
Rootstock Affects Floral Induction in Citrus Engaging the Expression of the FLOWERING LOCUS T (CiFT)



 ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
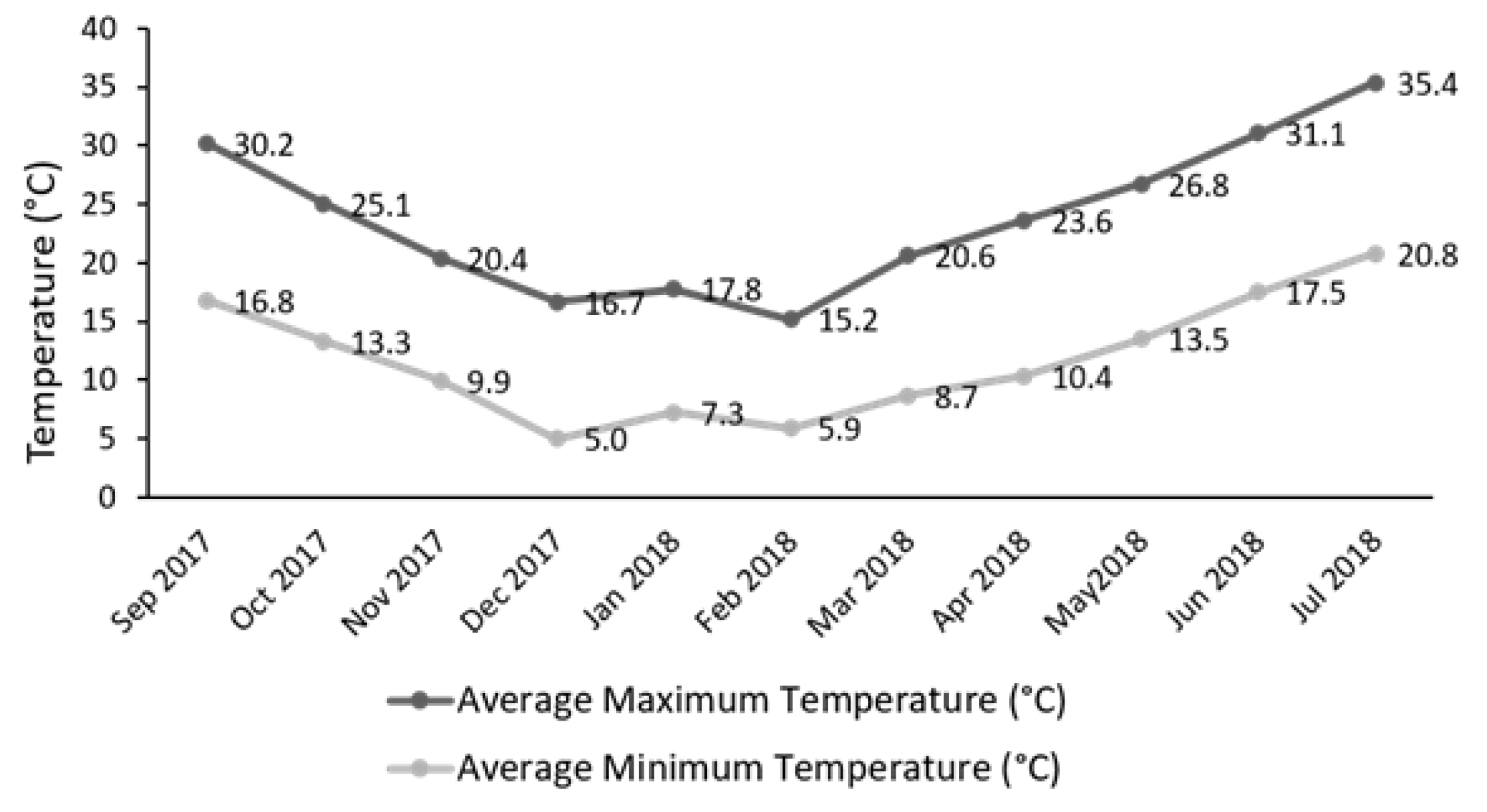
