
Phenolic Response to Walnut Anthracnose (Ophiognomonia leptostyla) Infection in Different Parts of Juglans regia Husks, Using HPLC-MS/MS



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extraction of the Individual Phenolic Compounds
2.3. HPLC–Mass Spectrometry Analysis of Individual Phenolic Compounds
2.4. Chemicals
2.5. Statistical Analysis
3. Results and Discussion
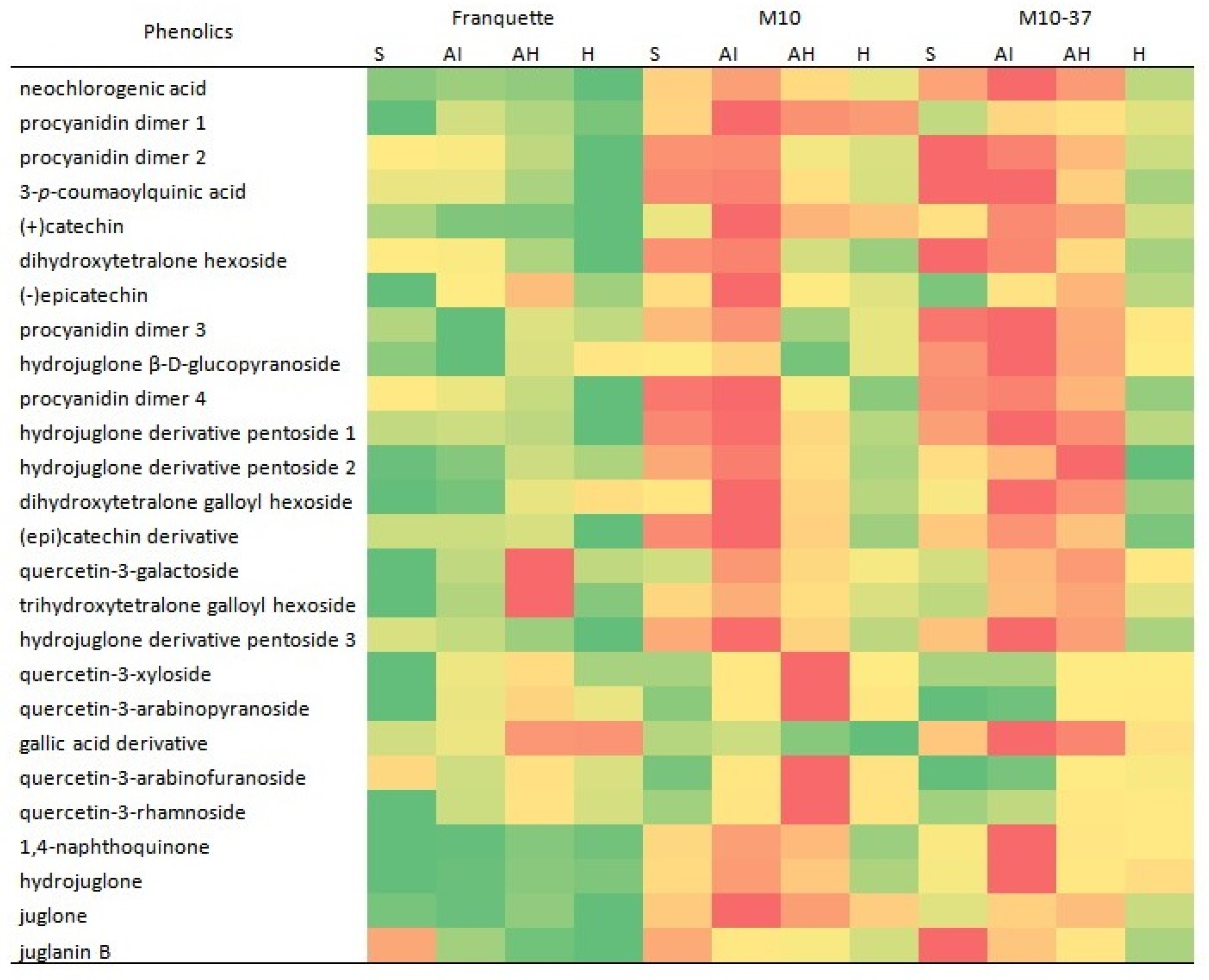
3.1. Identification of Individual Phenolics
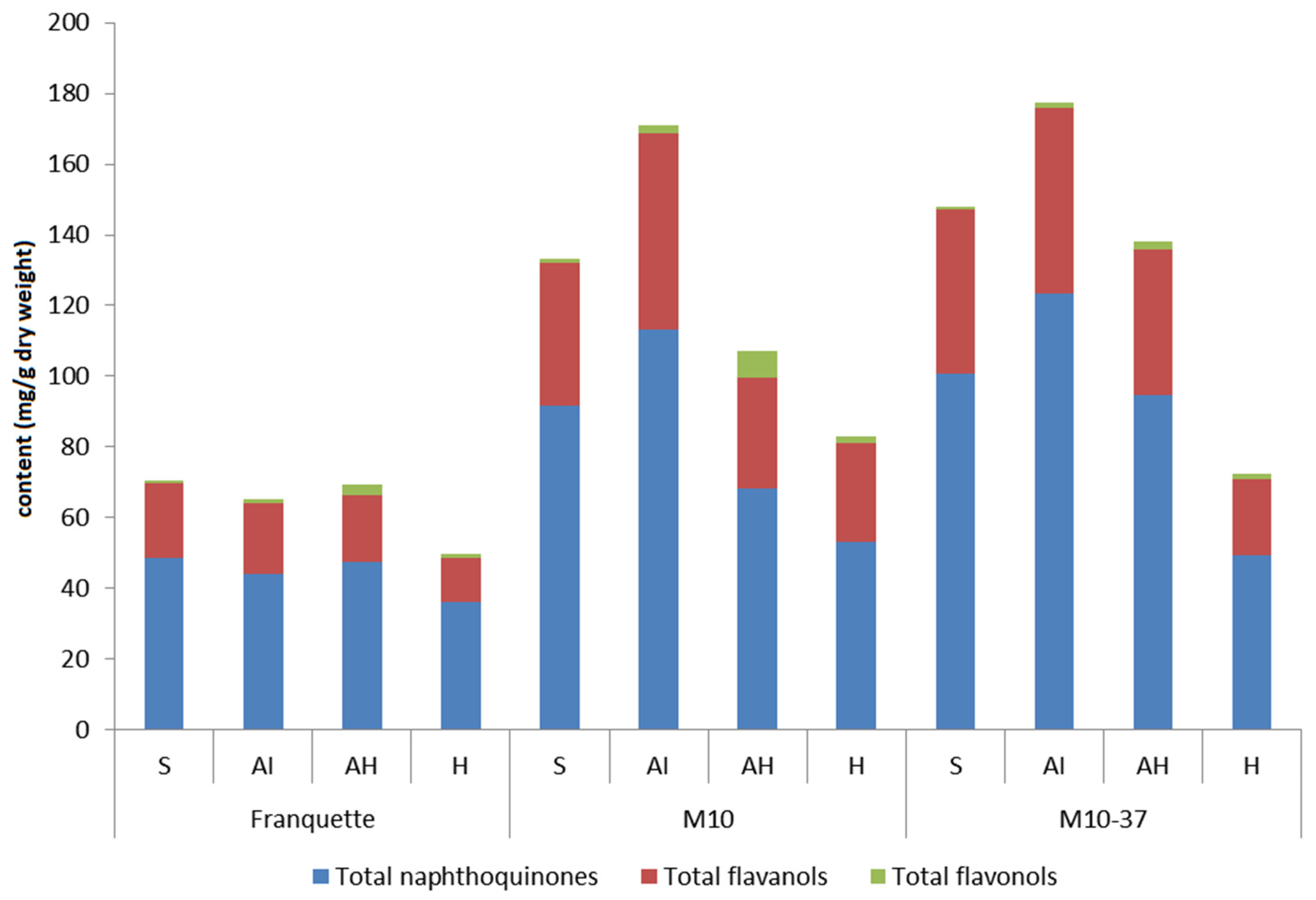
3.2. Phenolic Groups in Response to Walnut Anthracnose Infection
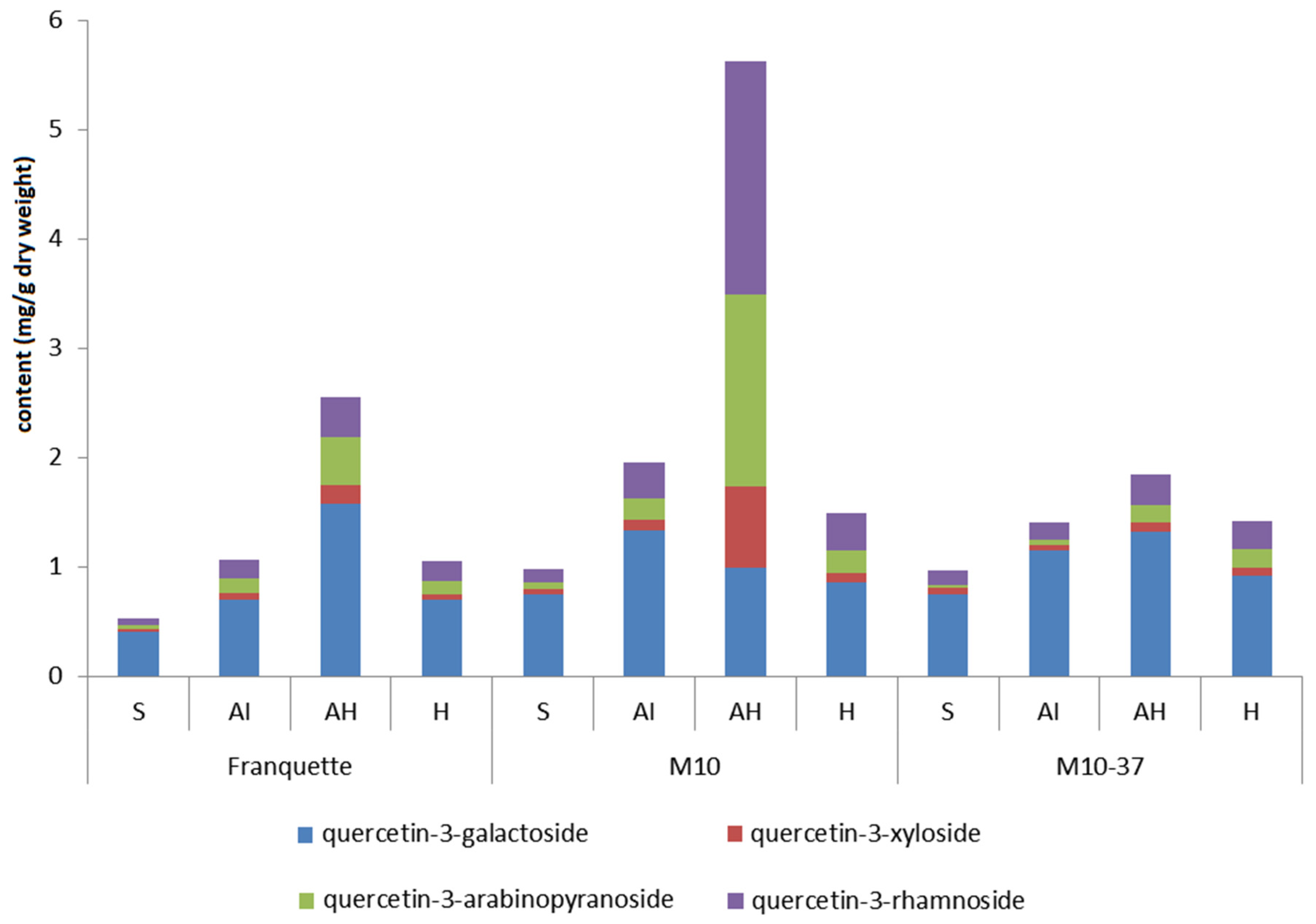
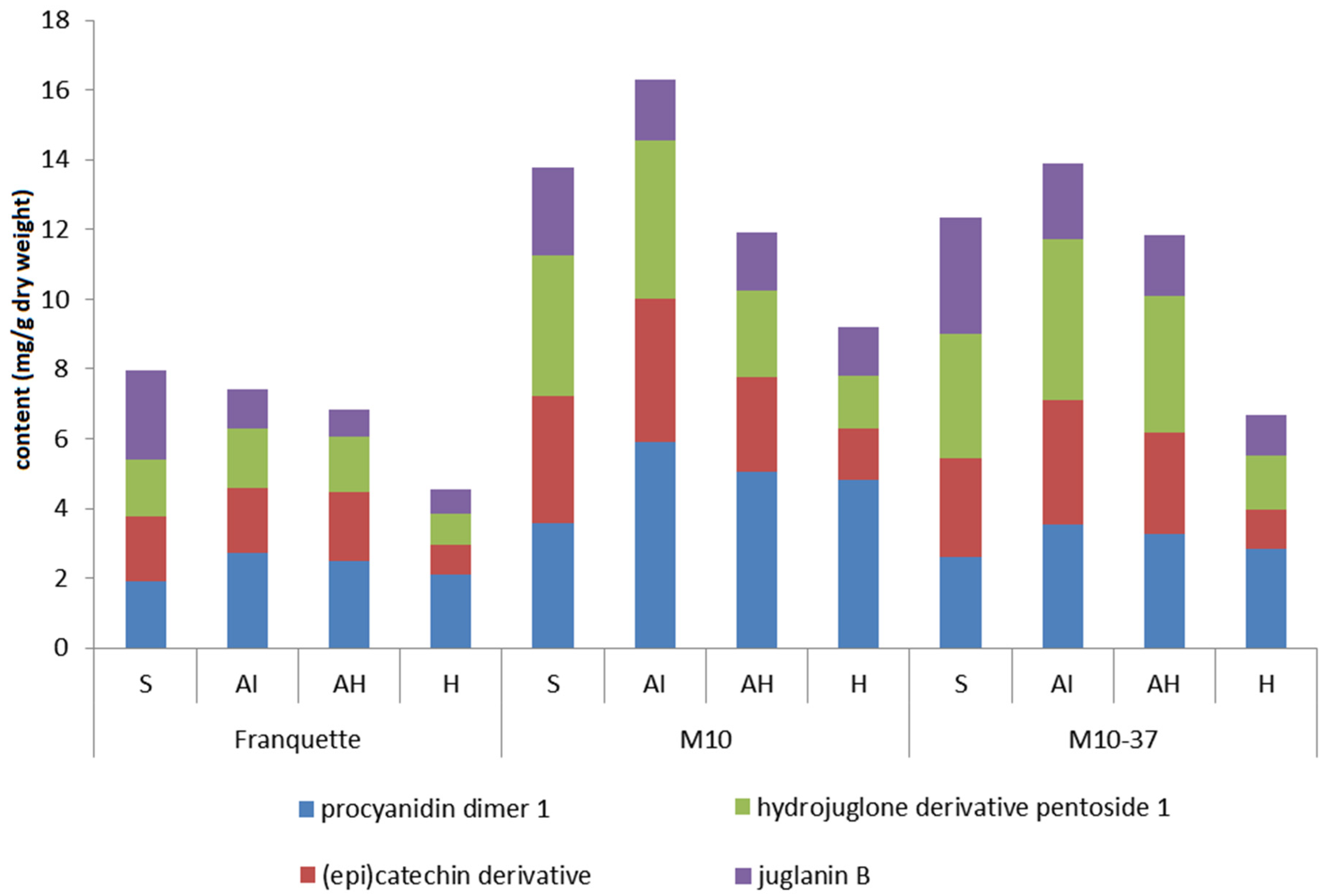
3.3. Individual Phenolic Compounds in Response to Walnut Anthracnose Infection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pollegioni, P.; Van der Linden, G.; Belisario, A.; Gras, M.; Anselmi, N.; Olimpieri, I.; Luongo, L.; Santini, A.; Turco, E.; Scarascia Mugnozza, G.; et al. Mechanisms governing the responses to anthracnose pathogen in Juglans spp. J. Biotechnol. 2012, 159, 251–264. [Google Scholar] [CrossRef]
- Yang, H.; Cao, G.; Jiang, S.; Han, S.; Yang, C.; Wan, X.; Zhang, F.; Chen, L.; Xiao, J.; Zhu, P.; et al. Identification of the anthracnose fungus of walnut (Juglans spp.) and resistance evaluation through physiological responses of resistant vs. susceptible hosts. Plant Pathol. 2021, 70, 1219–1229. [Google Scholar] [CrossRef]
