1. Introduction
Cover cropping is a multifunctional agroecological practice widely used in organic farming, aiming at higher crop diversity, reduced erosion and nitrate leaching [
1]. Cover crops have the potential to reduce the yield gap between organic and conventional farming or between conservation agriculture and intensive tillage owing to reduction of weed pressure and additional nitrogen provided by legume cover crops [
2]. Moreover, cover crops are being discussed as a measure to mitigate climate change due to carbon sequestration, reduced fertilizer use after legume cover crops and change in surface albedo [
3].
Except for weed suppression [
4], the main benefits of cover crops are associated with the roots: prevention of nitrate leaching due to nutrient uptake [
5], reduction of soil erosion [
6], organic matter input to enhance soil organic carbon content [
7,
8] and improvement of soil structure [
9,
10]. Thus, it is important to know more about the root intensity, distribution and dynamics of cover crops in organic cropping systems.
Poeplau and Don [
7] as well as Jian et al. [
8] showed the potential of cover crops for carbon sequestration in arable soils. As roots contribute more effectively to the carbon pool than carbon input from above ground plant material [
11], cover crop roots might have an important effect on carbon sequestration. Topsoil resistance against erosion can be increased due to a high root length density (RLD) in upper soil layers, as it was shown for a grass mixture by Baets et al. [
6]. Moreover, surface runoff can be reduced by cover crops. Cover crops with coarse root axis, especially legumes, enhance hydraulic conductivity and thus diminish surface runoff [
12]. Hence, soil physical properties can be managed by cover crop roots. Bodner et al. [
13] showed with a range of different cover crops that coarse roots increase macropores whereas fine roots increase micropores. With their dense and ramified root system, the winter hardy cover crops annual ryegrass and rye increased aggregate stability in the topsoil [
9].
On the other hand, roots of cover crops are important for nutrient uptake and therefore for nutrient saving in arable cropping systems, especially of highly mobile nutrients as nitrate, during winter. To highlight this property, they are also called catch crops. Sainju et al. [
14] reported that higher RLD of cover crops correlated positively with shoot nitrogen uptake and negatively with nitrate concentration in soil. In contrast, Herrera et al. [
15] found no relationship between nitrogen uptake and nitrate leaching for three different cover crops. However, they showed a relationship between rapid root establishment and reduction of nitrate leaching. In a study with three cover crops differentiated by rooting depth, Italian ryegrass, winter rye and oil radish, Kristensen and Thorup-Kristensen [
16] described a linear relationship between nitrogen uptake and root intensity or root frequency. The deep rooting cover crops resulted in higher N uptake and lower residual soil nitrate. Similarly, Sapkota et al. [
17] recorded that perennial ryegrass with a shallow but dense root system caused high nitrogen depletion in upper layers while the deep rooting oil radish depleted more mineral soil nitrogen in the total soil layer. Thus, deep rooting of cover crops is important to prevent nitrogen losses from the subsoil to deeper layers and to the groundwater. With a focus on deep rooting, Thorup-Kristensen [
18] showed that residual subsoil nitrate is well correlated with root intensity and rooting depth, but not with aboveground measurements. For deep-rooted undersown cover crops it was shown that subsoil nitrate reduction was correlated with root growth in the subsoil, but not with root growth in the topsoil [
19]. A high rooting depth was found for non-legume dicot species; temperature sum (growing degree-days) needed for these crops to reach one meter soil depth was much smaller than for grass species [
18].
Consequently, cover crops with large root systems are desirable. For some beneficial effects such as improvement of topsoil structure with consequent reduction of surface runoff and erosion, intensive rooting in the topsoil is crucial. For other desired effects such as depletion of nitrate prone to be leached to the groundwater from deeper soil layers, deep rooting is the most important trait. This means high RLD in topsoil and deep rooting into the subsoil are efficient to explore the soil volume more completely, reduce erosion, improve soil structure and aggregate stability and to store organic carbon in arable soils. Thus, for optimizing cropping systems in an informed and targeted way, detailed knowledge on the root systems and their vertical distribution for different cover crops is indispensable.
Wendling et al. [
20] gave detailed information on root traits of a wide range of cover crop species. However, root traits were analysed only until 50 cm soil depth and no function was used to illustrate the distribution of RLD by soil depth. Some information on RLD profiles can be found in Bodner et al. [
21] for phacelia, rye, mustard and hairy vetch, and in Vos et al. [
22] for winter rye and forage rape. Both studies used an exponential function for distribution of RLD by depth. However, they only examined RLD until 40 cm and 60 cm soil depth, respectively. To describe the exploration of the total soil volume, the cumulative root distribution of the RLD of different cover crops is of interest. In a meta-study, Fan et al. [
23] modified a logistic dose-response curve for cumulative root distribution by depth for agricultural crops and showed different root distributions for different types of agricultural crops. However, cover crops were not included.