2.1. Plant Material
2.2. Harvest Data and Sampling
2.3. Total RNA Extraction and cDNA Synthesis
2.4. Real Time-PCR
2.5. Statistical Analysis
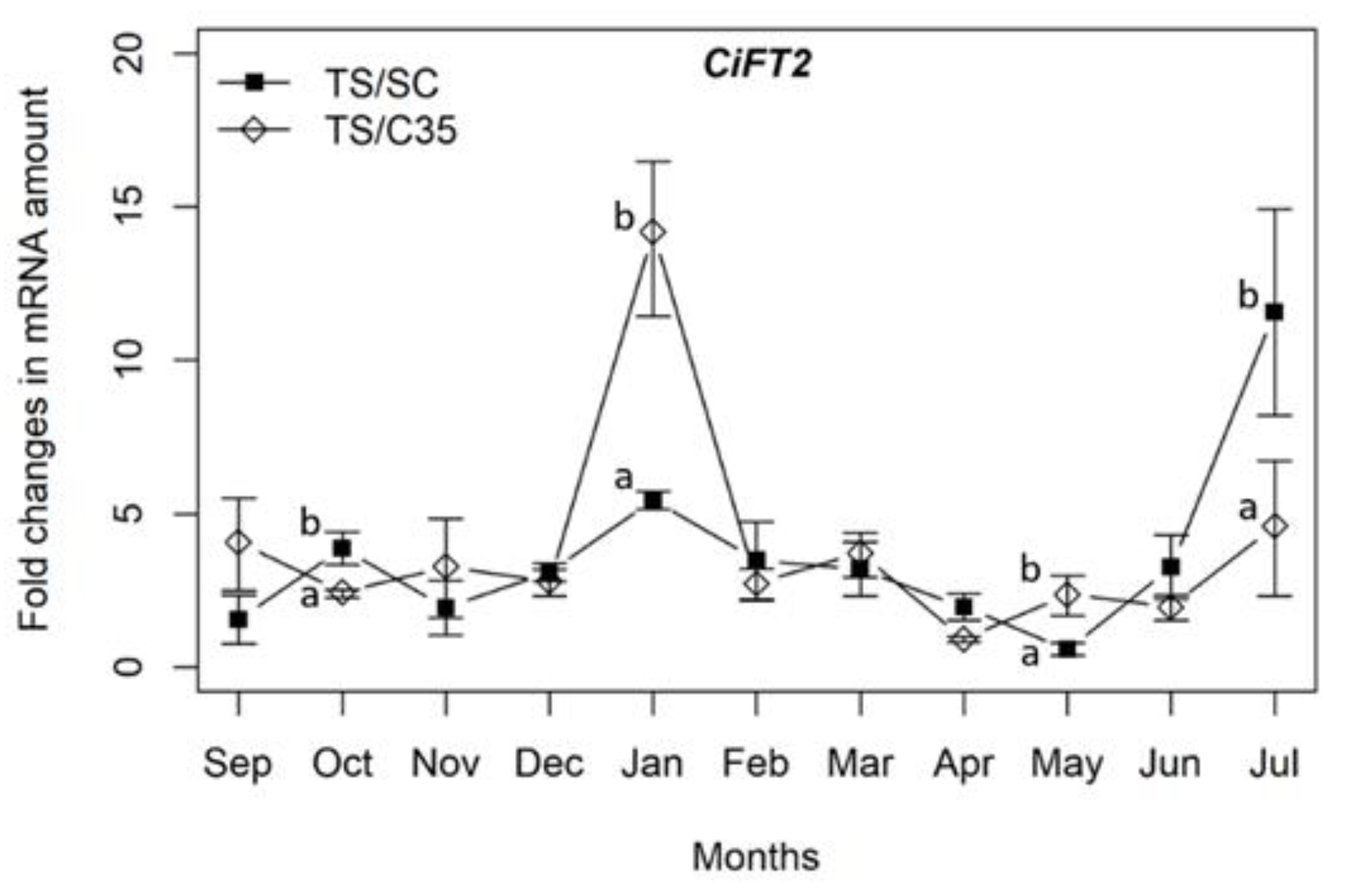
