The Critical Role of Zinc in Plants Facing the Drought Stress


 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Effects of Drought Stress on Plants
3. General Mechanisms of Plant Response to Drought Stress
4. Zinc Uptake and Transport in Plants
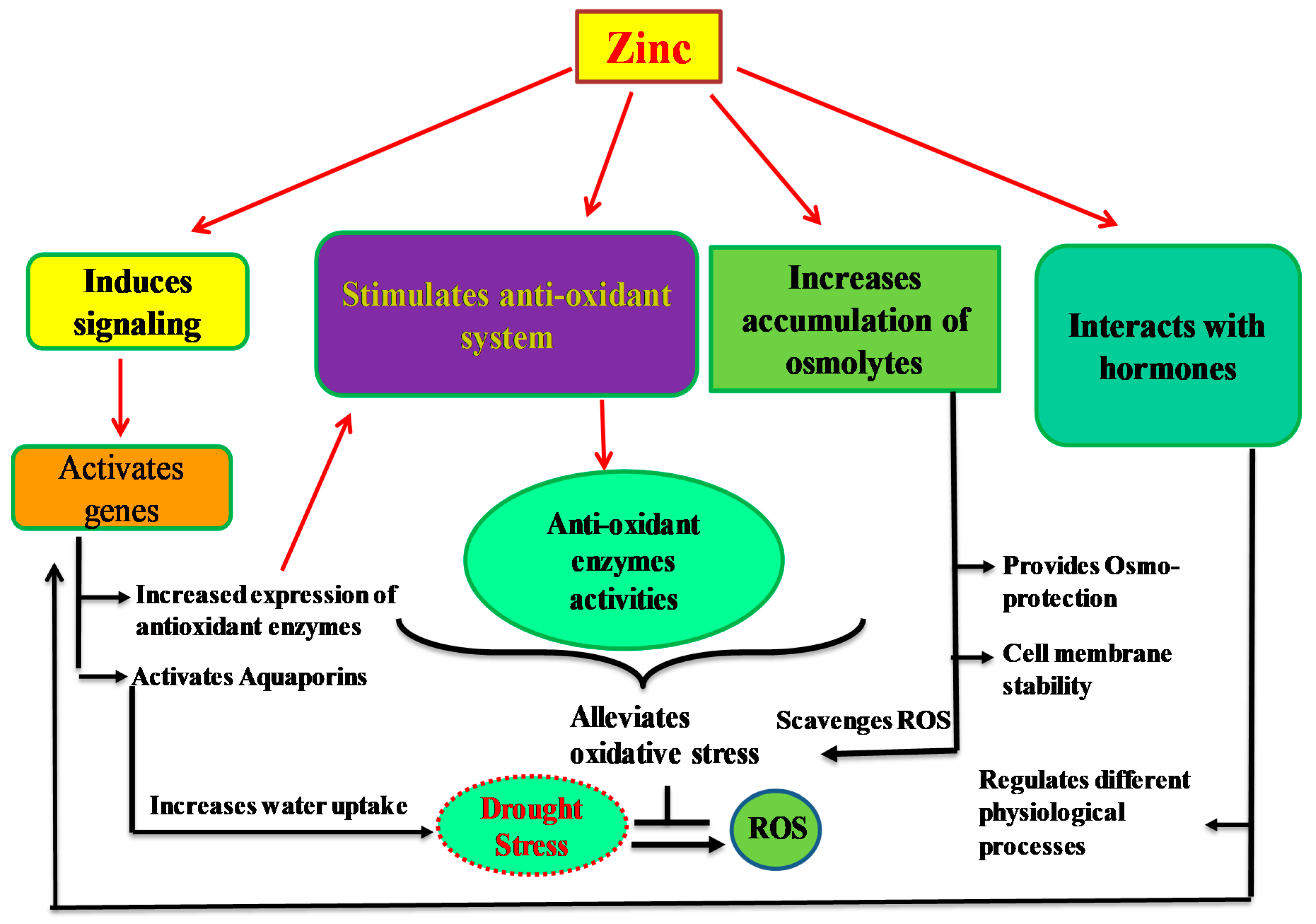
5. Roles of Zn in Plants under Drought Stress
5.1. Effects of Zn on Plant Growth and Development under Drought Stress
5.2. Effect of Zn on Plant Water Relations in Drought Stress

{kind=link}
| Crop | Drought Stress | Effects | References |
|---|---|---|---|
| Maize | 40% field capacity | Zn application increases plant biomass, stomatal conductance and quantum yield of photosystem II under drought stress. | [80] |
| Maize | Drought stress at pollination and seed filling | Zn improves grain yield, RWC and chlorophyll content. | [81] |
| Sunflower | Drought stress at seed filling stage | Zn improves seed and oil yield, chlorophyll and proline contents and SOD activities. | [82] |
| Wheat | Drought stress at grain filling stage | Zn application improves yield, chlorophyll content and activities of SOD, POD and CAT. | [83] |
| Triticale | Water stress at heading and booting | Zn application increases the activities of POD, CAT, quantum yield of PS-II, chlorophyll content, stomatal conductance and reduces the electrical conductivity. | [75] |
| Common bean | Irrigation applied at interval of 6, 12 and 18 days | Zn increases plant height, shoot biomass, chlorophyll and carotenoid contents, leaf NPK content and decreases the MDA contents | [84] |
| Tomato | With-held irrigation | Zn supplementation improves the stomatal aperture and chlorophyll content. | [85] |
| Chickpea | 35% field capacity | Zn leads to improvement in PS-II efficiency, CO2 assimilation rate, proline content and activities of SOD, APX, while reduces the MDA contents. | [86] |
| Chickpea | 35% field capacity | Zn application increases chlorophyll and carotenoid contents, seedling vigor and seed yield. | [87] |
| Wheat | Drought stress at heading and an-thesis stage | Zn improves stomatal conductance, WUE, efficiency of PS-II, chlorophyll content and reduces energy dissipation | [88] |
5.3. Effect of Zn on Cell Membrane Stability under Drought Stress
5.4. Regulation of Osmolyte Accumulation by Zn under Drought Stress
5.5. Effect of Zn on Stomatal Regulation and Water Use Efficiency under Drought Stress
5.6. Effect of Zn on Nutrient Uptake under Drought Stress
5.7. Effect of Zn on Photosynthesis under Drought Stress
5.8. Zn-Mediated Cross Talk with Hormones under Drought Stress
5.9. Detoxification of Reactive Oxygen Species by Zn under Drought Stress
5.10. Effect of Zn on Stress Proteins under Drought Stress
5.11. Effect of Zn on the Activities of Aquaporins under Drought Stress
6. Methods of Zinc Application to Crops
7. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba. J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar]
- Whitmore, T.C. The case of tropical rain forests. The sustainable development of forests: Aspiration and the reality. Naturzale 2000, 15, 13–15. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Naeem, M.; Muhammad, S.N.; Rashid, A.; Muhammad, Z.I.; Muhammad, Y.A.; Yasir, H.; Fahad, S. Foliar calcium spray confers drought stress tolerance in maize via modulation of plant growth, water relations, proline content and hydrogen peroxide activity. Arch. Agron. Soil Sci. 2018, 64, 116–131. [Google Scholar] [CrossRef]
- Saud, S.; Chen, Y.; Fahad, S.; Hussain, S.; Na, L.; Xin, L.; Alhussien, S. Silicate application increases the photosynthesis and its associated metabolic activities in Kentucky bluegrass under drought stress and post-drought recovery. Environ. Sci. Pollut. Res. 2016, 23, 17647–17655. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Effects of drought, heat and their interaction on the growth, yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity. Front. Plant Sci. 2017, 8, 1776. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.M.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148. [Google Scholar] [CrossRef] [Green Version]
- Rahbarian, R.; Khavari-Nejad, R.; Ganjeali, A.; Bagheri, A.; Najafi, F. Drought stress effects on photosynthesis, chlorophyll fluorescence and water relations in tolerant and susceptible chickpea genotypes. Acta Biol. Cracov. Bot. 2011, 53, 47–56. [Google Scholar] [CrossRef]