In the otherwise densely packed subsoil, tubular continuous biopores formed by plant roots or earthworms are sites of preferential flow for water and solutes and may thus potentially increase nitrate leaching, as shown in a microcosm study [
24]. However, in the field, nitrate leaching in biopores may be counteracted by root growth, since these pores also provide paths of preferential root growth and improved nutrient and water uptake in the subsoil [
25]. RLD of annual crops increased in the deeper subsoil when grown after a biopore density-increasing taprooted precrop compared to a fibrous rooted precrop [
26,
27]. Roots use biopores to grow through compacted soil layers, and in less compacted soil layers roots can exit biopores and re-enter bulk soil [
28]. In a study using in situ endoscopy to evaluate root growth of barley and oilseed rape in biopores, it was shown that in 85% of the evaluated biopores both barley and oilseed rape established root-soil contact on the biopore wall. However, homorhizous barley had predominantly thin vertical or ingrowing roots contacting the pore wall, while allorhizous oilseed rape use mainly lateral roots to contact the pore wall [
29]. Thus, it can be assumed that biopores are also used by cover crop species, possibly to different extents depending on their root system and rooting depth.
In this study we compared the RLD of seven different cover crops down to a depth of 1 m. We investigated allorhizous cover crops with a taproot-dominated root system (blue lupin, oil radish, winter turnip rape, phacelia), homorhizous cover crops with shoot-borne roots (winter rye, bristle oat) and crimson clover as an intermediate type which has a taproot with strong lateral roots. RLD was analysed with the profile wall method [