- Belisario, A.; Scotton, M.; Santori, A.; Onofri, S. Variability in the Italian population of Gnomonia leptostyla, homothallism and resistance of Juglans species to anthracnose. For. Pathol. 2008, 38, 129–145. [Google Scholar] [CrossRef]
- Woeste, K.E.; Beineke, W.F. An efficient method for evaluating black walnut for resistance to walnut anthracnose in field plots and the identification of resistant genotypes. Plant Breed. 2001, 120, 454–456. [Google Scholar] [CrossRef]
- Arnaudov, V.A.; Gandev, S.I. Susceptibility of some walnut cultivars to Gnomonia Leptostila. Acta Hortic. 2009, 825, 407–412. [Google Scholar] [CrossRef]
- Belisario, A.; Forti, E.; Cichello, A.M.; Zoina, A.; Barbieri, E.; Valier, A. Epidemilogical surveys of Gnomonia leptostyla in Juglans regia hedgerow trained orchard. Acta Hortic. 2001, 544, 405–408. [Google Scholar] [CrossRef]
- Pollegioni, P.; Van der Linden, G.; Gras, M.; Olimpieri, I.; Anselmi, N.; Scarascia, G. Identification of resistant genotypes to anthracnose (Gnomonia leptostyla Fr. Ces) in Juglans spp. by functional and neutral markers. J. Biotechnol. 2010, 150, 114. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, Y.; Yang, K.; Li, J.; Sang, Y.; Huang, L.; Fan, S. Construction of a high-density genetic map using specific length amplified fragment markers and identification of a quantitative trait locus for anthracnose resistance in walnut (Juglans regia L.). BMC Genom. 