3.1. Grain Yield
Maize grain yield showed great variability under the influence of experimental factors and ranged from 8.50 to 11.03 t ha
−1. A detailed assessment of the experimental factors showed significant differences between the tested varieties (
Table 2). Higher yields were recorded for the ES Paroli SG variety and the difference in yield between the varieties was 1.06 t ha
−1. Yields also varied between the research years (
Table 3). The lowest grain yield for both varieties was found in 2010 and it was significantly different from the other two years of the study.
The level of yields also varied depending on the mineral fertilization variant applied. Regardless of the year of research, a significant increase in grain yield was noted in all fertilized objects compared to the control variant (
Table 4 and
Table 5). Significant differences were also found between fertilized objects, which means that the variant of mineral fertilization under the conditions of the experiment significantly influenced the differences in maize grain yield, although its effect varied depending on the variety and was more pronounced in the variety ES Paroli SG. The risk of a lower yield can be reduced by using balanced mineral fertilization of all nutrients, which has been confirmed by the results of the present research recorded for the tested varieties in NPKMgS variants, which amounted to 10 t ha
−1 and 11.26 t ha
−1, respectively. Maize grain yield reflects the state of ingredient imbalance during plant growing period. The amount of the main nutrients used in fertilizers is highly unbalanced both on a global and national scale [
12]. In the last decade, the N:P
2O
5:K
2O ratio was approximately 1:0.3:0.40, while the physiologically determined ratio of these nutrients in the biomass of high-yielding maize at harvest was 1:0.45:1.0 (1.2) [
13]. The role of nitrogen in maize yielding is very well recognized, but at the same time the intensity of the discussion on the yield-forming efficiency of this mineral in maize fertilization does not decrease [
14,
15]. Particularly noteworthy are variants where no phosphorus or potassium was applied (
Table 4 and
Table 5). Lack of fertilization with these components caused a decrease in yield compared to the optimally balanced variant with respect to the nitrogen dose. Lack of phosphorus in the fertilizer dose compared to the variant optimally fertilized with NPKMgS, resulted in a decrease in maize grain yield by an average of 4.1% (ES Palazzo) and 8.7% (ES Paroli SG). The above relationship confirms the general view that maize is a species with specific phosphorus requirements [
16]. According to Shenoy and Kalagudi [
17] insufficient amount of available P may cause a yield reduction in the range from 10 to 15% compared to maximum yield. In the field experiments, maize was grown in sites with low available phosphorus content. Banaj et al. [
18] have indicated that phosphorus fertilization can increase maize grain yield, but on the other hand, excessive application of this ingredient usually does not cause additional yield increases [
19] and can lead to environmental pollution [
20,
21] and contribute to micronutrient deficiencies in maize plants [
22,
23]. Yi et al. [
24] have proved that high maize performance when soil fertility is low would likely depend on other factors such as seeding density and application of additional complementary fertilizers during the growing season, and particularly during the grain formation phase. Huang et al. (1) reported that unbalanced mineral fertilization increased maize yield in a short period of time but had a negative effect on soil fertility in a longer perspective. This regularity has also been confirmed in studies of Gaj [
25] and Bąk and Gaj [
16]. The yield response of maize to the lack of potassium fertilization compared to objects without phosphorus was weaker and showed dependence on the tested variety. The reduction of yield due to the lack of potassium in fertilization was more pronounced in the variety ES Palazzo, for which the decrease in grain yield was 2.4% as compared to the NPKMgS object. The role of potassium in shaping the size of maize yield was emphasized by many researchers while pointing to the importance of utilization of this ingredient from sub-arable layers [
26]. In order to increase the effectiveness of nitrogen, apart from the dose, attention should be paid to other nutritional factors that determine nitrogen utilization from fertilizers, i.e., phosphorus, magnesium and sulfur. In the current study, the highest maize grain yields, regardless of the variety, were recorded in variants optimally balanced in terms of nitrogen (NPKMgS). The introduction of magnesium and sulfur caused a slight increase in grain yield, which was below 1% compared to NPK objects. Salvagiotti et al. [
4] showed that the average grain yield response to S and P addition was approximately 13% and 20%, respectively.