30]. We differentiated between root length in large-sized biopores and root length in bulk soil and compared RLD before and after winter.
The objective of the present study was to evaluate rooting patterns of different cover crops. We hypothesized that (i) topsoil RLD is higher for fibrous rooted crops, (ii) subsoil RLD is higher for taprooted than fibrous rooted crops, (iii) 50% of the cumulative root length is reached in lower soil depths for fibrous rooted compared to taprooted crops, (iv) non-winter hardy cover crops lose RLD during winter compared to winter hardy cover crops, and (v) allorhizous cover crop use large-sized biopores to a greater extent than homorhizous cover crops.
4. Discussion
4.1. Characterization of Cover Crop Rooting: Root Length Density and Rooting Depth
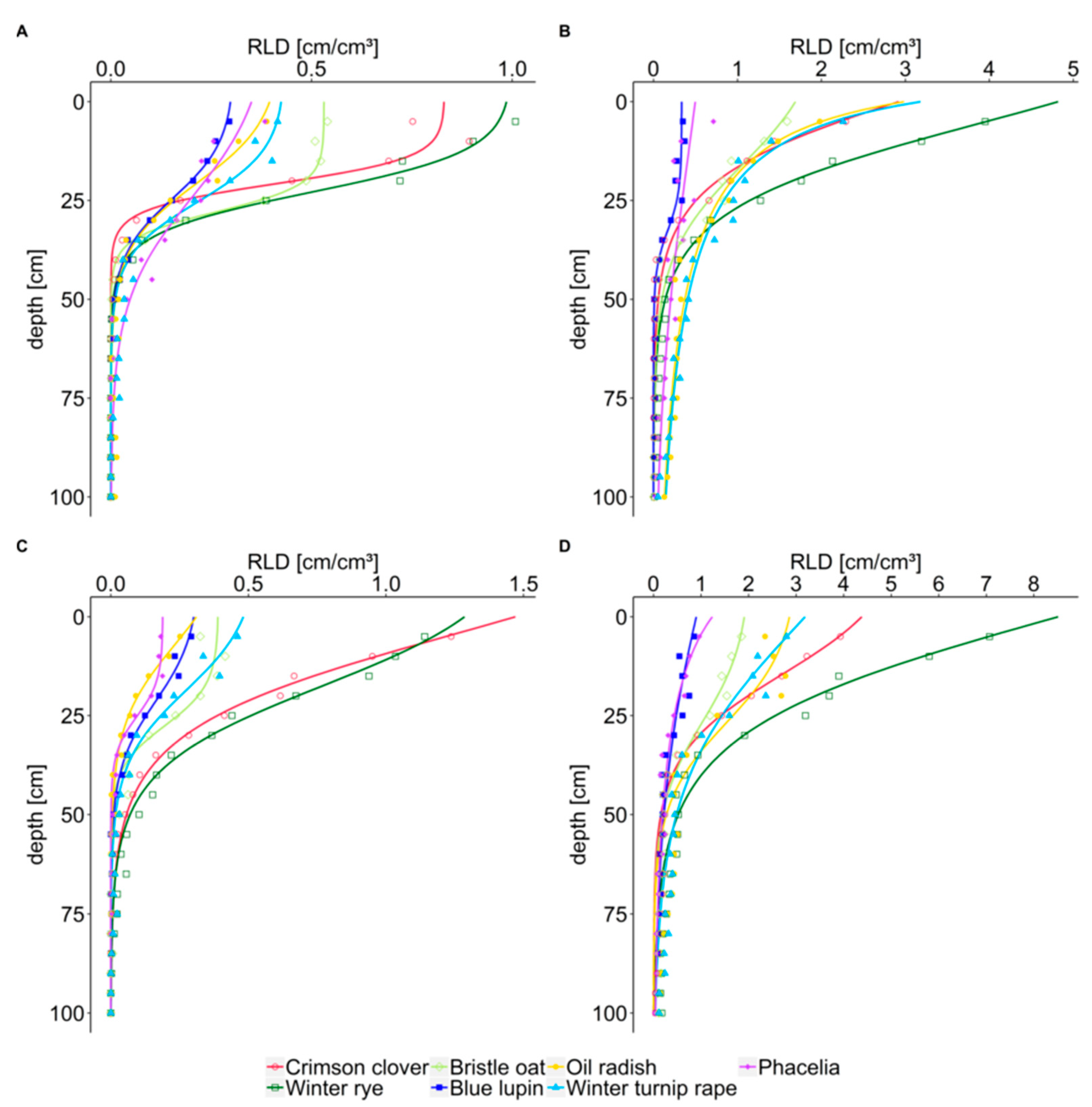
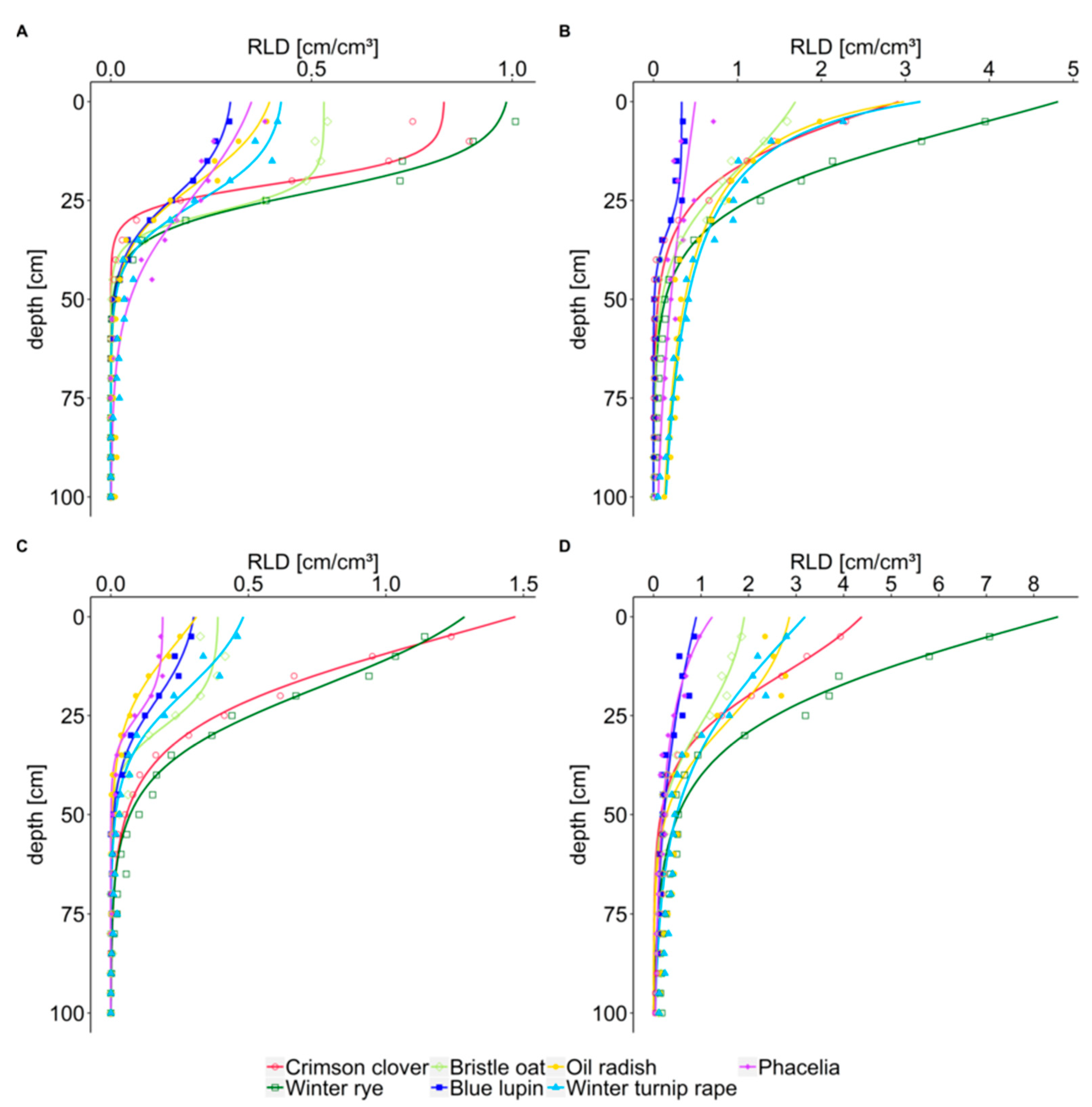
In our study, crimson clover, the intermediate type between taprooted and fibrous rooted crops, rooted intensively in the topsoil, reaching up to 3.9 cm cm
−3 in 5 cm depth. These values are in accordance with Sainju et al. [
14], who measured an RLD of up to 10 cm cm
−3 for crimson clover between 0–30 cm soil depth, which was significantly lower than RLD of rye.
In the topsoil, the homorhizous winter rye was the strongest rooting cover crop, increasing RLD over winter to more than 7.1 cm cm
−3 in 5 cm depth. In several other field studies, winter rye was also the crop that rooted most intensively [
21] and with higher RLD than for oilseed rape [
22]. However, Bodner et al. [
13] measured a significantly smaller RLD for winter rye than for oil radish and phacelia. This is contrary to our results with much higher RLD for winter rye compared to oil radish and phacelia in the surface near topsoil, and to the results of Vos et al. [
22].