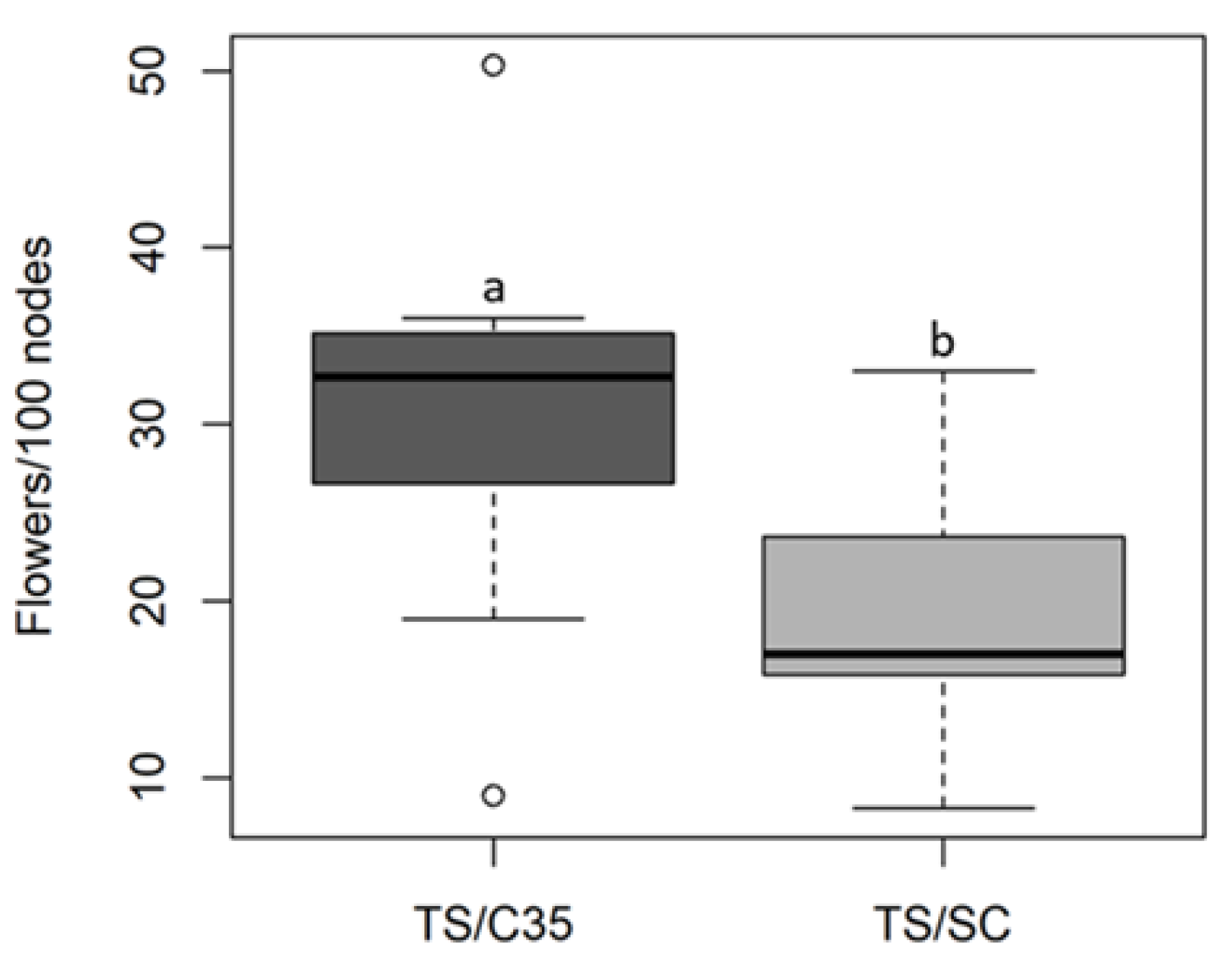
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Available online: http://faostat.fao.org/default.aspx (accessed on 30 April 2018).