3.2. Assessment of Maize Nutritional Status
It is assumed that the initial growth phase of the 4–5th leaf is important for maize growth and development [
27]. The first ear buds appear in the period from the leaf stage 3 to 6. The assessment of plant nutritional status during vegetation is performed in order to diagnose the current state of mineral nutrition and predict the final yield. The prognostic value increases when the content of components is referenced to their current content in the plant. Nutritional status of plants in the stage of 5–6 developed maize leaves, based on the average ingredient contents from 2009–2011, showed nitrogen malnutrition in control variants of the tested varieties (
Table 4 and
Table 5). Lower nitrogen content compared to normative values was also recorded in NPKMgS objects for the variety ES Palazzo and NK, NMg, NMgS for the variety ES Paroli SG. The remaining variants were characterized by optimal nitrogen and potassium contents compared to the normative values. It can therefore be assumed that maize plants were well nourished with these ingredients at the beginning of the growing season and were prepared for intensive CO
2 assimilation and had sufficient resources of this component, both from soil and fertilizers. An insufficient amount of nitrogen when maize reaches a height of 20 cm reduces the number of grain rows in the ear bud and, consequently, reduces the final yield [
28]. According to Subedi and Ma [
29], plant nitrogen malnutrition before the 6–8 leaf stage leads to an irreversible reduction in the number of ears and potential kernels by up to 30%.
Mineral fertilization significantly differentiated potassium content in maize leaves of the variety ES Paroli SG at the BBCH 1516 stage. Omission of potassium in fertilization (objects: Control, N, NMg, NMgS, NP, NS) resulted in a significant decrease of this component in plants compared to the NPKMgS object (
Table 5). Many literature data [
30,
31,
32] indicate that the genetic factor is an element that significantly differentiates potassium content in plants and not the K dose used in the fertilizer. Askegaard et al. [
33] emphasized that the assessment of potassium content in plants is a key factor in the effective management of this element and is a complement to soil tests. The analyzed varieties were cultivated in conditions of low abundance in available potassium (87.9–165.2 mg kg
−1). Every agricultural process or procedure that interferes with potassium plant nutrition reduces the plant’s metabolic activity, and thus contributes to the reduction of nitrogen yield-forming efficiency. Moreover, Yang et al. [
32] has emphasized that potassium content in maize organs was significantly dependent on the cultivation system and application technique of this ingredient. Maize is a very sensitive plant to potassium supply, especially in the period from 5/6 leaves to flowering. Appropriate plant nutrition in the critical growth phase results in a reduced risk of lodging (better developed mechanical tissue), higher grain number in the ear, higher weight of kernels (longer grain filling stage), even maturation and lower water content during the maturation stage [
34].
Irrespective of the analyzed variety, a significant deficiency of phosphorus, magnesium and calcium was noted in maize leaves (
Table 4 and
Table 5). Except for the control object, the plants were well-fed with nitrogen. Maize nutrition with phosphorus, magnesium and calcium was significantly below the critical value, which in the case of phosphorus is 4.0 g kg
−1, 3–6 g for Mg kg
−1 and 5–16 g for Ca kg
−1 [
35]. At this stage of development, phosphorus determines the rate of growth of the root system and thus the plant’s ability to absorb water and nutrients from the soil. In the period from emergence to the BBCH-12 leaf stage, maize builds a root system, and the basic factors determining the conditions of its formation is the availability of phosphorus as well as the lack of limiting factors. Modern approach to fertilization requires balancing all ingredients, including secondary ones. Magnesium and sulfur play a specific role in this context. In addition to the specific physiological functions in the plant, both elements play an important role in nitrogen metabolism [
36]. In the BBCH 15/16 stage, no significant differences were found in magnesium and calcium content under the influence of variable levels of mineral fertilization. Calcium and magnesium content was 1.01–2.05 Ca g·kg
−1 and 2.0–2.7 Mg g·kg
−1, respectively (
Table 1). The obtained contents of ingredients, regardless of the analyzed experimental variant, were significantly below the normative values (Ca: 5.1–16 g·kg
−1; Mg: 3–6 g·kg
−1) set by Schulte and Kelling [
37]. The causes of Ca and Mg deficiency in plants can be complex and result from both a deficiency of soil components, acidic soil reaction, excess of other cations as well as disruption of their uptake and translocation in the plant during develops [
38,
39]. Literature data shows that intensive potassium fertilization can sometimes affect the content not only of potassium [
40,
41], but also other ingredients in crop yields, causing their excessive or deficient concentration, especially in relation to two elements i.e., calcium and magnesium [
42]. The issue of interaction between potassium and magnesium has been discussed in many studies [
43,
44,
45]. Magnesium ions passively translocate through the cytoplasmic membrane with the electrochemical gradient of cation concentration and are first exposed to competition with actively taken up potassium ions [
46]. Secondly, Mg
2+ ions compete with calcium ions, which are also passively taken up by the plant [
47].