- Samarah, N.H.; Alqudah, A.M.; Amayreh, J.A.; McAndrews, G.M. The effect of lateterminal drought stress on yield components of four barley cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Gunes, A.; Cicel, N.; Inal, A.; Alpaslan, M.; Eraslan, F.; Guneri, E.; Guzelordu, T. Genotypic response of chickpea (Cicer arietinum L.) cultivars to drought stress implemented at pre- and post-anthesis stages and its relations with nutrient uptake and efficiency. Plant Soil Environ. 2006, 52, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Mar, J.T.; Munn-Bosch, S.; Alegre, L. Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol. Biochem. 2010, 48, 351–358. [Google Scholar]
- Wu, S.; Hu, C.; Tan, Q.; Nie, Z.; Sun, X. Effects of molybdenum on water utilization, antioxidative defense system and osmotic adjustment ability in winter wheat (Triticum aestivum) under drought stress. Plant Physiol. Biochem. 2014, 83, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Molden, D.; Mainuddin, M.; Sharma, B.; Mohyddin, A.; Karini, P. Producing more food with less water in the changing world: Assessment of water productivity in 10 major river basins. Water Int. 2013, 36, 421–462. [Google Scholar] [CrossRef]
- Tsonev, T.; Lidon, F.J.C. Zinc in plants—An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Mengel, K.; Kosegarten, H.; Kirkby, E.A.; Appel, T. Principles of Plant Nutrition; Springer: Berlin, Germany, 2001. [Google Scholar]
- Chattha, M.U.; Hassan, M.U.; Khan, I.; Chattha, M.B.; Mahmood, A.; Chattha, M.U.; Nawaz, M.; Subhani, M.N.; Kharal, M.; Khan, S. Biofortification of Wheat Cultivars to Combat Zinc Deficiency. Front. Plant Sci. 2017, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.U.; Chattha, M.U.; Ullah, A.; Khan, I.; Qadeer, A.; Aamer, M.; Khan, A.U.; Nadeem, F.; Khan, T.A. Agronomic biofortification to improve productivity and grain Zn concentration of bread wheat. Int. J. Agric. Biol. 2019, 21, 615–620. [Google Scholar]
- Mousavi, S.R.; Galavi, M.; Ahmadvand, G. Effect of zinc and manganese foliar application on yield, quality and enrichment on potato (Solanum tuberosum L.). Asian J. Plant Sci. 2007, 6, 1256–1260. [Google Scholar]
- Hajiboland, R.; Amirazad, F. Drought tolerance in Zn-deficient red cabbage (Brassica oleracea L. var. capitata f. rubra) plants. Hortic. Sci. 2010, 37, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Sadoogh, F.S.; Shariatmadari, H.; Khoshgoftarmanesh, A.H.; Mosaddeghi, M.R. Adjusted nutrition of tomato with potassium and zinc in drought stress conditions induced by polyethylene glycol 6000 in hydroponic culture. J. Sci. Technol. Greenh. Cult. 2014, 18, 67–80. [Google Scholar]
- Eslami, M.; Dehghanzadeh, H. The effect of zinc on yield and yield components of sunflower (Helianthus annuus L.) under drought stress. Sci. J. Crop Sci. 2014, 3, 61–65. [Google Scholar]
- Zlatev, Z.; Lidon, F.C. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emir. J. Food Agric. 2012, 24, 57–72. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, E. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zare, M.; Azizi, M.H.; Bazrafshan, F. Effect of drought stress on some agronomic traits in ten barley (Hordeum vulgare) cultivars. Tech. J. Eng. Appl. Sci. 2011, 1, 57–62. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Sapeta, H.; Costa, M.; Lourenc, T.; Marocod, J.; Van-der Linde, P.; Oliveiraa, M.M. Drought stress response in Jatropha curcas: Growth and physiology. Environ. Exp. Bot. 2013, 85, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, Y.K.; Yadav, S.K. Effect of high-temperature stress on ascorbate–glutathione cycle in maize. Agric. Res. 2020, 9, 179–187. [Google Scholar] [CrossRef]
- Xoconostle-Cazares, B.; Ramirez-Ortega, F.A.; Flores-Elenes, L.; Ruiz-Medrano, R. Drought tolerance in crop plants. Am. J. Plant Physiol. 2010, 5, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Iqbal, M.; Jameel, M.; Nazeer, W.; Shakir, S.; Aslam., M.T.; Iqbal, B. Potentials of molecular based breeding to enhance drought tolerance in wheat (Triticum aestivum L.). Afr. J. Biotechnol. 2011, 10, 11340–11344. [Google Scholar]
- Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- Ding, Y.; Tao, Y.; Zhu, C. Emerging roles of microRNAs in the mediation of srought stress responses in plants. J. Exp. Bot. 2013, 64, 3077–3086. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Chernyad’ev, I.I. Effect of water stress on the photosynthetic apparatus of plants and the protective role of cytokinins: A review. Appl. Biochem. Microbiol. 2005, 41, 115–128. [Google Scholar] [CrossRef]
- Kheradmand, M.A.; Shahmoradzadeh, F.S.; Fatahi, E.; Raoofi, M.M. Effect of water stress on oil yield and some characteristics of Brassica napus. Int. Res. J. Appl. Basic Sci. 2014, 8, 1447–1453. [Google Scholar]