- Goldschmidt, E.; Monselise, S. Hormonal control of flowering in citrus trees and other woody perennials. In PlantGrowth Substances; Carr, D.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1972; pp. 758–766. [Google Scholar]
- Spiegel-Roy, P.; Goldschmidt, E.E. Biology of Citrus; Cambridge University Press: New York, NY, USA, 1996. [Google Scholar]
- Moss, G.I. Influence of temperature and photoperiod on flower induction and inflorescence development in sweet orange (Citrus sinensis L. Osbeck). Hortic. Res. 1969, 44, 311–320. [Google Scholar]
- Cassin, J.; Bourdeuet, J.; Fougue, A.; Furon, V.; Gailland, J.; Bourdelles, P.; Montaguad, G.; Moreuil, C. The influence of climate upon the blooming of citrus in tropical areas. Proc. First Int. Citrus Symp. 1969, 1, 315–323. [Google Scholar]
- Garcia-Luis, A.; Kanduser, M.; Santamarina, P.; Guardiola, J.L. Low temperature influence on flowering in Citrus. The separation of inductive and bud dormancy releasing factors. Physiol. Plant. 1992, 86, 648–652. [Google Scholar] [CrossRef]
- Southwick, S.M.; Davenport, T.L. Characterization of water stress and low temperature effects on flower induction in Citrus. Plant. Physiol. 1986, 81, 26–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.R.G.; Ai, X.-Y.; Zhang, J.-Z. Genetic regulation of flowering time in annual and perennial. WIREs RNA 2014, 5, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Komeda, Y. Genetic regulation of time to flower in Arabidopsis thaliana. Annu. Rev. Plant. Biol. 2004, 55, 521–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albani, M.C.; Coupland, G. Comparative analysis of flowering in annual and perennial plants. Curr. Top. Dev. Biol. 2010, 91, 323–348. [Google Scholar] [CrossRef]
- Pin, P.A.; Ove, N. The multifaceted roles of FLOWERING LOCUS T in plant development. Plant. Cell Environ. 2012, 35, 1742–1755. [Google Scholar] [CrossRef]
- Dornelas, M.; Camargo, R.; Figueiredo, L.; Takita, M. A genetic framework for flowering-time pathways in Citrus spp. Genet. Mol. Biol. 2007, 30, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, K.; Agüero, J.; Vives, M.; Aleza, P.; Pina, J.; Moreno, P.; Navarro, L.; Guerri, J. Precocious flowering of juvenile citrus induced by a viral vector based on Citrus leaf blotch virus: A new tool for genetics and breeding. Plant. Biotechnol. J. 2016, 14, 1976–1985. [Google Scholar] [CrossRef] [Green Version]
- Endo, T.; Shimada, T.; Fujii, H.; Kobayashi, Y.; Araki, T.; Omura, M. Ectopic expression of an FT homolog from Citrus confers an early flowering phenotype on trifoliate orange (Poncirus trifoliata L. Raf.). Transgenic Res. 2005, 14, 703–712. [Google Scholar] [CrossRef]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Kobayashi, Y.; Araki, T.; Omura, M. Transcriptional changes in CiFT-introduced transgenic trifoliate orange (Poncirus trifoliata L. Raf.). Tree Physiol. 2010, 30, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, E.; Alquézar, B.; Rodríguez, A.; Martorell, P.; Genovés, S.; Ramón, D.; Zacarías, L.; Peña, L. Metabolic engineering of β-carotene in orange fruit increases its in vivo antioxidant properties. Plant. Biotechnol. J. 2014, 12, 17–27. [Google Scholar] [CrossRef]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Omura, M.; Ikoma, Y. Increased CiFT abundance in the stem correlates with floral induction by low temperature in Satsuma mandarin (Citrus unshiu Marc.). J. Exp. Bot. 2007, 58, 3915–3927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samach, A. Congratulations, you have been carefully chosen to represent an important developmental regulator! Ann. Bot. 2013, 111, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Pillitteri, L.J. Isolation and Characterization of a TERMINAL FLOWER Homolog and Its Correlation with Juvenility in Citrus. Plant. Physiol. 2004, 135, 1540–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chica, E.; Albrigo, L. Changes in CsFT Transcript Abundance at the Onset of Low-temperature Floral Induction in Sweet Orange. J. Am. Soc. Hortic. Sci. 2013, 138, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Chica, E.; Albrigo, L. Expression of flower promoting genes in sweet orange during floral inductive water deficits. J. Am. Soc. Hortic. Sci. 2013, 138, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef]