For the second homorhizous crop in our study, bristle oat, Wendling et al. [
20] reported a higher RLD than for oil radish and phacelia, and bristle oat represented the highest RLD of all 20 tested cover crops. In their study, rye was not included. This shows the intensive rooting of
Poaceae. However, in our experiment, bristle oat was intermediate in RLD with up to 1.8 cm cm
−3 in the 5 cm depth, i.e., not as intensive as winter rye and similar to or lower than the brassica cover crops. Results of Bodner et al. [
33] reporting rather low total root length for bristle oat compared to other cover crops in a rhizobox experiment, support our findings that bristle oat is less intensively rooting than winter rye.
RLD of taprooted blue lupin was found to be low compared to all other crops, especially in the topsoil with up to 0.8 cm cm
−3 in 5 cm. This result of low RLD of blue lupin is confirmed by a comparison of blue lupin as a main crop compared to cereals [
34].
RLD of taprooted oil radish was intermediate in the topsoil with up to 2.8 cm cm
−3 in 5 cm depth, and high in the subsoil. Similarly, in the study by Wendling et al. [
20], RLD of oil radish was lower than that of fibrous rooted oats. However, in other cover crop comparisons, oil radish exhibited high RLD not only in the subsoil, but also in the topsoil [
13]. In a minirhizotron study, oil radish RLD outscored winter rye RLD in both topsoil and subsoil [
16]. Bodner et al. [
33] reported oil radish to be the species with the highest total root length among 10 cover crop species in a rhizobox experiment, in which the only homorhizous species included was bristle oats. Munkholm and Hansen [
35] determined a RLD of oil radish, which was slightly higher than that of homorhizous ryegrass in topsoil and subsoil. In the present study, RLD of taprooted winter turnip rape was very similar to that of oil radish, in both topsoil and subsoil. Wendling et al. [
20] tested a summer turnip rape and reported a RLD of 6.00 cm cm
−3 in the top 50 cm, which was also in the same range of oil radish (5.76 cm cm
−3).
RLD of taprooted phacelia was low in the topsoil compared to other crops, with up to 1.0 cm cm
−3 in 5 cm depth. In the literature, higher RLD for phacelia are found [
13,
20,
21]. In the rhizobox experiment of Bodner et al. [
33], phacelia was among the crops with highest total root length and highest proportion in deep soil layers out of 10 cover crop species. RLD of phacelia in the subsoil might be underestimated by the profile wall method as proportion of RLD in large-sized biopores for phacelia is high and high root length in biopores are more difficult to count than in bulk soil as there can be several roots in one biopore–however this does not explain the lower values we found in the topsoil.
In our study, differences in rooting depth were not very pronounced. Rooting depth of crimson clover was significantly lower than that of brassica cover crops in October 2018, supporting the potential of brassica crops for capturing nitrate in deeper soil layers already early after cover crop establishment. Crops with low rooting depth in autumn caught up with rooting depth during winter. However, at our study site, root analysis was limited by gravel layers in about 1 m soil depth. Studies from sites with a larger soil volume available for roots compared to our experimental site showed that rye, ryegrass and oat needed more growing degree-days to reach one meter soil depth than radish, winter rape and phacelia [
18], and that oil radish rooted much deeper, down to more than 2 m soil depth, as compared to winter rye with about 1 m soil depth [
16].
In summary, from the results of our study, our hypothesis (i) that topsoil root length density is higher for fibrous rooted crops can only partly be confirmed. Winter rye had the highest RLD of all cover crops, but RLD of bristle oat was only sometimes higher than RLD of brassica cover crops. Crimson clover, an intermediate type between fibrous and taprooted cover crops, had the second highest RLD in the topsoil. Hypothesis (ii) that subsoil RLD is higher for taprooted than fibrous rooted crops is partly confirmed; namely, before winter and for the lower subsoil (>60 cm), for all taprooted crops except for lupin. In the upper subsoil (30–60 cm) winter rye reached higher RLD than the taprooted cover crops. Literature results at large support the notion of winter rye and crimson clover as crops with intensive rooting in the upper soil layers and bristle oats with less intensive rooting in the topsoil. In contrast to our study, in some other studies topsoil RLD of the taprooted crops oil radish and phacelia outscored topsoil RLD of fibrous rooted crops. However, in these studies differences in the subsoil between these two taprooted and the respective fibrous rooted crops tested were even more pronounced.