- Bernacchia, G.; Furini, A. Biochemical and molecular responses to water stress in resurrection plants. Physiol. Plant. 2004, 121, 175–181. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. Book Am. Soc. Plant Biol. 2013, 11, e0166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahdari, P.; Hoseini, S.M. Drought stress: A review. Int. J. Agron. Plant Prod. 2012, 3, 443–446. [Google Scholar]
- Salehi-Lisar, S.Y.; Bakhshayeshan, A.H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2016; Volume 1, pp. 1–16. [Google Scholar]
- Rana, R.M.; Rehman, S.U.; Ahmed, J.; Bilal, M. A comprehensive overview of recent advances in drought stress tolerance research in wheat (Triticum aestivum L.). Asian J. Agric. Biol. 2013, 1, 29–37. [Google Scholar]
- Gupta, N.; Ram, H.; Kumar, B. Mechanism of Zinc absorption in plants: Uptake, transport, translocation and accumulation. Rev. Environ. Sci. Biol. Technol. 2016, 15, 89–109. [Google Scholar] [CrossRef]
- Palmer, C.M.; Guerinot, M.L. Facing the challenges of Cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, B.; Johnson, A.; Walter, P.; Lewis, J.; Raff, M.; Roberts, K. Molecular Biology of the Cell, 5th ed.; Taylor & Francis, Garland Science: New York, NY, USA, 2007. [Google Scholar]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta (BBA) Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Clemens, S.; Palmgren, M.G.; Kramer, U. A long way ahead: Understanding and engineering plant metal accumulation. Trends Plant Sci. 2002, 7, 309–315. [Google Scholar] [CrossRef]
- Demidchik, V.; Davenport, R.J.; Tester, M. Non selective cation channels in plants. Annu. Rev. Plant Biol. 2002, 53, 67–107. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, B.; Yu, J.; Ao, G.; Zhao, Q. Cloning and characterisation of ZmZLP1, a gene encoding an endoplasmic reticulum-localised Zn transporter in Zea mays. Funct. Plant Biol. 2010, 37, 194–205. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Choi, H.; Segami, S.; Cho, H.T.; Martinoia, E.; Maeshima, M.; Lee, Y. AtHMA1 contributes to the detoxification of excess Zn (II) in Arabidopsis. Plant J. 2009, 58, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Hanikenne, M.; Talke, I.N.; Haydon, M.J.; Lanz, C.; Nolte, A.; Motte, P.; Kroymann, J.; Weigel, D.; Kramer, U. Evolution of metal hyper accumulation required cis-regulatory changes and triplication of HMA4. Nature 2008, 453, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Le-Jean, M.; Schikora, A.; Mari, S.; Briat, J.F.; Curie, C. A lossof-function mutation in AtYSL1 reveals its role in iron and nicotianamine seed loading. Plant J. 2005, 44, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Desbrosses, F.A.G.; Voigt, K.; Schroder, A.; Arrivault, S.; Thomine, S.; Kramer, U. Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 2005, 579, 4165–4174. [Google Scholar] [CrossRef]
- Song, W.Y.; Choi, K.S.; Kim, D.Y.; Geisler, M.; Park, J.; Vincenzetti, V.; Schellenberg, M.; Kim, S.H.; Lim, Y.P.; Noh, E.W.; et al. ArabidopsisPCR2 is a zinc exporter involved in both zinc extrusion and long-distance zinc transport. Plant Cell 2010, 22, 2237–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xu, Y.H.; Yi, H.Y.; Gong, J.M. Vacuolar membrane transporters OsVIT1 and OsVIT2 modulate iron translocation between flag leaves and seeds in rice. Plant J. 2012, 72, 400–410. [Google Scholar] [CrossRef]
- Sondergaard, T.E.; Schulz, A.; Palmgren, M.G. Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase? Plant Physiol. 2004, 136, 2475–2482. [Google Scholar] [CrossRef] [Green Version]
- Sedghi, M.; Hadi, M.; Toluie, S.G. Effect of nano zinc oxide on the germination parameters of soybean seeds under drought stress. Ann. West Univ. Timis. Ser. Bio. 2013, 16, 73. [Google Scholar]
- Harris, D.; Rashid, A.; Arif, M.; Yunas, M. Alleviating micronutrient deficiencies in alkaline soils of the North-West Frontier Province of Pakistan: On-farm seed priming with zinc in wheat and chickpea. In Micronutrients in South and South East Asia; Andersen, P., Tuladhar, J.K., Karki, K.B., Maskey, S.L., Eds.; International Centre for Integrated Mountain Development International: Kathmandu, Nepal, 2005; pp. 143–151. [Google Scholar]
- Harris, D.; Rashid, A.; Miraj, G.; Arif, M.; Shah, H. On-farm seed priming with zinc sulphate solution—A cost-effective way to increase the maize yields of resource-poor farmers. Field Crops Res. 2007, 10, 119–127. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification. Plant Soil 2008, 30, 1–17. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: New York, NY, USA, 1995. [Google Scholar]
- Hera, M.H.R.; Hossain, M.; Paul, A.K. Effect of foliar zinc spray on growth and yield of heat tolerant wheat under water stress. Int. J. Biol. Environ. Eng. 2018, 1, 10–16. [Google Scholar]