2015, 16, 614. [Google Scholar] [CrossRef] [Green Version]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Hedin, P.A.; Langhans, V.E.; Graves, C.H. Identification of juglone in pecan as a possible factor of resistance to Fusicladium effusum. J. Agric. Food Chem. 1979, 27, 92–94. [Google Scholar] [CrossRef]
- Slatnar, A.; Mikulic Petkovsek, M.; Halbwirth, H.; Stampar, F.; Stich, K.; Veberic, R. Polyphenol metabolism of developing apple skin of a scab resistant and a susceptible apple cultivar. Trees 2012, 26, 109–119. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Slatnar, A.; Veberic, R.; Stampar, F.; Solar, A. Phenolic response in green walnut husk after the infection with bacteria Xanthomonas arboricola pv. juglandis. Physiol. Mol. Plant Pathol. 2011, 76, 159–165. [Google Scholar] [CrossRef]
- Solar, A.; Jakopic, J.; Veberic, R.; Stampar, F. Correlations between Xanthomonas arboricola pv. juglandis severity and endogenous juglone and phenolic acids in walnut. J. Plant Pathol. 2012, 94, 229–235. [Google Scholar] [CrossRef]
- Cline, S.; Neely, D. Relationship between juvenile-leaf resistance to anthracnose and the presence of juglone and hydrojuglone glucoside in black walnut. Phytopathology 1984, 74, 185–188. [Google Scholar] [CrossRef]
- Medic, A.; Jakopic, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and quantification of the major phenolic constituents in Juglans regia L. peeled kernels and pellicles, using HPLC–MS/MS. Food Chem. 2021, 352, 129404. [Google Scholar] [CrossRef] [PubMed]