4.2. Cumulative Distribution of Root Length
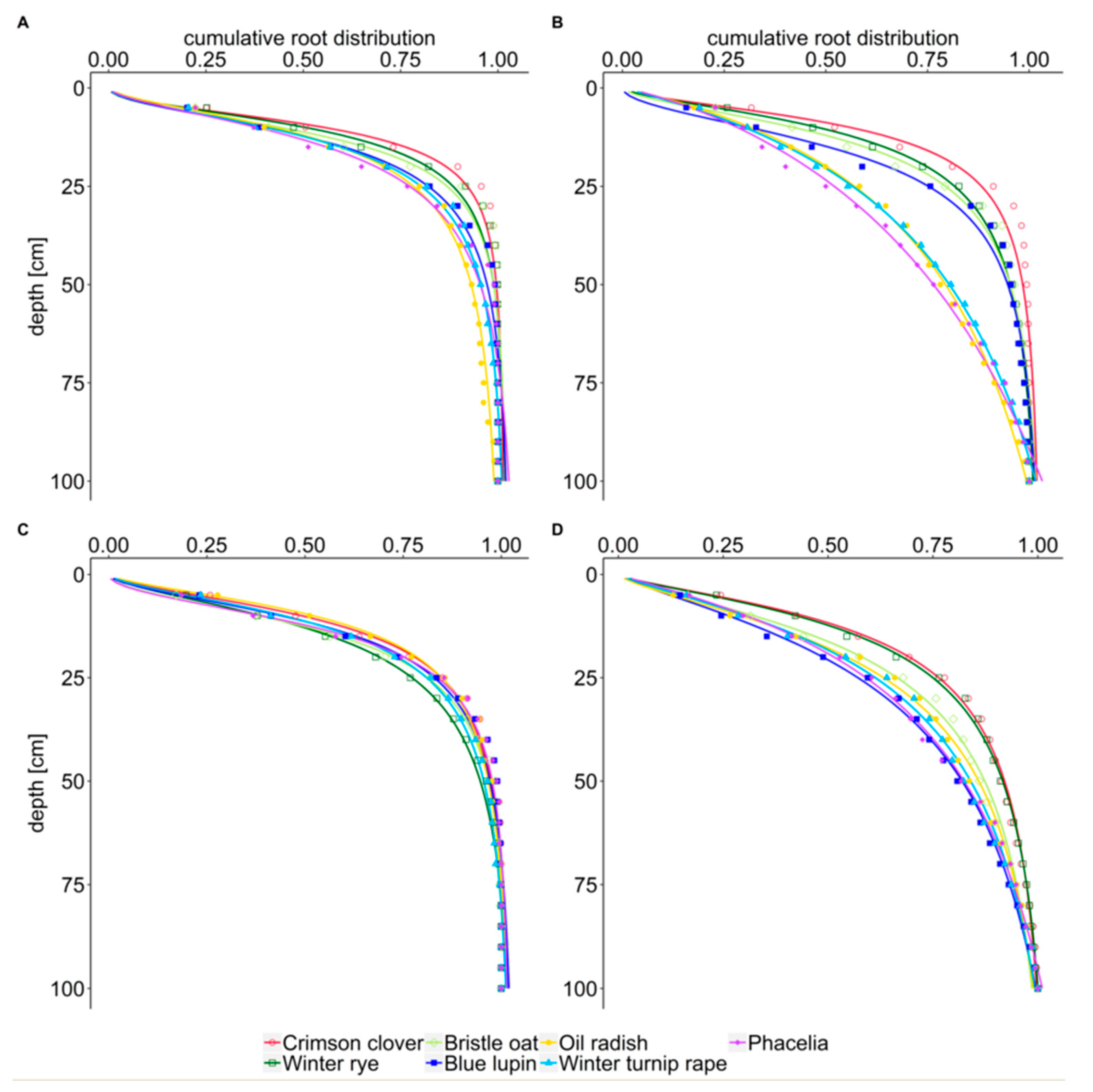
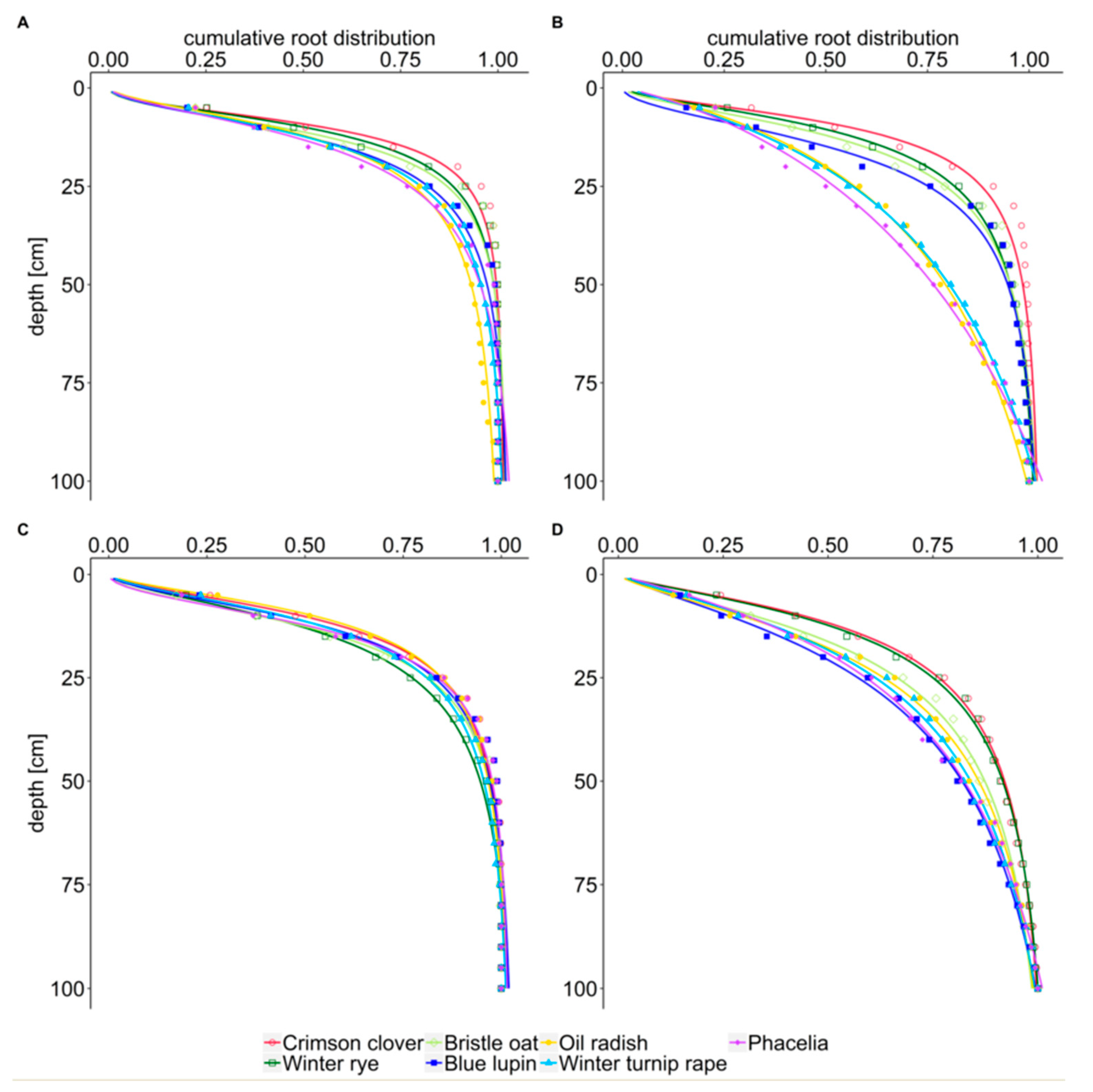
The cumulative distribution of root length is another approach to describe rooting characteristics. By comparing the cumulative percentage of root length with soil depth rather than the measured absolute values, this approach enables a rapid comparison of how the different species distribute their roots in the soil. As hypothesized (hypothesis iii), x50 and x95 values showed a shallow distribution of root length for fibrous rooted winter rye, bristle oat, and intermediate crimson clover, and a deep distribution for taprooted oil radish, phacelia, and winter turnip rape.
This result is confirmed by Thorup-Kristensen [
18] who showed a constant root frequency up to 1.2 m soil depth for radish, winter rape and phacelia, while root frequency of rye, oat and ryegrass declined between 0.0 m and 1.2 m soil depth. In their rhizobox experiment, Bodner et al. [
33] determined the highest proportion of roots in lower layers among ten species for oil radish and phacelia. Both studies confirm the high proportion of deep roots for allorhizous phacelia and brassica cover crops compared to homorhizous grasses. The intermediate position of blue lupin might be influenced by the low number of fine roots. The deeper root distribution of bristle oat compared to winter rye, especially after winter of the second experimental year, might be explained by its characteristics close to that of taprooted species, e.g., half maximal RLD at greater soil depths. Similarly, Bodner et al. [
33] determined bristle oat out of all cover crops with low root length as the species with the highest proportion of root length in deeper layers.
4.3. Winter Hardiness
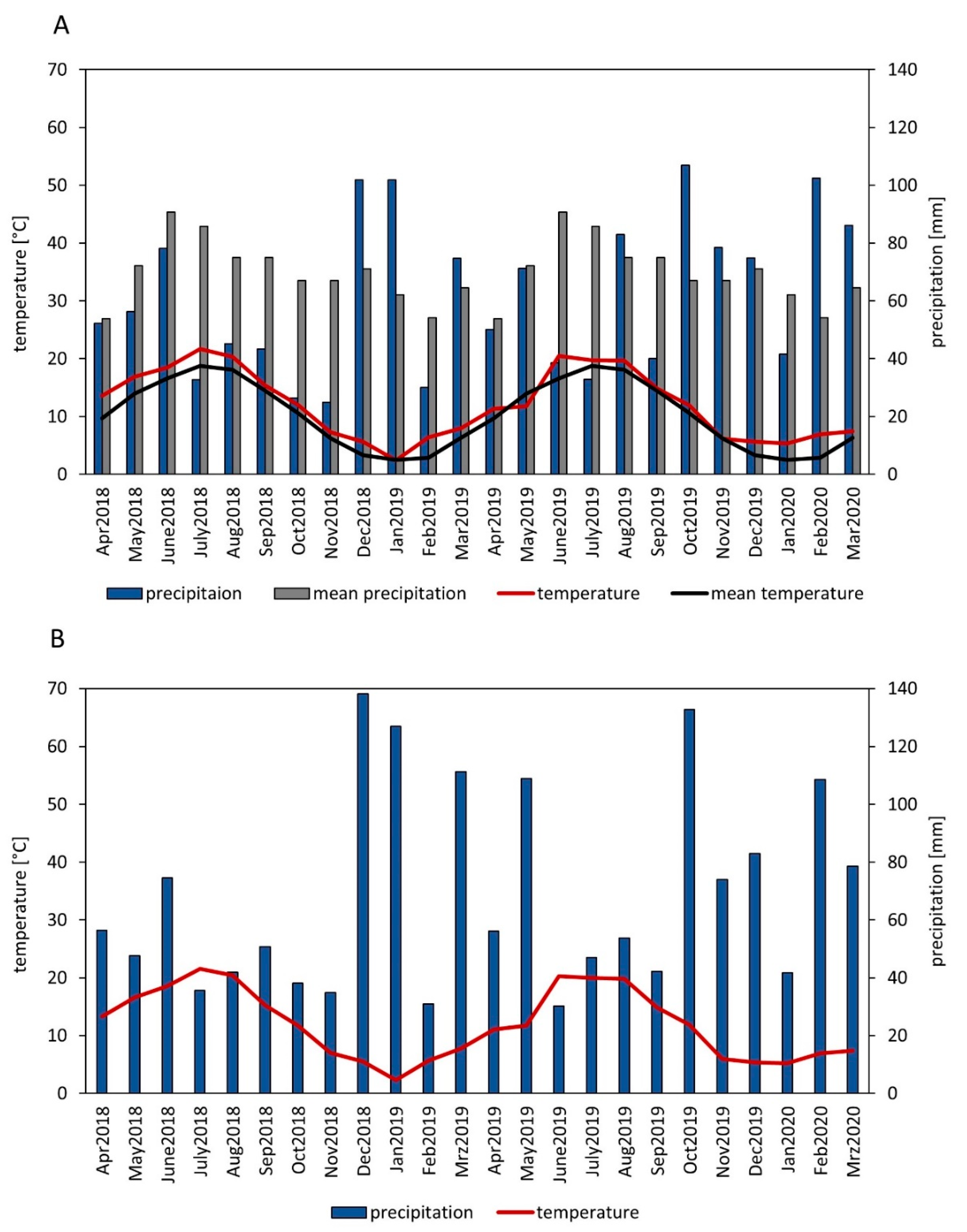
Compared to other cover crops RLD of winter rye and crimson clover increased strongly during winter (