- Bajjii, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Karam, F.; Lahoud, R.; Masaad., R.; Kabalan, R.; Breidi, J.; Chalita, C.; Rouphael, Y. Evaporation, seed yield and water use efficiency of drip irrigated sunflower under full and deficit irrigation conditions. Agric. Water Manag. 2007, 90, 213–223. [Google Scholar] [CrossRef]
- Bagci, S.A.; Ekiz, H.; Yilmaz, A.; Cakmak, I. Effect of zinc deficiency and drought on grain yield of field-grown wheat cultivars in Central Anatolia. J. Agron. Crop Sci. 2007, 193, 198–206. [Google Scholar] [CrossRef]
- Babaeian, M.; Heidari, M.; Ghanbari, A. Effect of water stress and foliar micronutrient application on physiological characterstics and nutrient uptake in sunflower (Helianthus annuus L.). Iran. J. Crop Sci. 2010, 12, 311–391. [Google Scholar]
- Karim, M.; Zhang, Y.Q.; Zhao, R.R.; Chen, X.P.; Zhang, F.S.; Zou, C.Q. Alleviation of drought stress in winter wheat by late foliar application of zinc, boron, and manganese. J. Plant Nutr. Soil Sci. 2012, 175, 142–151. [Google Scholar] [CrossRef]
- Sultana, S.; Naser, H.M.; Shil, N.C.; Akhter, S.; Begum, R.A. Effect of foliar application of zinc on yield of wheat grown by avoiding irrigation at different growth stages. Bangladesh J. Agric. Res. 2016, 41, 323–334. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Rodríguez, P.; Morales, M.A.; Ortuño, M.F.; Torrecillas, A. Comparative growth and water relation of Cistusalbidus and Cistusmons peliensis plants during water deficit conditions and recovery. Plant Sci. 2002, 162, 107–113. [Google Scholar] [CrossRef]
- Velu, G.; Palanisami, K. Impact of moisture stress on growth and yield of sunflower. Madras Agric. J. 2002, 88, 660–665. [Google Scholar]
- Khan, R.; Gul, S.; Hamayun, M.; Shah, M.; Sayyed, A.; Ismail, H.; Begum, A.; Gul, H. Effect of foliar application of zinc and manganese on growth and some biochemical constituents of Brassica junceae grown under water stress. Am.-Eurasian J. Agric. Environ. Sci. 2016, 16, 984–997. [Google Scholar]
- Arough, Y.; Seyed-Sharifi, R.; Seyed-Sharifi, R. Bio fertilizers and zinc effects on some physiological parameters of triticale under water-limitation condition. J. Plant Interact. 2016, 11, 167–177. [Google Scholar] [CrossRef]
- Grewal, H.S.; Williams, R. Zinc nutrition affects alfalfa responses to water stress and excessive moisture. J. Plant Nutri. 2000, 23, 949–962. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2002, 51, 1531–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadallah, M.A.A.; Ramadan, T. Effects of zinc and salinity on growth and anatomical structure of Carthamus tinctorius L. Biolgia Plant. 1997, 39, 411–418. [Google Scholar] [CrossRef]
- Gadallah, M.A.A. Effects of indole-3-acetic acid and zinc on growth, osmotic potential and soluble carbon and nitrogen components of soybean plants growing under water deficit. J. Arid Environ. 2000, 44, 451–467. [Google Scholar] [CrossRef]
- Wang, H.; Liu, R.L.; Jin, J.Y. Effects of zinc and soil moisture on photosynthetic rate and chlorophyll fluorescence parameters of maize. Biol. Plant. 2009, 53, 191–194. [Google Scholar] [CrossRef]
- Vazin, F. Effect of zinc sulfate on quantitative and qualitative characteristics of corn (Zea mays) in drought stress. Cercet. Agron. Mold. 2012, 45, 15–24. [Google Scholar] [CrossRef]
- Zafar, S.; Nasri, M.; Moghadam, H.R.T.; Zahedi, H. Effect of zinc and sulfur foliar applications on physiological characteristics of sunflower (Helianthus annuus L.) under water deficit stress. Int. J. Biosci. 2014, 5, 87–96. [Google Scholar]
- Yavas, I.; Unay, A. Effects of zinc and salicylic acid on wheat under drought stress. J. Anim. Plant Sci. 2016, 26, 1012–1101. [Google Scholar]
- Ibrahim, S.A.; Desoky, E.; Elrys, A.S. Influencing of water stress and micronutrients on physio-chemical attributes, yield and anatomical features of Common Bean plants (Phaseolus vulgaris L.). Egypt. J. Agron. 2017, 39, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Sakya, A.T.; Sulistyaningsih, E.; Indradewa, D.; Purwanto, B.H. Stomata character and chlorophyll content of tomato in response to Zn application under drought condition. IOP Conf. Ser. Earth Environ. Sci. 2018, 142, 012033. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Romdhane, L.; Rehman, A.; Farooq, M. Adequate zinc nutrition improves the tolerance against drought and heat stresses in chickpea. Plant Physiol. Biochem. 2019, 143, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Kanwal, H.; Kausar, A.; Ilyas, A.; Akhter, N.; Ilyas, M.; Nisa, Z.; Khalid, H. Seed priming with zinc modulate growth, pigments and yield of chickpea (Cicer Arietinum L.) under water deficit conditions. Appl. Ecol. Environ. Res. 2019, 17, 147–160. [Google Scholar] [CrossRef]