- Medic, A.; Jakopic, J.; Solar, A.; Hudina, M.; Veberic, R. Walnut (J. regia) agro-residues as a rich source of phenolic compounds. Biology 2021, 10, 535. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.-H.; Du, X.-W.; Sun, G.-D.; Dong, W.-T.; Wang, W.-M. Identification and characterization of major constituents in Juglans mandshurica using ultra performance liquid chromatography coupled with time-of-flight mass spectrometry (UPLC-ESI-Q-TOF/MS). Chin. J. Nat. Med. 2018, 16, 525–545. [Google Scholar] [CrossRef]
- Wang, T.-M.; Fu, Y.; Yu, W.-J.; Chen, C.; Di, X.; Zhang, H.; Zhai, Y.-J.; Chu, Z.-Y.; Kang, T.-G.; Chen, H.-B. Identification of polar constituents in the decoction of Juglans mandshurica and in the medicated egg prepared with the decoction by HPLC-Q-TOF MS2. Molecules 2017, 22, 1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solar, A.; Colaric, M.; Hudina, M.; Stampar, F. Phenolic content of walnut fruit as affected by cultivar and developmental stage. Acta Hortic. 2005, 705, 231–240. [Google Scholar] [CrossRef]
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.M. Food flavonols: Nutraceuticals with complex health benefits and functionalities. Trends Food Sci. Technol. 2021. [Google Scholar] [CrossRef]
- Abian, O.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Vega, S.; Reyburn, H.T.; Rizzuti, B.; Velazquez-Campoy, A. Structural stability of SARS-CoV-2 3CLpro and identification of quercetin as an inhibitor by experimental screening. Int. J. Biol. Macromol. 2020, 164, 1693–1703. [Google Scholar] [CrossRef]