Figure 2). The winter 2018/2019, esp. January 2019, was colder than the winter 2019/2020, (
Figure 1). Thus, in 2018/2019, all non-winter hardy species except oil radish, which was only partially dead, were completely frost-killed, whereas in 2019/2020 oil radish was not frost-killed and the other non-winter hardy species were only partially frost-killed. This might explain the differences in RLD change of the non-winter hardy cover crops. Hence, winter-hardy cover crops increase RLD during winter, whereas non-winter hardy species decrease or slightly increase RLD depending on frost intensity and duration. Thus, our hypothesis (iv) that non winter-hardy cover crops lose RLD during winter compared to winter hardy cover crops can only be confirmed for winters with enough frost.
4.4. Share of Root Length in Large-Sized Biopores in the Subsoil
Up to 32% of root length in large-sized biopores in the subsoil was determined (
Table 6). The values are in a similar range as in studies with barley on a Haplic Luvisol, reporting up to 21% [
26] or 25% [
28] of RLD in large-sized biopores. The highest share of root length in large-sized biopores was found for the allorhizous species phacelia, oil radish, and winter turnip rape. Thus, taprooted crops seem to use biopores to a larger extent. This might be due to different strategies for exploring biopores by different rooting types. With in situ endoscopy, it was shown that homorhizous crops had thin vertical or ingrowing roots in biopores, while taproots of allorhizous crops grow centrally through biopores, contacting the pore wall with laterals [
29] and that taprooted oilseed rape with thinner roots used biopores more intensively for root growth than taprooted faba bean with thick roots and only few laterals [
36]. When supposing similarity of oilseed rape with winter turnip rape and oil radish and between faba bean and blue lupin, the results of Athmann et al. [
