Hydrogen Sulfide Affects the Root Development of Strawberry During Plug Transplant Production




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemical Treatments
2.2. Measurement of Root Development and Sampling
2.3. H2S and H2O2 Quantification
2.4. Measurements of Soluble Sugar, Starch, and Soluble Protein Contents
2.5. Determination of the Antioxidant Enzyme Activities
2.6. Statistical Analysis
3. Results
3.1. The Optimal Concentration of NaHS for Strawberry (Experiment 1)
3.2. Root Development Induced by H2S (Experiment 2)
3.3. Soluble Sugar, Starch, and Soluble Protein Content
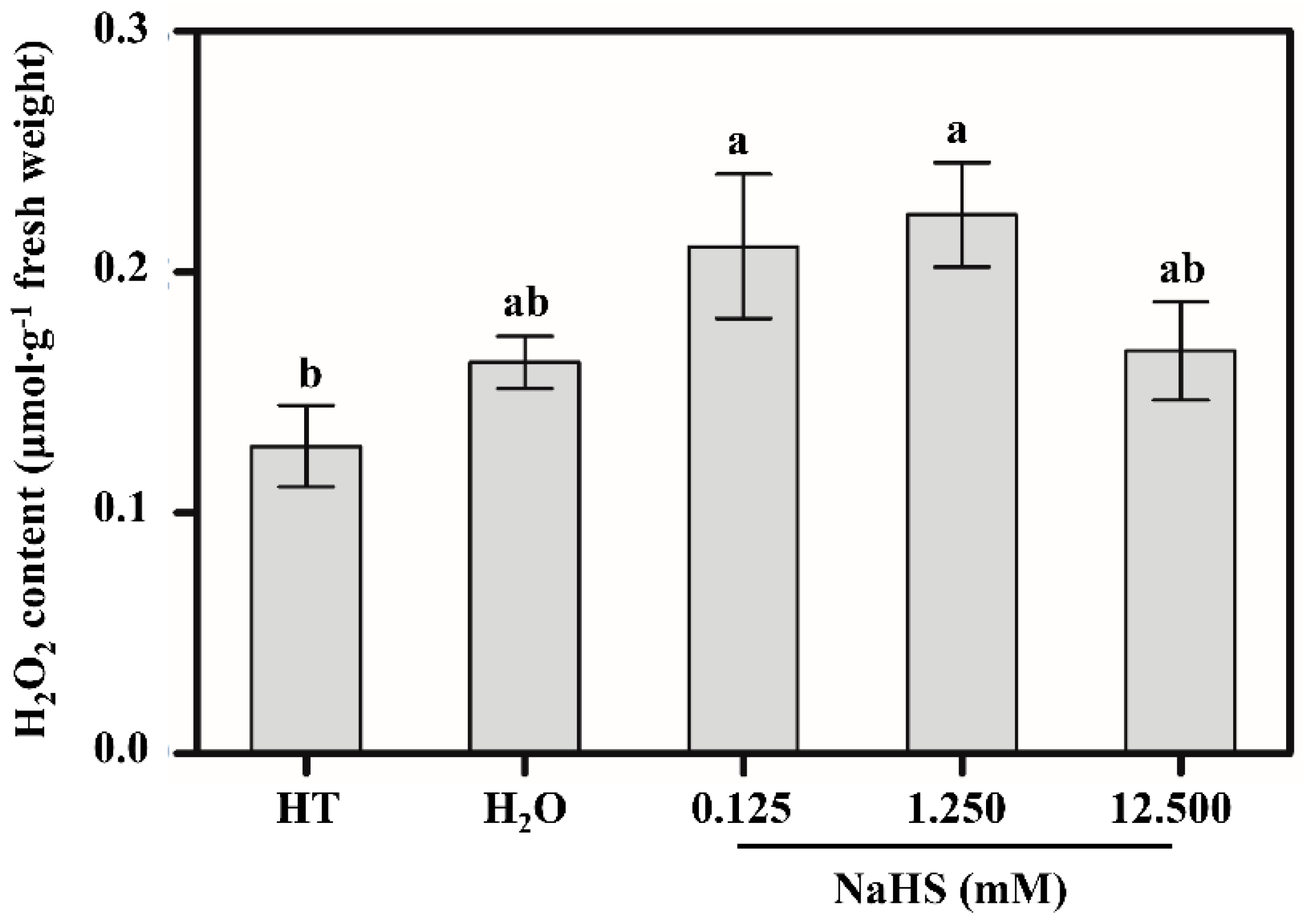
3.4. H2O2 Content
3.5. Antioxidant Enzyme Activities
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Durner, E.F.; Poling, E.B.; Maas, J.L. Recent advances in strawberry plug transplant technology. HortTechnology 2002, 12, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Giménez, G.; Andriolo, J.L.; Janisch, D.; Cocco, C.; Picio, M.D. Cell size in trays for the production of strawberry plug transplants. Pesqui. Agropecu. Bras. 2009, 44, 726–729. [Google Scholar] [CrossRef]
- Türkben, C. Propagation of strawberry plants in pots: Effect of runner order and rooting media. Int J. Biol. Sci. 2008, 2, 1–4. [Google Scholar]
- Hokanson, S.C.; Takeda, F.; Enns, J.M.; Black, B.L. Influence of plant storage duration on strawberry runner tip viability and field performance. HortScience 2004, 39, 1596–1600. [Google Scholar] [CrossRef]
- Saito, Y.; Imagawa, M.; Yabe, K.; Bantog, N.; Yamada, K.; Yamaki, S. Stimulation of rooting by exposing cuttings of runner plants to low temperatures to allow the raising of strawberry seedlings during summer. J. Jpn. Soc. Hortic. Sci. 2008, 77, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Massetani, F.; Savini, G.; Neri, D. Effect of rooting time, pot size and fertigation technique on strawberry plant architecture. J. Berry Res. 2014, 4, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Hu, Y.; Fan, T.; Li, J. Hydrogen sulfide modulates actin-dependent auxin transport via regulating ABPs results in changing of root development in Arabidopsis. Sci. Rep. 2015, 5, 8251. [Google Scholar] [CrossRef] [Green Version]
- Fang, T.; Cao, Z.; Li, J.; Shen, W.; Huang, L. Auxin-induced hydrogen sulfide generation is involved in lateral root formation in tomato. Plant Physiol. Bioch. 2014, 76, 44–51. [Google Scholar] [CrossRef]
- Fang, T.; Li, J.; Cao, Z.; Chen, M.; Shen, W.; Huang, L. Heme oxygenase-1 is involved in sodium hydrosulfide-induced lateral root formation in tomato seedlings. Plant Cell Rep. 2014, 33, 969–978. [Google Scholar] [CrossRef]
- Mei, Y.; Chen, H.; Shen, W.; Shen, W.; Huang, L. Hydrogen peroxide is involved in hydrogen sulfide-induced lateral root formation in tomato seedlings. BMC Plant Biol. 2017, 17, 162. [Google Scholar] [CrossRef]