- Pavia, I.; Roque, J.; Rocha, L.; Ferreira, H.; Castro, C.; Carvalho, A.; Silva, E.; Brito, C.; Gonçalves, A.; Lima-Brito, J.; et al. Zinc priming and foliar application enhances photoprotection mechanisms in drought-stressed wheat plants during anthesis. Plant Physiol. Biochem. 2019, 140, 27–42. [Google Scholar] [CrossRef]
- Liu, X.Z.; Huang, B.R. Heat stress injury in relation to membrane lipid peroxidation in creeping bent grass. Crop Sci. 2000, 40, 503–510. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weshke, W. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiolgia Plant. 2000, 109, 435–442. [Google Scholar] [CrossRef]
- Ghanepour, S.; Shakiba, M.R.; Toorchi, M.; Oustan, S. Role of Zn nutrition in membrane stability, leaf hydration status, and growth of common bean grown under soil moisture stress. J. Biodivers. Environ. Sci. 2015, 6, 9–20. [Google Scholar]
- Blokhina, O.; Virolainen, E.; Fagersttedt, K.V. Antioxidants, oxidative damage, and oxygen deprivation stress. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- DaCosta, M.; Huang, B.R. Osmotic adjustment associated with variation in bent grass tolerance to drought stress. J. Am. Soc. Hortic. Sci. 2006, 131, 338–344. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential, are they compatible, dissonant, or mutually exclusive? Crop Pasture Sci. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Kiani, S.P.; Talia, P.; Maury, P.; Grieu, P.; Heinz, R.; Perrault, A.; Nishinakamasu, V.; Hopp, E.; Gentzbittel, L.; Paniego, N. Genetic analysis of plant water status and osmotic adjustment in recombinant inbred lines of sunflower under two water treatments. Plant Sci. 2007, 172, 773–787. [Google Scholar] [CrossRef]
- Mousavi, S.R. Zinc in crop production and interaction with phosphorus. Aust. J. Basic Appl. Sci. 2011, 9, 1503–1509. [Google Scholar]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Bai, X.; Zhu, D.; Li, Y.; Ji, W.; Cai, H.; Zhu, Y. GsZFP1, a new Cys2/His2-type zinc-finger protein, is a positive regulator of plant tolerance to cold and drought stress. Planta 2012, 235, 1141–1155. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hu, C.; Tan, Q.; Li, L.; Shi, K.; Zheng, Y.; Sun, X. Drought stress tolerance mediated by zinc-induced antioxidative defense and osmotic adjustment in cotton (Gossypium hirsutum). Acta Physiol. Plant. 2015, 37, 167. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, E.; Ezz El-Din, A.; Omer, E.A. Influence of zinc and iron on plant growth and chemical constituents of Cymbopogon citrates L. grown in newly reclaimed land. Int. J. Acad. Res. 2010, 2, 278–283. [Google Scholar]
- Dehnavi, M.M.; Sheshbahre, M.J. Soybean leaf physiological responses to drought stress improved via enhanced seed zinc and iron concentrations. J. Plant Process. Funct. 2017, 5, 13–21. [Google Scholar]
- Dehnavi, M.M.; Misagh, M.; Yadavi, A.; Merajipoor, M. Physiological responses of sesame (Sesamum indicum L.) to foliar application of boron and zincunder drought stress. J. Plant Process Funct. 2017, 6, 27–36. [Google Scholar]
- Zushi, K. Comparison of chemical composition contents of tomato fruit grown under water and salinity stresses. J. SHITA 2005, 17, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Mishra, L.K.; Abidi, A.B. Phosphorus-zinc interaction: Effects on yield components, biochemical composition and bread making qualities of wheat. World Appl. Sci. 2010, 10, 568–573. [Google Scholar]
- Bharti, K.; Pandey, N.; Shankhdhar, D.; Srivastava, P.C.; Shankhdhar, S.C. Improving nutritional quality of wheat through soil and foliar zinc application. Plant Soil Environ. 2013, 59, 348–352. [Google Scholar] [CrossRef]
- Sharma, P.N.; Tripathi, A.; Bisht, S.S. Zinc requirement for stomatal opening in cauliflower. Plant Physiol. 1995, 107, 751–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.R.; McDonald, G.K.; Rengel, Z. Zinc fertilization and water stress affects plant water relations, stomatal conductance and osmotic adjustment in chickpea (Cicer arientinum L.). Plant Soil 2004, 267, 271–284. [Google Scholar] [CrossRef]
- Khan, H.R.; McDonald, G.K.; Rengel, Z. Zn fertilization improves water use efficiency, grain yield and seed Zn content in chickpea. Plant Soil 2003, 249, 389–400. [Google Scholar] [CrossRef]
- Hall, A.E.; Richards, R.A.; Condon, A.G.; Wright, G.C.; Farquhar, G.D. Carbon isotope discrimination and plant breeding. Plant Breed. Rev. 1994, 12, 81–113. [Google Scholar]
- Ahmed, N.; Ahmad, F.; Abid, M.; Ullah, M.A. Impact of zinc fertilization on gas exchange characteristics and water use efficiency of cotton crop under arid environment. Pak. J. Bot. 2009, 41, 2189–2197. [Google Scholar]
