Physiology of the neurotrophins. Annu. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef]
- Lanave, C.; Colangelo, A.M.; Saccone, C.; Alberghina, L. Molecular evolution of the neurotrophin family members and their Trk receptors. Gene 2009, 394, 1–12. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef]
- Skaper, S.D. Compartmented chambers for studying neurotrophic factor action. Methods Mol. Biol. 2006, 846, 213–222. [Google Scholar]
- Heinrich, G.; Lum, T. Fish neurotrophins and Trk receptors. Int. J. Dev. Neurosci. 2000, 18, 1–27. [Google Scholar] [CrossRef]
- Götz, R.; Köster, R.; Winkler, C.; Raulf, F.; Lottspeich, F.; Schartl, M.; Thoenen, H. Neurotrophin-6 is a new member of the nerve growth factor family. Nature 1994, 372, 266–269. [Google Scholar] [CrossRef]
- Nilsson, A.S.; Fainzilber, M.; Falck, P.; Ibáñez, C.F. Neurotrophin-7: A novel member of the neurotrophin family from the zebrafish. FEBS Lett. 1998, 424, 285–290. [Google Scholar] [CrossRef]
- Lai, K.O.; Fu, W.Y.; Ip, F.C.; Ip, N.Y. Cloning and expression of a novel neurotrophin, NT-7, from carp. Mol. Cell Neurosci. 1998, 11, 64–76. [Google Scholar] [CrossRef]
- Dethleffsen, K.; Heinrich, G.; Lauth, M.; Knapik, E.W.; Meyer, M. Insert-containing neurotrophins in teleost fish and their relationship to nerve growth factor. Mol. Cell Neurosci. 2003, 24, 380–394. [Google Scholar] [CrossRef]
- Nittoli, V.; Sepe, R.M.; Coppola, U.; D’Agostino, Y.; De Felice, E.; Palladino, A.; Vassalli, Q.A.; Locascio, A.; Ristoratore, F.; Spagnuolo, A.; et al. A comprehensive analysis of neurotrophins and neurotrophin tyrosine kinase receptors expression during development of zebrafish. J. Comp. Neurol. 2018, 526, 1057–1072. [Google Scholar] [CrossRef]
- Terzibasi, E.; Valenzano, D.R.; Benedetti, M.; Roncaglia, P.; Cattaneo, A.; Domenici, L.; Cellerino, A. Large differences in aging phenotype between strains of the short-lived annual fish Nothobranchius furzeri. PLoS ONE 2008, 3, e3866. [Google Scholar] [CrossRef]
- Baumgart, M.; Groth, M.; Priebe, S.; Appelt, J.; Guthke, R.; Platzer, M.; Cellerino, A. Age-dependent regulation of tumor-related microRNA in the brain of the annual fish Nothobranchius furzeri. Mech. Ageing Dev. 2012, 133, 226–233. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef]
- Thomsen, R.; Nielsen, P.S.; Jensen, T.H. Dramatically improved RNA in situ hybridization signals using LNA-modified probes. RNA 2005, 11, 1745–1748. [Google Scholar] [CrossRef]
- D’Angelo, L. Brain atlas of an emerging teleostean model: Nothobranchius furzeri. Anat. Rec. (Hoboken) 2013, 296, 681–691. [Google Scholar]
- Petzold, A.; Reichwald, K.; Groth, M.; Taudien, S.; Hartmann, N.; Priebe, S. The transcript catalogue of the short-lived fish Nothobranchius furzeri provides insights into age-dependent changes of mRNA levels. BMC Genom. 2013, 14, 185. [Google Scholar] [CrossRef]
- D’Angelo, L.; de Girolamo, P.; Cellerino, A.; Tozzini, E.T.; Varricchio, E.; Castaldo, L.; Lucini, C. Immunolocalization of S100-like protein in the brain of an emerging model organism: Nothobranchius furzeri. Microsc. Res Tech. 2012, 75, 441–447. [Google Scholar] [CrossRef]
- Zupanc, G.K.; Hinsch, K.; Gage, F.H. Proliferation, migration, neuronal differentiation, and long-term survival of new cells in the adult zebrafish brain. J. Comp. Neurol. 2005, 488, 290–319. [Google Scholar] [CrossRef]
- Pellegrini, E.; Mouriec, K.; Anglade, I.; Menuet, A.; Le Page, Y.; Gueguen, M.M.; Marmignon, M.H.; Brion, F.; Pakdel, F.; Kah, O. Identification of aromatase-positive radial glial cells as progenitor cells in the ventricular layer of the forebrain in zebrafish. J. Comp. Neurol. 2007, 501, 150–167. [Google Scholar] [CrossRef]
- Kroehne, V.; Freudenreich, D.; Hans, S.; Kaslin, J.; Brand, M. Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors. Development 2011, 138, 4831–4841. [Google Scholar] [CrossRef]
- Wohl, S.G.; Reh, T.A. miR-124-9-9* potentiates Ascl1-induced reprogramming of cultured Müller glia. Glia 2016, 64, 743–762. [Google Scholar] [CrossRef]
- Olivera-Pasilio, V.; Lasserre, M.; Castellò, M.E. Cell Proliferation, Migration, and Neurogenesis in the Adult Brain of the Pulse Type Weakly Electric Fish, Gymnotus omarorum. Front. Neurosci. 2017, 11, 437. [Google Scholar] [CrossRef]
- Cacialli, P.; D’angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.M.; Pellegrini, E.; Lucini, C. Neuronal expression of brain derived neurotrophic factor in the injured telencephalon of adult zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef]