- Kou, N.; Xiang, Z.; Cui, W.; Li, L.; Shen, W. Hydrogen sulfide acts downstream of methane to induce cucumber adventitious root development. J. Plant Physiol. 2018, 228, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.Y.; Hu, S.L.; Wu, J.; Li, Y.H.; Zheng, J.L.; Wei, Z.J.; Liu, J.; Wang, H.L.; Liu, Y.S.; Zhang, H. Hydrogen sulfide prolongs postharvest shelf life of strawberry and plays an antioxidative role in fruits. J. Agric. Food Chem. 2012, 60, 8684–8693. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Jingying, S.; Liqin, Z.; Changle, L.; Qingguo, W. Cooperative effects of hydrogen sulfide and nitric oxide on delaying softening and decay of strawberry. Int. J. Agric. Biol. Eng. 2014, 7, 114–122. [Google Scholar]
- Christou, A.; Manganaris, G.A.; Papadopoulos, I.; Fotopoulos, V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defense pathways. J. Exp. Bot. 2013, 64, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fotopoulos, V.; Christou, A.; Antoniou, C.; Manganaris, G. Hydrogen sulphide: A versatile tool for the regulation of growth and defense responses in horticultural crops. J. Hort. Sci. Biotechnol. 2015, 90, 227–234. [Google Scholar] [CrossRef]
- Husen, A.; Pal, M. Metabolic changes during adventitious root primordium development in Tectona grandis Linn. f. (teak) cuttings as affected by age of donor plants and auxin (IBA and NAA) treatment. New For. 2007, 33, 309–323. [Google Scholar] [CrossRef]
- Husen, A. Stock-plant etiolation causes drifts in total soluble sugars and anthraquinones, and promotes adventitious root formation in teak (Tectona grandis L. f.) coppice shoots. Plant Growth Regul. 2008, 54, 13–21. [Google Scholar] [CrossRef]
- Reza-Shahsavar, A. Endogenous soluble sugars, starch contents and phenolic compounds in easy-and difficult-to-root olive cuttings. J. Biol. Environ. Sci. 2010, 4, 83–86. [Google Scholar]
- Haissig, B.E. Metabolic processes in adventitious rooting of cuttings. In New Root Formation in Plants and Cuttings; Springer: Dordrecht, The Netherlands, 1986; pp. 141–189. [Google Scholar]
- Mohan, R.; Chui, E.A.; Biasi, L.A.; Soccol, C.R. Alternative in vitro propagation: Use of sugarcane bagasse as a low cost support material during rooting stage of strawberry Cv. Dover. Braz. Arch. Biol. Technol. 2005, 48, 37–42. [Google Scholar] [CrossRef]
- Chen, J.; Shang, Y.T.; Wang, W.H.; Chen, X.Y.; He, E.M.; Zheng, H.L.; Shangguan, Z. Hydrogen sulfide-mediated polyamines and sugar changes are involved in hydrogen sulfide-induced drought tolerance in Spinacia oleracea seedlings. Front. Plant Sci. 2016, 7, 1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.W.; Leng, Y.; Feng, L.; Zeng, X.Y. Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean (Vigna radiata (L.) Wilczek) seedlings under cadmium stress. Environ. Sci. Pollut. Res. 2014, 21, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Kose, C.; Erdal, S.; Kaya, O.; Atici, O. Comparative evaluation of oxidative enzyme activities during adventitious rooting in the cuttings of grapevine rootstocks. J. Sci. Food Agric. 