- Huang, T.; Böhlenius, H.; Eriksson, S.; Parcy, F.; Nilsson, O. The mRNA of the arabidopsis gene FT moves from leaf to shoot apex and induces flowering. Science 2005, 309, 1694–1696. [Google Scholar] [CrossRef] [Green Version]
- Wigge, P.; Kim, M.; Jaeger, K.; Busch, W.; Schmid, M.; Lohmann, J.; Weigel, D. Integration of spatial and temporal information during floral induction in Arabidopsis. Science 2005, 309, 1056–1059. [Google Scholar] [CrossRef]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Moss, G.I. Effect of fruit on flowering in relation to biennial bearing in sweet orange (Citrus sinensis). Hortic. Sci. 1971, 46, 177–184. [Google Scholar] [CrossRef]
- Muñoz-Fambuena, N.; Mesejo, C.; Carmen González-Mas, M.; Primo-Millo, E.; Agustí, M.; Iglesias, D.J. Fruit regulates seasonal expression of flowering genes in alternate-bearing “Moncada” mandarin. Ann. Bot. 2011, 108, 511–519. [Google Scholar] [CrossRef]
- Shalom, L.; Samuels, S.; Zur, N.; Shlizerman, L.; Zemach, H.; Weissberg, M. Alternate bearing in Citrus: Changes in the expression of flowering control genes and in global gene expression in ON- versus OFF-crop trees. PLoS ONE 2012, 7, e46930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Fuentes, A.; Mesejo, C.; Reig, C.; Agustí, M. Timing of the inhibitory effect of fruit on return bloom of ‘Valencia’ sweet orange Citrus sinensis (L.) Osbeck). J. Sci. Food Agric. 2010, 90, 1936–1943. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fambuena, N.; Mesejo, C.; González-Mas, M.; Iglesias, D.; Primo-Millo, E.; Agustí, M. Gibberellic acid reduces flowering intensity in sweet orange [Citrus sinensis (L.) Osbeck] by repressing CiFT gene expression. J. Plant. Growth Regul. 2012, 31, 529–536. [Google Scholar] [CrossRef]
- Goldberg-Moeller, R.; Shalom, L.; Shlizerman, L.; Samuels, S.; Zur, N.; Ophir, R.; Blumwald, E.; Adka, A. Effects of gibberellin treatment during flowering induction period on global gene expression and the transcription of flowering-control genes in Citrus buds. Plant. Sci. 2013, 198, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Shimada, T.; Nakata, Y.; Fujii, H.; Matsumoto, H.; Nakajima, N.; Ikoma, Y.; Omura, M. Abscisic acid affects expression of citrus FT homologs upon floral induction by low temperature in Satsuma mandarin (Citrus unshiu Marc.). Tree Physiol. 2017, 38, 755–771. [Google Scholar] [CrossRef] [PubMed]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock—scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Koepke, T.; Dhingra, A. Rootstock scion somatogenetic interactions in perennial composite plants. Plant. Cell Rep. 2013, 32, 1321–1337. [Google Scholar] [CrossRef]
- Foster, T.M.; Watson, A.E.; Van Hooijdonk, B.M.; Schaffer, R.J. Key flowering genes including FT-like genes are upregulated in the vasculature of apple dwarfing rootstocks. Tree Genet. Genomes 2014, 10, 189–202. [Google Scholar] [CrossRef]
- Sharma, N.; Kumar, S.; Kumar, A.; Ravishankar, H.; Dubey, A.K.; Kumar, N. Physiological and molecular basis of alternate bearing in perennial fruit crops. Sci. Hortic. 2019, 243, 214–225. [Google Scholar] [CrossRef]
- Webster, A.D. Rootstock and interstock effects on deciduous fruit tree vigour, precocity, and yield productivity. N. Z. J. Crop. Hortic. Sci. 2010, 23, 373–382. [Google Scholar] [CrossRef]
- Moreno, P.; Ambrós, S.; Albiach-Martí, M.R.; Guerri, J.; Peña, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant. Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Continella, A.; Pannitteri, C.; Malfa, S.L.; Legua, P.; Distefano, G.; Nicolosi, E.; Gentile, A. Influence of different rootstocks on yield precocity and fruit quality of ‘Tarocco Scirè’ pigmented sweet orange. Sci. Hortic. 2018, 230, 62–67. [Google Scholar] [CrossRef]