A comprehensive analysis of the assessment taking into account the dependence of maize grain yield of the tested varieties on the produced biomass of plants in the BBCH 15/16 stage and component contents (N, P, K, Ca, Mg, Na) in leaves and N:K, N:P and N:Mg ratios was performed using multiple retrograde linear regression. Regression analysis showed that a strong relationship between the yield size and produced biomass as well as calcium content and N:K ratio was found in the variety ES Palazzo at the initial growth stage (BBCH 15/16). These variables determined the yield of ES Palazzo maize in 69%. The above relationship is expressed by the following regression Equation (1):
Significant codes—***: p-value < 0.001; **: p-value < 0.01.
In the present study, maize was grown on acidic soil (pH 5.1–5.4), which in consequence resulted in a low Mg and Ca content in the plant at the early developmental stage. Low calcium and magnesium content in plants at the BBCH 15/16 stage showed a significant impact on the grain yield and this relationship was confirmed by regression Equations (1) and (2).
As regards the second variety ES Paroli SG, the yield was largely shaped by plant biomass and the content of potassium (K), calcium (Ca), magnesium and N:Mg ratio (Equation (2)). These variables determined maize grain yield in 59.7%.
Significant codes—***: p-value < 0.001; *: p-value < 0.05.
The relationships found prove that the more disturbed nutritional homeostasis, the greater the role of the analyzed ingredients in shaping the yield. The content of individual components changes with plant age, thus the relationships between the components that provide more information about the physiological state of plants are more useful for diagnostic purposes [
48,
49]. Confirmation of this thesis were path analyses (
Figure 2 and
Figure 3), which indicated a number of interactions between ingredients in maize leaves at the BBCH15/16 stage. In the case of the variety ES Paroli SG, interactions between Ca and Mg, Ca and N:Mg, Ca and N:P and Ca and Na were found. For the variety EC Palazzo, relationships between P and Mg, P and Ca and N:Mg and Na were observed. In addition, it should be noted that the yield of the variety ES Paroli SG (
Figure 2) was mainly influenced by the content of potassium, phosphorus and N:K and N:P ratios, while nitrogen content and N:Mg ratio had a significantly lower effect. Yield of the variety ES Palazzo (
Figure 3) was most strongly dependent on potassium content and N:K ratio, while the content of nitrogen, phosphorus, N:P ratio and plant biomass in the BBCH 15/16 phase had a lower effect. The issue of interaction between nitrogen and phosphorus has been raised in many works [
50,
51,
52,
53] emphasized that the prolonged phosphorus deficiency in the plant reduced the ATP energy pool and, as a result, nitrate nitrogen (N-NO
3) uptake was reduced. Excess nitrogen, during the lack of phosphorus, causes the first sign of phosphorus deficiency.
3.3. Component Accumulation in Plants at the BBCH 15/16 Stage
The accumulation of components in maize biomass at the BBCH 15/16 stage was significantly dependent on the action of experimental factors (
Table 6). Regardless of the experimental variant, an increase in the accumulation of macronutrients under the influence of mineral fertilization was found in comparison to the control object, but significant differences were noted only in phosphorus and potassium accumulation for the variety ES Palazzo and calcium and magnesium for the variety ES Paroli SG (
Table 7 and
Table 8). Nitrogen needs are minimal in the initial period of maize growth, they increase at a later stage of development and reach maximum between the beginning of flowering and grain formation [
54]. Until the 6–8 leaf stage, maize absorbs about 3% of the total amount of nitrogen, while 85% of this component is taken up from the 6–8 leaf stage to the silk drying stage. Potassium removal from fertilization resulted in a significantly greater reduction in the accumulation of this component in the biomass of ES Palazzo plants compared to ES Paroli SG. A significant decrease in potassium uptake compared to the optimally fertilized variant NPKMgS was observed in N and NS positions for both varieties, but significant differences were found only for the variety ES Palazzo and on average they amounted to 36% for the mentioned objects (
Table 7).
Particular attention should be paid to NMg objects for both tested cultivars, where the highest reduction in leaf potassium accumulation was noted, which indicated antagonistic effect of the components.