2011, 91, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Elmongy, M.S.; Zhou, H.; Cao, Y.; Liu, B.; Xia, Y. The effect of humic acid on endogenous hormone levels and antioxidant enzyme activity during in vitro rooting of evergreen azalea. Sci. Hortic. 2018, 227, 234–243. [Google Scholar] [CrossRef]
- Li, S.W.; Zeng, X.Y.; Leng, Y.; Feng, L.; Kang, X.H. Indole-3-butyric acid mediates antioxidative defense systems to promote adventitious rooting in mung bean seedlings under cadmium and drought stresses. Ecotox. Environ. Saf. 2018, 161, 332–341. [Google Scholar] [CrossRef]
- Veal, E.; Day, A. Hydrogen Peroxide as a Signaling Molecule; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2011. [Google Scholar]
- Yang, W.; Zhu, C.; Ma, X.; Li, G.; Gan, L.; Ng, D.; Xia, K. Hydrogen peroxide is a second messenger in the salicylic acid-triggered adventitious rooting process in mung bean seedlings. PLoS ONE 2013, 8, e84580. [Google Scholar] [CrossRef] [Green Version]
- Li, S.W.; Xue, L.; Xu, S.; Feng, H.; An, L. IBA-induced changes in antioxidant enzymes during adventitious rooting in mung bean seedlings: The role of H2O2. Environ. Exp. Bot. 2009, 66, 442–450. [Google Scholar] [CrossRef]
- Li, C.; Bian, B.; Gong, T.; Liao, W. Comparative proteomic analysis of key proteins during abscisic acid-hydrogen peroxide-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Physiol. 2018, 229, 185–194. [Google Scholar] [CrossRef]
- Zhao, Y.; Biggs, T.D.; Xian, M. Hydrogen sulfide (H2S) releasing agents: Chemistry and biological applications. Chem. Commun. 2014, 50, 11788–11805. [Google Scholar] [CrossRef] [Green Version]
- Nashef, A.S.; Osuga, D.T.; Feeney, R.E. Determination of hydrogen sulfide with 5,5’-dithiobis-(2-nitrobenzoic acid), n-ethylmaleimide, and parachloromercuribenzoate. Anal. Biochem. 1977, 79, 394–405. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Wang, S.; Zhang, P.; Zhu, F.; Ren, X.; Liu, C.; Zhang, X. On the role of physiological substances, abscisic acid and its biosynthetic genes in seed maturation and dormancy of tree peony (Paeonia ostii ‘Feng Dan’). Sci. Hortic. 2015, 182, 92–101. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Arum, L.S.; Ko, C.H.; Muneer, S.; Jeong, B.R. Blue LED light enhances growth phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Jin, Z.; Pei, Y. Physiological implications of hydrogen sulfide in plants: Pleasant exploration behind its unpleasant odour. Oxid. Med. Cell. Longev. 2015, 2015, 397502. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Li, M.Y.; Cui, W.T.; Lu, W.; Shen, W.B. Haem oxygenase-1 is involved in hydrogen sulfide-induced cucumber adventitious root formation. J. Plant Growth Regul. 2012, 31, 519–528. [Google Scholar] [CrossRef]