- Louzada, E.; del Rio, H.; Setamou, M.; Watson, J.; Swietlik, D. Evaluation of citrus rootstocks for the high pH, calcareous soils of South Texas. Euphytica 2008, 164, 13–18. [Google Scholar] [CrossRef]
- Reforgiato-Recupero, G.; Russo, G.; Recupero, S.; Zurru, R.; Deidda, B.; Mulas, M. Horticultural Evaluation of New Citrus latipes Hybrids as Rootstocks for Citrus. HortScience 2009, 44, 595–598. [Google Scholar] [CrossRef] [Green Version]
- Legua, P.; Forner, J.; Hernández, F.; Forner-giner, M. Total phenolics, organic acids, sugars and antioxidant activity of mandarin (Citrus clementina Hort. ex Tan.): Variation from rootstock rootstock. Sci. Hortic. 2014, 174, 60–64. [Google Scholar] [CrossRef]
- Fu, S.; Shao, J.; Zhou, C.; Hartung, J.S. Transcriptome analysis of sweet orange trees infected with ‘Candidatus Liberibacter asiaticus ’ and two strains of Citrus Tristeza Virus. BMC Genom. 2016, 17, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Distefano, G.; Cirilli, M.; Las Casas, G.; La Malfa, S.; Continella, A.; Rugini, E.; Thomas, B.; Long, G.; Gentile, A.; Muleo, R. Ectopic expression of Arabidopsis phytochrome B in Troyer citrange affects photosynthesis and plant morphology. Sci. Hortic. 2013, 159, 1–7. [Google Scholar] [CrossRef]
- Distefano, G.; Caruso, M.; La Malfa, S.; Gentile, A.; Tribulato, E. Histological and molecular analysis of pollen–pistil interaction in clementine. Plant. Cell Rep. 2009, 28, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017.
- Amasino, R. Seasonal and developmental timing of flowering. Plant. J. 2010, 61, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, F. Regulation of floral induction in Citrus. J. Jpn. Soc. Hortic. 2013, 82, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fuentes, A.; Mesejo, C.; Juan, M.; Almela, V.; Agustí, M. Restrictions on the exogenous control of flowering in Citrus. Acta Hortic. 2004, 632, 91–98. [Google Scholar] [CrossRef]
- Inoue, H. Effects of soil drought and temperature on flower bud differentiation of satsuma mandarin. J. Jpn. Soc. Hortic. Sci. 1989, 58, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Omura, M. Differences in seasonal expression of flowering genes between deciduous trifoliate orange and evergreen satsuma mandarin. Tree Physiol. 2009, 29, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajon, M.; Febres, V.J.; Moore, G.A. Expression patterns of flowering genes in leaves of “Pineapple” sweet orange [Citrus sinensis (L.) Osbeck] and pummelo (Citrus grandis Osbeck). BMC Plant. Biol. 2017, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, F.; Iwasaki, M.; Fukamachi, H.; Nonaka, K.; Imai, A.; Takishita, F.; Yano, T.; Endo, T. Fruit bearing suppresses citrus FLOWERING LOCUS T expression in vegetative shoots of satsuma mandarin (Citrus unshiu Marc.). J. Jpn. Soc. Hortic. Sci. 2012, 81, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, F.; Iwasaki, M.; Fukamachi, H.; Endo, T. Predicting the number of flowers in Satsuma Mandarin (Citrus unshiu Marc.) trees based on Citrus FLOWERING LOCUS T mRNA levels. Hortic. J. 2017, 86, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Agustí, M.; Mesejo, C.; Muñoz-Fambuena, N.; Vera-Sirera, F.; de Lucas, M.; Martinez-Fuentes, A.; Reig, C.; Iglesias, D.J.; Primo-Millo, E.; Miguel, A. Fruit-dependent epigenetic regulation of flowering in Citrus. New Phytol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Almansa, M.; Hernandez, J.; Jimenez, A.; Botella, M.; Sevilla, F. Effect of salt stress on the superoxide dismutase activity in leaves of Citrus limonum in different rootstock–scion combinations. Biol. Plant. 2002, 45, 545–549. [Google Scholar] [CrossRef]
- Jensen, P.; Rytter, J.; Detwiler, E.; Travis, J.; McNellis, T. Rootstock effects on gene expression patterns in apple tree scions. Plant. Mol. Biol. 2003, 53, 493–511. [Google Scholar] [CrossRef]