- Iulita, F.M.; Cuello, C.A. The NGF Metabolic Pathway in the CNS and its Dysregulation in Down Syndrome and Alzheimer’s Disease. Curr. Alzheimer Res. 2016, 13, 53–67. [Google Scholar] [CrossRef]
- Peng, S.; Wuu, J.; Mufson, E.J.; Fahnestock, M. Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer’s disease. J. Neurochem. 2005, 93, 1412–1421. [Google Scholar] [CrossRef]
- Hallböök, F.; Lundin, L.G.; Kullander, K. Lampetra fluviatilis neurotrophin homolog, descendant of a neurotrophin ancestor, discloses the early molecular evolution of neurotrophins in the vertebrate subphylum. J. Neurosci. 1999, 18, 8700–8711. [Google Scholar] [CrossRef]
- Hallböök, F.; Wilson, K.; Thorndyke, M.; Olinski, R.P. Formation and evolution of the chordate neurotrophin and Trk receptor genes. Brain Behav. Evol. 2006, 68, 133–144. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef]
- Válóczi, A.; Hornyik, C.; Varga, N.; Burgyán, J.; Kauppinen, S.; Havelda, Z. Sensitive and specific detection of microRNAs by northern blot analysis using LNA-modified oligonucleotide probes. Nucleic Acids Res. 2004, 32, e175. [Google Scholar] [CrossRef]
- Kloosterman, W.P.; Wienhold, E.; de Bruijn, E.; Kauppinen, S.; Plasterk, R.H. In situ detection of miRNAs in animal embryos using LNA-modified oligonucleotide probes. Nat. Methods 2006, 3, 27–29. [Google Scholar] [CrossRef]
- Darnell, D.K.; Stanislaw, S.; Kaur, S.; Antin, P.B. Whole mount in situ hybridization detection of mRNAs using short LNA containing DNA oligonucleotide probes. RNA 2010, 16, 632–637. [Google Scholar] [CrossRef]
- Darnell, D.K.; Antin, P.B. LNA-based in situ hybridization detection of mRNAs in embryos. Methods Mol. Biol. 2014, 1211, 69–76. [Google Scholar]
- McTigue, P.M.; Peterson, R.J.; Kahn, J.D. Sequence-dependent thermodynamic parameters for locked nucleic acid (LNA)-DNA duplex formation. Biochem 2004, 43, 5388–5405. [Google Scholar] [CrossRef]
- D’Angelo, L.; de Girolamo, P.; Lucini, C.; Terzibasi, E.T.; Baumgart, M.; Castaldo, L.; Cellerino, A. Brain-derived neurotrophic factor: mRNA expression and protein distribution in the brain of the teleost Nothobranchius furzeri. J. Comp. Neurol. 2014, 522, 100–430. [Google Scholar]
- D’Angelo, L.; Castaldo, L.; Cellerino, A.; de Girolamo, P.; Lucini, C. Nerve growth factor in the adult brain of a teleostean model for aging research: Nothobranchius furzeri. Ann. Anat. 2014, 196, 183–191. [Google Scholar] [CrossRef]
- D’Angelo, L.; Avallone, L.; Cellerino, A.; de Girolamo, P.; Paolucci, M.; Varricchio, E.; Lucini, C. Neurotrophin-4 in the brain of adult Nothobranchius furzeri. Ann. Anat. 2016, 207, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Dalton, V.S.; Borich, S.M.; Murphy, P.; Roberts, B.L. Brain-derived neurotrophic factor mRNA expression in the brain of the teleost fish, Anguilla anguilla, the European Eel. Brain Behav. Evol. 2009, 73, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Gueguen, M.M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF Expression in Larval and Adult Zebrafish Brain: Distribution and Cell Identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef]
- Cacialli, P.; Gatta, C.; D’Angelo, L.; Leggieri, A.; Palladino, A.; de Girolamo, P.; Pellegrini, E.; Lucini, C. Nerve growth factor is expressed and stored in central neurons of adult zebrafish. J. Anat. 2019. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target mRNA | 5′-Mod | Synthesis Sequence (5′–3′) | 3′-Mod |
|---|---|---|---|
| NT-6 | DIG | TTGTCTCCTGCTGTCCTGCTCTG | DIG |
| * mut NT-6 | DIG | TTGTCTCTCGCTGCTCTGCTTCG | DIG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leggieri, A.; Attanasio, C.; Palladino, A.; Cellerino, A.; Lucini, C.; Paolucci, M.; Terzibasi Tozzini, E.; de Girolamo, P.; D’Angelo, L. Identification and Expression of Neurotrophin-6 in the Brain of Nothobranchius furzeri: One More Piece in Neurotrophin Research. J. Clin. Med. 2019, 8, 595. https://doi.org/10.3390/jcm8050595
Leggieri A, Attanasio C, Palladino A, Cellerino A, Lucini C, Paolucci M, Terzibasi Tozzini E, de Girolamo P, D’Angelo L. Identification and Expression of Neurotrophin-6 in the Brain of Nothobranchius furzeri: One More Piece in Neurotrophin Research. Journal of Clinical Medicine. 2019; 8(5):595. https://doi.org/10.3390/jcm8050595
Chicago/Turabian StyleLeggieri, Adele, Chiara Attanasio, Antonio Palladino, Alessandro Cellerino, Carla Lucini, Marina Paolucci, Eva Terzibasi Tozzini, Paolo de Girolamo, and Livia D’Angelo. 2019. "Identification and Expression of Neurotrophin-6 in the Brain of Nothobranchius furzeri: One More Piece in Neurotrophin Research" Journal of Clinical Medicine 8, no. 5: 595. https://doi.org/10.3390/jcm8050595