- Ahanger, M.A.; MoradTalab, N.; Fathi Abd-Allah, E.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. In Water Stress and Crop Plants: A Sustainable Approach; John Wiley & Sons, Ltd.: West Sussex, UK, 2016; Volume 2. [Google Scholar]
- Hajiboland, R. Effect of Micronutrient Deficiencies on Plants Stress Responses. In Abiotic Stress Responses in Plants; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 281–330. [Google Scholar]
- Hong, W.; Ji-Yun, J. Effects of zinc deficiency and drought on plant growth and metabolism of reactive oxygen species in maize (Zea mays L.). Agric. Sci. China 2007, 6, 988–995. [Google Scholar]
- Babaeian, M.; Piri, I.; Tavassoli, A.; Esmaeilian, Y.; Gholami, H. Effect of water stress and micronutrients (Fe, Zn and Mn) on chlorophyll fluorescence, leaf chlorophyll content and sunflower nutrient uptake in Sistan region. Afr. J. Agric. Res. 2011, 6, 3526–3531. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764. [Google Scholar]
- Moradi, L.; Ehsanzadeh, P. Effects of Cd on photosynthesis and growth of safflower (Carhamus tinctorius L.) genotypes. Photosynthetica 2015, 53, 506–518. [Google Scholar] [CrossRef]
- Peleg, Z.; Saranga, Y.; Yazici, A.; Fahima, T.; Ozturk, L.; Cakmak, I. Grain zinc, iron and protein concentrations and zinc-efficiency in wild emmer wheat under contrasting irrigation regimes. Plant Soil 2008, 306, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological responses and yield of wheat plants in zinc-mediated alleviation of drought stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balashouri, P. Effect of zinc on germination, growth, pigment content and phytomass of Vigna radiata and Sorghum bicolor. J. Ecobiol. 1995, 7, 109–114. [Google Scholar]
- Cakmak, I. Tansley Review No. 111–possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D. Response of tomato (Lycopersicon esculentum L.) cultivars to application of zinc when grown in sand culture at low zinc. Sci. Hortic. 2002, 93, 53–64. [Google Scholar] [CrossRef]
- Ayad, H.S.; Reda, F.; Abdalla, M.S.A. Effect of putrescine and zinc on vegetative growth, photosynthetic pigments, lipid peroxidation and essential oil content of geranium (Pelargonium graveolens L.). World J. Agri. Sci. 2010, 6, 601–608. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates Inc. Publishers: Sunderland, MA, USA, 2006. [Google Scholar]
- Bennett, J.P.; Skoog, F. Preliminary Experiments on the Relation of Growth-promoting Substances to the Rest Period in Fruit Trees. Plant Physiol. 2002, 13, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Moghadam, H.R.T.; Zahedi, H.; Ashkiani, A. Effect of zinc foliar application on auxin and gibberellin hormones and catalase and superoxide dismutase enzyme activity of corn (Zea mays L) under water stress. Maydica 2013, 58, 218–223. [Google Scholar]
- Jamil, M.; Ahamd, M.; Anwar, F.; Zahir, Z.A.; Kharal, M.A.; Nazli, F. Inducing drought tolerance in wheat through combined use of L-tryptophan and pseudomonas fluorescens. Pak. J. Agric. Sci. 2018, 55, 331–337. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a potent and inducible endogenous antioxidant: Synthesis and metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, I.U.; Baloch, M.S.; Sadozai, N.S.; Sulemani, M.Z. Stimulatory effect of GA3 and IAA on ripening process, kernel development and quality of rice. Pak. J. Biol. Sci. 1999, 2, 410–412. [Google Scholar]
- Fleta-Soriano, E.; Diaz, L.; Bonet, E.; Munne-Bosch, S. Melatonin may exert a protective role against drought stress in maize. J. Agron. Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P. Melatonin and its effects on plant systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Song, F.; Guo, J.; Zhu, X.; Liu, S.; Liu, F.; Li, X. Nano-ZnO-Induced Drought Tolerance Is Associated with Melatonin Synthesis and Metabolism in Maize. Int. J. Mol. Sci. 2020, 21, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Gratao, P.L.; Monteiro, C.C.; Antunes, A.M.; Peres, L.; Azevedo, R.A. Acquired tolerance of tomato (Lycopersicon esculentum cv. Micro-Tom) plants to cadmium-induced stress. Ann. Appl. Biol. 2008, 153, 321–333. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Galavi, M.; Rezaei, M. Zinc importance for crop production—A review. Int. J. Agron. Plant Prod. 2013, 4, 64–68. [Google Scholar]
- Li, G.; Tai, F.; Zheng, Y.; Luo, J.; Gong, S.; Zhang, Z.; Li, X. Two cotton Cys2/His2-type zinc-finger proteins, GhDi19-1 and GhDi19-2, are involved in plant response to salt/drought stress and abscisic acid signaling. Plant Mol. Biol. 2010, 74, 437–452. [Google Scholar] [CrossRef]