- Quiles, J.L.; Rivas-García, L.; Varela-López, A.; Llopis, J.; Battino, M.; Sánchez-González, C. Do nutrients and other bioactive molecules from foods have anything to say in the treatment against COVID-19? Environ. Res. 2020, 191, 110053. [Google Scholar] [CrossRef] [PubMed]
- Belgacem, I.; Li Destri Nicosia, M.G.; Pangallo, S.; Abdelfattah, A.; Benuzzi, M.; Agosteo, G.E.; Schena, L. Pomegranate peel extracts as safe natural treatments to control plant diseases and increase the shelf-life and safety of fresh fruits and vegetables. Plants 2021, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Belgacem, I.; Pangallo, S.; Abdelfattah, A.; Romeo, F.V.; Cacciola, S.O.; Li Destri Nicosia, M.G.; Ballistreri, G.; Schena, L. Transcriptomic analysis of orange fruit treated with pomegranate peel extract (PGE). Plants 2019, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangallo, S.; Destri Nicosia, M.G.; Agosteo, G.E.; Abdelfattah, A.; Romeo, F.V.; Cacciola, S.O.; Rapisarda, P.; Schena, L. Evaluation of a Pomegranate Peel Extract as an Alternative Means to Control Olive Anthracnose. Phytopathology 2017, 107, 1462–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolics | Rt (min) | [M − H]− (m/z) | MS2 (m/z) | Expressed as |
|---|---|---|---|---|
| neochlorogenic acid (3-caffeoylquinic acid) | 9.36 | 353 | 191, 179, 135 | neochlorogenic acid |