- Sairanen, I.; Novak, O.; Pencik, A.; Ikeda, Y.; Jones, B.; Sandberg, G.; Ljung, K. Soluble carbohydrates regulate auxin biosynthesis via PIF proteins in Arabidopsis. Plant Cell 2012, 24, 4907–4916. [Google Scholar] [CrossRef] [Green Version]
- Buettner, G.R.; Ng, C.F.; Wang, M.; Rodgers, V.G.J.; Schafer, F.Q. A new paradigm: Manganese superoxide dismutase influences the production of H2O2 in cells and thereby their biological state. Free Radical Biol. Med. 2006, 41, 1338–1350. [Google Scholar] [CrossRef] [Green Version]
- Aghdam, M.S.; Mahmoudi, R.; Razavi, F.; Rabiei, V.; Soleimani, A. Hydrogen sulfide treatment confers chilling tolerance in hawthorn fruit during cold storage by triggering endogenous H2S accumulation, enhancing antioxidant enzymes activity and promoting phenols accumulation. Sci. Hortic. 2018, 238, 264–271. [Google Scholar] [CrossRef]
- Min, Y.; Qin, B.P.; Ping, W.; Li, M.L.; Chen, L.L.; Chen, L.T.; Sun, A.Q.; Wang, Z.L.; Yin, Y.P. Foliar application of sodium hydrosulfide (NaHS), a hydrogen sulfide (H2S) donor, can protect seedlings against heat stress in wheat (Triticum aestivum L.). J. Integr. Agric. 2016, 15, 2745–2758. [Google Scholar]
- Da Silva, C.J.; Fontes, E.P.B.; Modolo, L.V. Salinity-induced accumulation of endogenous H2S and NO is associated with modulation of the antioxidant and redox defense systems in Nicotiana tabacum L. cv. Havana. Plant Sci. 2017, 256, 148–159. [Google Scholar] [CrossRef]
- Li, Z.G.; Gong, M.; Xie, H.; Yang, L.; Li, J. Hydrogen sulfide donor sodium hydrosulfide-induced heat tolerance in tobacco (Nicotiana tabacum L.) suspension cultured cells and involvement of Ca2+ and calmodulin. Plant Sci. 2012, 185, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Filippou, P.; Tanou, G.; Molassiotis, A.; Fotopoulos, V. Plant acclimation to environmental stress using priming agents. In Plant Acclimation to Environmental Stress; Springer: New York, NY, USA, 2013; pp. 1–27. [Google Scholar]
- He, H.; Li, Y.; He, L.F. The central role of hydrogen sulfide in plant responses to toxic metal stress. Ecotox. Environ. Saf. 2018, 157, 403–408. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, Y.; Liu, Y.; Kang, D.I.; Wei, H.; Jeong, B.R. Hydrogen Sulfide Affects the Root Development of Strawberry During Plug Transplant Production. Agriculture 2020, 10, 12. https://doi.org/10.3390/agriculture10010012
Hu J, Li Y, Liu Y, Kang DI, Wei H, Jeong BR. Hydrogen Sulfide Affects the Root Development of Strawberry During Plug Transplant Production. Agriculture. 2020; 10(1):12. https://doi.org/10.3390/agriculture10010012
Chicago/Turabian StyleHu, Jiangtao, Yali Li, Ya Liu, Dong Il Kang, Hao Wei, and Byoung Ryong Jeong. 2020. "Hydrogen Sulfide Affects the Root Development of Strawberry During Plug Transplant Production" Agriculture 10, no. 1: 12. https://doi.org/10.3390/agriculture10010012