- Thounaojam, T.C.; Panda, P.; Choudhury, S.; Patra, H.K.; Panda, S.K. Zinc ameliorates copper-induced oxidative stress in developing rice (Oryza sativa L.) seedlings. Protoplasma 2014, 251, 61–69. [Google Scholar] [CrossRef]
- Chen, Z.; Young, T.E.; Ling, J.; Chang, S.C.; Gallie, D.R. Increasing vitamin C content of plants through enhanced ascorbate recycling. Proc. Natl. Acad. Sci. USA 2003, 100, 3525–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Tang, Y.Z.; Zhang, R.X.; Wang, W. Effect of fertilizer activating agent on the ascorbate-glutathione cycle and potassium content in tobacco leaves. J. Nanjing Agric. Univ. 2000, 23, 13–16. [Google Scholar]
- Tavallali, V.; Rahemi, M.; Eshghi, S.; Kholdebarin, B.; Ramezanian, A. Zinc alleviates salt stress and increase antioxidant enzyme activity in the leaves of pistachio (Pistacia vera L. ‘Badami’) seedings. Turk. J. Agric. For. 2010, 34, 349–359. [Google Scholar]
- Song, C.Z.; Liu, M.Y.; Meng, J.F.; Chi, M.; Xi, Z.M.; Zhang, Z.W. Promoting effect of foliage sprayed zinc sulfate on accumulation of sugar and phenolics in berries of Vitis vinifera cv. Merlot growing on zinc deficient soil. Molecules 2015, 20, 2536–2554. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Yuan, Y.; Liu, Y.Y.; Liu, Y.; Fu, J.J.; Zheng, J.; Wang, G.Y. Gene families of maize glutathione-ascorbate redox cycle respond differently to abiotic stresses. J. Plant Physiol. 2012, 169, 183–192. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, B.; Li, M.J.; Yin, X.M.; Huang, L.F.; Cui, Y.C.; Wang, M.L.; Xia, X. OsMSR15 encoding a rice C2H2-type zinc finger protein confers enhanced drought tolerance in transgenic Arabidopsis. J. Plant Biol. 2016, 59, 271–281. [Google Scholar] [CrossRef]
- Han, G.; Lu, C.; Guo, J.; Qiao, Z.; Sui, N.; Qiu, N.; Wang, B. C2H2 Zinc Finger Proteins: Master Regulators of Abiotic Stress Responses in Plants. Front. Plant Sci. 2020, 11, 115. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Chao, D.Y.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, H.; Maruyama, K.; Sakuma, Y.; Meshi, T.; Iwabuchi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Cys2/His2-type zinc-finger proteins function as transcription repressors under drought, cold, and high-salinity stress conditions. Plant Physiol. 2004, 136, 2734–2746. [Google Scholar] [CrossRef] [Green Version]
- Coca, M.A.; Almoguera, C.; Jordano, J. Expression of sunflower low molecular weight heat shock proteins during embryogenesis and persistence after germination: Localization and possible functional implications. Plant Mol. Biol. 1994, 25, 479–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Heinen, R.B.; Ye, Q.; Chaumont, F. Role of aquaporins in leaf physiology. J. Exp. Bot. 2009, 60, 2971–2985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, H.L.; Yu, X.; Ye, Q.; Ding, X.S.; Kitagawa, Y.; Kwak, S.S.; Su, W.A.; Tang, Z.C. The role of aquaporin RWC3 in drought avoidance in rice. Plant Cell Physiol. 2004, 45, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galmes, J.; Pou, A.; Alsina, M.M.; Tomas, M.; Medrano, H.; Flexas, J. Aquaporin expression in response to different water stress intensities and recovery in Richter-110 (Vitis sp.): Relationship with Eco physiological status. Planta 2007, 226, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Kaldenhoff, R.; Ribas-Carbo, M.; Flexas, J.; Lovisolo, C.; Heckwolf, M.; Uehlein, N. Aquaporins and plant water balance. Plant Cell Environ. 2008, 31, 658–666. [Google Scholar] [CrossRef]
- Danielson, J.Å.H.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol. 2008, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin water channels in mammals. Clin. Exp. Nephrol. 2009, 13, 107–117. [Google Scholar] [CrossRef]
- Henzler, T.; Ye, Q.; Steudle, E. Oxidative gating of water channels (aquaporins) in Chara by hydroxyl radicals. Plant Cell Environ. 2004, 27, 1184–1195. [Google Scholar] [CrossRef]
- Tyerman, S.D.; Bohnert, H.J.; Maurel, C.; Steudle, E.; Smith, J.A.C. Plant aquaporins: Their molecular biology, biophysics and significance for plant water relations. J. Exp. Bot. 1999, 50, 1055–1071. [Google Scholar] [CrossRef] [Green Version]
- Rucinska, S.R. Water relations in plants subjected to heavy metal. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, H.; Zaghdoud, C.; Nortes, P.A.; Carvajal, M.; Martínez-Ballesta, M.D.C. Differential Aquaporin Response to Distinct Effects of Two Zn Concentrations after Foliar Application in Pak Choi (Brassica rapa L.) Plants. Agronomy 2020, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Aroca, R.; Ferrante, A.; Vernieri, P.; Chrispeels, M.J. Drought, abscisic acid and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants. Ann. Bot. 2006, 98, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandeleur, R.K.; Sullivan, W.; Athman, A.; Jordans, C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. Rapid shoot-to-root signalling regulates root hydraulic conductance via aquaporins. Plant Cell Environ. 2014, 37, 520–538. [Google Scholar] [CrossRef]