| procyanidin dimer 1 | 10.38 | 577 | 425, 407, 289 | procyanidin B1 |
| procyanidin dimer 2 | 11.47 | 577 | 425, 407, 289 | procyanidin B1 |
| 3-p-coumaoylquinic acid | 12.01 | 337 | 163, 191, 173 | 4-O-caffeoylquinic acid |
| (+) catechin | 12.22 | 289 | 245, 205, 179 | (+) catechin |
| dihydroxytetralone hexoside | 13.04 | 339 | 159, 177 | juglone |
| (−) epicatechin | 14.53 | 289 | 245, 205, 179 | (−) epicatechin |
| procyanidin dimer 3 | 15.53 | 577 | 425, 407, 289 | procyanidin B1 |
| hydrojuglone β-d-glucopyranoside | 16.26 | 337 | 175 | juglone |
| procyanidin dimer 4 | 16.89 | 577 | 425, 407, 289 | procyanidin B1 |
| hydrojuglone derivative pentoside 1 | 17.98 | 435 | 303, 285 | juglone |
| hydrojuglone derivative pentoside 2 | 18.26 | 435 | 303, 285 | juglone |
| dihydroxytetralone galloyl hexoside | 19.06 | 491 | 271, 331 | juglone |
| (epi)catechin derivative | 20.17 | 441 | 289 | (+) catechin |
| quercetin-3-galactoside | 20.53 | 463 | 301 | quercetin-3-galactoside |
| trihydroxytetralone galloyl hexoside | 20.76 | 507 | 331, 271 | juglone |
| hydrojuglone derivative pentoside 3 | 21.27 | 435 | 303, 285 | juglone |
| quercetin-3-xyloside | 21.56 | 433 | 301 | quercetin-3-xyloside |
| quercetin-3-arabinopyranoside | 21.73 | 433 | 301 | quercetin-3-arabinopyranoside |
| gallic acid derivative | 21.99 | 489 | 271, 313 | gallic acid |
| quercetin-3-arabinofuranoside | 22.21 | 433 | 301 | quercetin-3-arabinofuranoside |