36] could explain the differences in proportion of roots in biopores for brassica cover crops and blue lupin.
Bristle oat is the homorhizous crop using the pores most intensively. It is notable that bristle oat was the fibrous rooted species with characteristics closest to that of taprooted species, as can be seen from half maximal RLD at great soil depth and from cumulative root distribution. Blue lupin had a very low RLD throughout the soil profile, and many thick roots. Therefore, blue lupin had not enough fine roots to use the biopores to a large extent.
With the strong pore users phacelia, oil radish and winter turnip rape, our hypothesis (v) that allorhizous cover crop use large-sized biopores to a greater extent than homorhizous cover crops is confirmed. To our knowledge this is the first study comparing cover crop species with respect to the use of large-sized biopores by roots. Future studies have to show if a larger share of roots in subsoil biopores is associated with higher nutrient uptake from nutrient rich biopore walls, which would likely imply higher nutrient uptake from the subsoil in total and higher prevention of nitrate leaching as biopores are preferential flow paths.
4.5. Cover Crop Rooting Types
The information derived from analysis of root growth of the different cover crop species under study can be condensed to describe two basic cover crop rooting types: First, crops with dense root systems with many fine roots such as winter rye and crimson clover with high RLD in the topsoil which increases further during winter and a small x50 value. Second, taprooted oil radish, winter turnip rape and phacelia with high RLD in the subsoil, a greater x50 value, and a high percentage of root length in large-sized biopores. Blue lupin and bristle oat do not fit into either one of the categories. Bristle oat has intermediate features and blue lupin has low RLD in topsoil, subsoil and biopores.