- Farooq, M.; Ullah, A.; Rehman, A.; Nawaz, A.; Nadeem, A.; Wakeel, A.; Nadeem, F.; Siddique, K.H.M. Application of zinc improves the productivity and biofortification of fine grain aromatic rice grown in dry seeded and puddled transplanted production systems. Field Crop Res. 2018, 216, 53–62. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kadambot, H.; Siddique, M. Micronutrient application through seed treatment. J. Soil Sci. Plant Nutr. 2012, 12, 125–142. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M. Zinc seed coating improves the growth, grain yield and grain biofortification of bread wheat. Acta Physiol. Plant. 2016, 3, 238. [Google Scholar] [CrossRef]
- Grzebisz, W.; Wronska, M.; Diatta, J.B.; Dullin, P. Effect of zinc foliar application at early stages of maize growth on patterns of nutrients and dry matter accumulation by the canopy. Part I. Zinc uptake patterns and its redistribution among maize organs. J. Elem. 2008, 13, 17–28. [Google Scholar]
- Baloch, Q.B.; Chachar, Q.I.; Tareen, M.N. Effect of foliar application of macro and micro nutrients on production of green chillies (Capsicum annuum L.). J. Agric. Technol. 2008, 4, 177–184. [Google Scholar]
- Leach, K.; Hameleers, A. The effects of a foliar spray containing phosphorus and zinc on the development, composition and yield of forage maize. Grass For Sci. 2008, 56, 3113–3115. [Google Scholar] [CrossRef]
- Thalooth, A.T.; Tawfik, M.M.; Mohamed, H.M. A comparative study on the effect of foliar application of zinc, potassium and magnesium on growth, yield and some chemical constituents of mungbean plants grown under water stress conditions. World J. Agric. Sci. 2006, 2, 37–46. [Google Scholar]
- Yadavi, A.; Aboueshaghi, R.S.; Dehnavi, M.M.; Balouchi, H. Effect of micronutrients foliar application on grain qualitative characteristics and some physiological traits of bean (Phaseolus vulgaris L.) under drought stress. Indian J. Fundam. Appl. Life Sci. 2014, 4, 124–131. [Google Scholar]
- Cakmak, I.; Pfeiffer, W.H.; Mcclafferty, B. Biofortification of durum wheat with zinc and iron. Cereal Chem. 2010, 87, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Randhawa, M.A.; Anjum, S.A.; Nadeem, M.; Naeem, M. Exploring the role of zinc in maize (Zea mays L.) through soil and foliar application. Univers. J. Agric. Res. 2015, 3, 69–75. [Google Scholar]
- Khan, H.; Hassan, Z.U.; Maitlo, A.A. Yield and micronutrients content of bread wheat (Triticum aestivum L.) under a multi-nutrient fertilizer-Hal-tonic. Int. J. Agric. Biol. 2006, 8, 366–370. [Google Scholar]

| Protein | Crop/Plant | Function | Reference |
|---|---|---|---|
| ZmZLP1 | Maize | Transport of Zn from ear to cytoplasm | [52] |
| AtHMA1-AtHMA4 | A. thaliana | Involved in export of Zn out of plastids | [53] |
| AhHMA4 | A. halleri | Involved in metal Zn partitioning amid root and shoot | [54] |
| OsYSL13, OsYSL14 | Rice | Enhances Zn translocation from senescing leaves to seed | [55] |
| AtMTP1 and AtMTP3 | A. thaliana | Involved in Zn partitioning among organs | [56] |
| AtPCR2 | A. thaliana | Involved in Zn transport and xylem loading | [57] |
| OsVIT1 and OsV1T2 | Rice | Mediates Zn transport from flag leaf to grain | [58] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umair Hassan, M.; Aamer, M.; Umer Chattha, M.; Haiying, T.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The Critical Role of Zinc in Plants Facing the Drought Stress. Agriculture 2020, 10, 396. https://doi.org/10.3390/agriculture10090396
Umair Hassan M, Aamer M, Umer Chattha M, Haiying T, Shahzad B, Barbanti L, Nawaz M, Rasheed A, Afzal A, Liu Y, et al. The Critical Role of Zinc in Plants Facing the Drought Stress. Agriculture. 2020; 10(9):396. https://doi.org/10.3390/agriculture10090396
Chicago/Turabian StyleUmair Hassan, Muhammad, Muhammad Aamer, Muhammad Umer Chattha, Tang Haiying, Babar Shahzad, Lorenzo Barbanti, Muhammad Nawaz, Adnan Rasheed, Aniqa Afzal, Ying Liu, and et al. 2020. "The Critical Role of Zinc in Plants Facing the Drought Stress" Agriculture 10, no. 9: 396. https://doi.org/10.3390/agriculture10090396
APA StyleUmair Hassan, M., Aamer, M., Umer Chattha, M., Haiying, T., Shahzad, B., Barbanti, L., Nawaz, M., Rasheed, A., Afzal, A., Liu, Y., & Guoqin, H. (2020). The Critical Role of Zinc in Plants Facing the Drought Stress. Agriculture, 10(9), 396. https://doi.org/10.3390/agriculture10090396