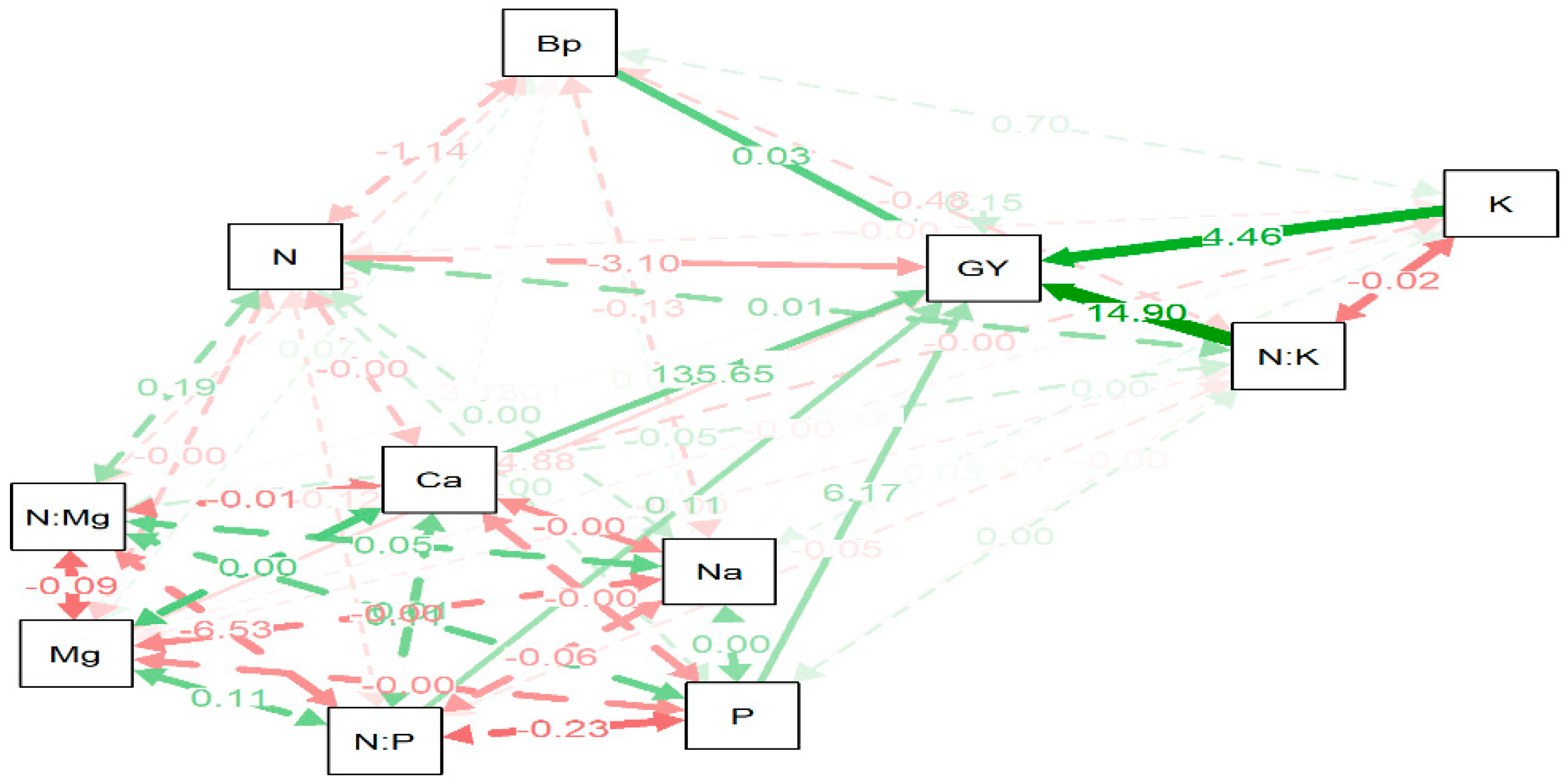
Analysis of the path diagram (
Figure 4) showing the dependence of ES Paroli SG grain yield on component accumulation in leaf biomass at the BBCH 15/16 stage indicated that grain yield largely depended on the accumulation of nitrogen, potassium and calcium in the leaves. In addition, strong interactions between phosphorus, sodium, potassium, nitrogen and magnesium accumulation were found.
In the case of the second variety tested, path analysis (
Figure 5) expressing the dependence of maize grain yield on component accumulation in the BBCH 15/16 stage showed that the yield largely depended on the accumulation of potassium, magnesium and calcium in the leaves in the initial growth phase. Strong interactions have also been noted between the accumulation of phosphorus and nitrogen and potassium and sodium, as well as between the accumulation of potassium and phosphorus, magnesium and sodium (
Figure 5).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}