Bodner et al. [
33] and Bodner et al. [
10] classified cover crop roots based on more detailed characteristics from root imaging and the Root Atlas [
37,
38], into four groups:
- (1)
Topsoil-allocated with large diameters: e.g., legumes like vetch, faba bean, Egyptian clover
- (2)
Topsoil-allocated with very dense root system and many fine roots: e.g., rye, buckwheat
- (3)
Root density group with highly branched dense rooting systems: e.g., oil radish, camelina, phacelia
- (4)
Low-order axes types like linseed and bristle oat with low-branched primary roots.
Out of the cover crops in our study, winter rye and crimson clover can be assigned to the second group, oil radish, winter turnip rape and phacelia to the third group, and bristle oat to the fourth group. According to the results of our study, lupin could be assigned to the first group. However, since lupin is well-known for deep rooting, following comparative studies have to verify this classification. Considering these different rooting types suggests combining cover crops with different root characteristics in intercropping to enhance RLD in the total soil volume. Thus, combining intensive topsoil rooting cover crops with a biopore-using taprooted cover crops may be auspicious.
4.6. The Profile Wall Method
Many studies with results of RLD for different cover crops can be found [
12,
13,
14,
16,
20,
21,
22,
33,
35]. In contrast to our study, they mostly rely on roots washed out from the soil, and only some differentiated into different depth layers. Often only values for single or cumulated depth levels are presented, and no profile is shown. For example, soil cores were examined at a soil depth ranging from 2–7 cm [
12,
13] to 100 cm depth [
35]. Segments of cores ranged between 5 cm [
20] and 40 cm [
35]. Diameter of soil cores ranged between 5.7 cm [
20] and 8.7 cm [
35]. In other studies, RLD was calculated from minirhizotron results reaching a depth of 2.42 m [
16] or images were taken of rhizoboxes of 30 × 100 × 1 cm [
33]. All these data provide valuable results. Different from the profile wall method, washed root samples enable determination of not only RLD, but also other root characteristics such as root mass, diameter, topology and chemical composition. In contrast, the strength of the profile wall method is that roots of a much larger number of plants enter into the assessment, that it is less time-consuming than root washing and therefore more depth levels and several assessment dates can be included to capture also root growth dynamics. RLD values are given in 5 cm steps for 1 m soil depth which allows more fine-grained regression analysis. Furthermore, as the position of the individual roots in the soil is maintained, it is possible to measure roots in biopores separately from roots in the bulk soil.
When comparing RLD results of the present study determined with the profile wall method with RLD results from the literature determined by washing roots from soil [
13,
14,
20,
21], it is conspicuous that RLD from the profile wall method is often smaller. However, relations of RLD between different cover crops are similar. That RLD determined by the profile wall method is lower than by washing roots from soil was already discussed by Böhm [
30].
It is noticeable that absolute RLD values differed between 2018/2019 and 2019/2020. In 2018/2019 RLD
5 ranges between 0.19 and 1.22 cm cm
−3, whereas in 2019/2020 RLD
5 ranged between 0.33 and 7.05 cm cm
−3. Absolute RLD values can vary depending on the person counting the root length at the profile wall, and for our study, two different persons were counting in the two study years. Consequently, the two seasons cannot be compared in terms of absolute RLD values. However, the relation of RLD between crops is reliable and allows a relative comparison between different crops. To improve the profile wall method and minimize the human factor, we suggest taking images of the profile wall and using computer image analysis. Suitable software has recently become available, e.g., RootPainter [
39].
5. Conclusions
The results of this study show that there are considerable differences between different cover crop species in RLD, root distribution between topsoil and subsoil and, in one of the two experimental years, also rooting depth. The notion that taprooted species explore the subsoil more and fibrous rooting species explore the topsoil more was confirmed for winter rye, oilseed rape, winter turnip rape and phacelia, but not for bristle oat and lupin. Fibrous rooted winter rye had a dense root system, a high topsoil RLD and a small x50 value. Similarly, crimson clover rooted very intensively in the topsoil despite not being classified as a fibrous rooted crop. Taprooted oilseed rape, winter turnip rape and phacelia had a high subsoil RLD and a greater x50 value. Bristle oat had intermediate features between fibrous rooted and taprooted species. Blue lupin has low RLD in topsoil as well as in subsoil.
Therefore, the range of currently widely used cover crops includes crops with very different rooting types as a basis for including this criterion in management decisions. Further research will show if the different rooting types result in differences in nitrate uptake, reduction of soil erosion, soil organic carbon enhancement and soil structure improvement, and if combining cover crops of contrasting rooting types improve overall exploitation of the total soil volume. Also, the implications of differences in biopore use of the different cover crop species for nutrient uptake from the subsoil need further research.
The profile wall method is well suitable to present RLD and cumulative root distribution with non-linear regression as a function of soil depth. Further improvement of the method is necessary to increase comparability of different years, possibly via replacing manual countings with computer image analysis.


 and
and 


{kind=link}
{kind=link}
{kind=link}