- Prassinos, C.; Ko, J.; Lang, G.; Iezzoni, A.; Han, K. Rootstock-induced dwarfing in cherries is caused by differential cessation of terminal meristem growth and is triggered by rootstock-specific gene regulation. Tree Physiol. 2009, 29, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Putterill, J.; Varkonyi-Gasic, E. FT and florigen long-distance flowering control in plants. Curr. Opin. Plant. Biol. 2016, 33, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lifschitz, E.; Eviatar, T.; Rozman, A.; Shalit, A.; Goldshmidt, A.; Amsellem, Z.; Alvarez, J.; Eshed, Y. The tomato FT ortholog triggers systemic signals that regulate growth and flowering and substitute for diverse environmental stimuli. Proc. Natl. Acad. Sci. USA 2006, 103, 6398–6403. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-K.; Belanger, H.; Lee, Y.-J.; Varkonyi-Gasic, E.; Taoka, K.-I.; Miura, E.; Xoconostle-Cázares, B.; Gendler, K.; Jorgensen, R.A.; Phinney, B.; et al. FLOWERING LOCUS T protein may act as the long-distance florigenic signal in the cucurbits. Plant. Cell Online 2007, 19, 1488–1506. [Google Scholar] [CrossRef] [Green Version]
- Notaguchi, M.; Abe, M.; Kimura, T.; Daimon, Y.; Kobayashi, T.; Yamaguchi, A.; Tomita, Y.; Dohi, K.; Mori, M.; Araki, T. Long-distance, grafttransmissible action of Arabidopsis FLOWERING LOCUS T protein to promote flowering. Plant. Cell Physiol. 2008, 49, 1645–1658. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Geng, Y.; Zhang, B.; Mao, H.; Qu, J.; Chua, N.-H. The Jatropha FT ortholog is a systemic signal regulating growth and flowering time. Biotechnol. Biofuels 2014, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, J.; Liu, M.; Yao, Q.; Chen, J. Transcriptome Profiling to Understand the Effect of Citrus Rootstocks on the Growth of ‘Shatangju’ Mandarin. PLoS ONE 2017, 2013, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Lovatt, C.J. Effects of low temperature and gibberellic acid on floral gene expression and floral determinacy in ‘Washington ’navel orange (Citrus sinensis L. Osbeck). Sci. Hortic. 2019, 243, 92–100. [Google Scholar] [CrossRef]
- Collani, S.; Neumann, M.; Yant, L.; Schmid, M. FT Modulates Genome-Wide DNA-Binding of the bZIP Transcription Factor FD. Plant. Physiol. 2019, 180, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, J.M.; Weber, K.C.; Qiu, W.; Stanton, D.; Mahmoud, L.M.; Wu, H.; Huyck, P.; Zale, J.; Jasim, K.A.; Grosser, J.W.; et al. The vascular targeted citrus FLOWERING LOCUS T3 gene promotes non-inductive early flowering in transgenic Carrizo rootstocks and grafted juvenile scions. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennici, S.; Las Casas, G.; Distefano, G.; Gentile, A.; Lana, G.; Di Guardo, M.; Nicolosi, E.; La Malfa, S.; Continella, A. Rootstock Affects Floral Induction in Citrus Engaging the Expression of the FLOWERING LOCUS T (CiFT). Agriculture 2021, 11, 140. https://doi.org/10.3390/agriculture11020140
Bennici S, Las Casas G, Distefano G, Gentile A, Lana G, Di Guardo M, Nicolosi E, La Malfa S, Continella A. Rootstock Affects Floral Induction in Citrus Engaging the Expression of the FLOWERING LOCUS T (CiFT). Agriculture. 2021; 11(2):140. https://doi.org/10.3390/agriculture11020140
Chicago/Turabian StyleBennici, Stefania, Giuseppina Las Casas, Gaetano Distefano, Alessandra Gentile, Giuseppe Lana, Mario Di Guardo, Elisabetta Nicolosi, Stefano La Malfa, and Alberto Continella. 2021. "Rootstock Affects Floral Induction in Citrus Engaging the Expression of the FLOWERING LOCUS T (CiFT)" Agriculture 11, no. 2: 140. https://doi.org/10.3390/agriculture11020140
APA StyleBennici, S., Las Casas, G., Distefano, G., Gentile, A., Lana, G., Di Guardo, M., Nicolosi, E., La Malfa, S., & Continella, A. (2021). Rootstock Affects Floral Induction in Citrus Engaging the Expression of the FLOWERING LOCUS T (CiFT). Agriculture, 11(2), 140. https://doi.org/10.3390/agriculture11020140