| quercetin-3-rhamnoside | 22.43 | 447 | 301 | quercetin-3-rhamnoside |
| 1,4-naphthoquinone | 28.21 | 173 | 111, 155, 129, 145 | 1,4-naphthoquinone |
| hydrojuglone | 28.21 | 175 | 131, 147, 157, 115, 103 | juglone |
| juglone (5-hydroxy-1,4-naphthoquinone) | 29.99 | 189 | 161 | juglone |
| juglanin B | 31.37 | 327 | 312, 253 | juglone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medic, A.; Solar, A.; Hudina, M.; Veberic, R. Phenolic Response to Walnut Anthracnose (Ophiognomonia leptostyla) Infection in Different Parts of Juglans regia Husks, Using HPLC-MS/MS. Agriculture 2021, 11, 659. https://doi.org/10.3390/agriculture11070659
Medic A, Solar A, Hudina M, Veberic R. Phenolic Response to Walnut Anthracnose (Ophiognomonia leptostyla) Infection in Different Parts of Juglans regia Husks, Using HPLC-MS/MS. Agriculture. 2021; 11(7):659. https://doi.org/10.3390/agriculture11070659
Chicago/Turabian StyleMedic, Aljaz, Anita Solar, Metka Hudina, and Robert Veberic. 2021. "Phenolic Response to Walnut Anthracnose (Ophiognomonia leptostyla) Infection in Different Parts of Juglans regia Husks, Using HPLC-MS/MS" Agriculture 11, no. 7: 659. https://doi.org/10.3390/agriculture11070659
APA StyleMedic, A., Solar, A., Hudina, M., & Veberic, R. (2021). Phenolic Response to Walnut Anthracnose (Ophiognomonia leptostyla) Infection in Different Parts of Juglans regia Husks, Using HPLC-MS/MS. Agriculture, 11(7), 659. https://doi.org/10.3390/agriculture11070659