
Phenotypic Characterisation for Growth and Nut Characteristics Revealed the Extent of Genetic Diversity in Wild Macadamia Germplasm

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Materials
2.2. Data Collection
2.3. Data Analysis
3. Results
3.1. Variation in Growth and Nut Traits
3.2. Correlation among Growth and Nut Traits
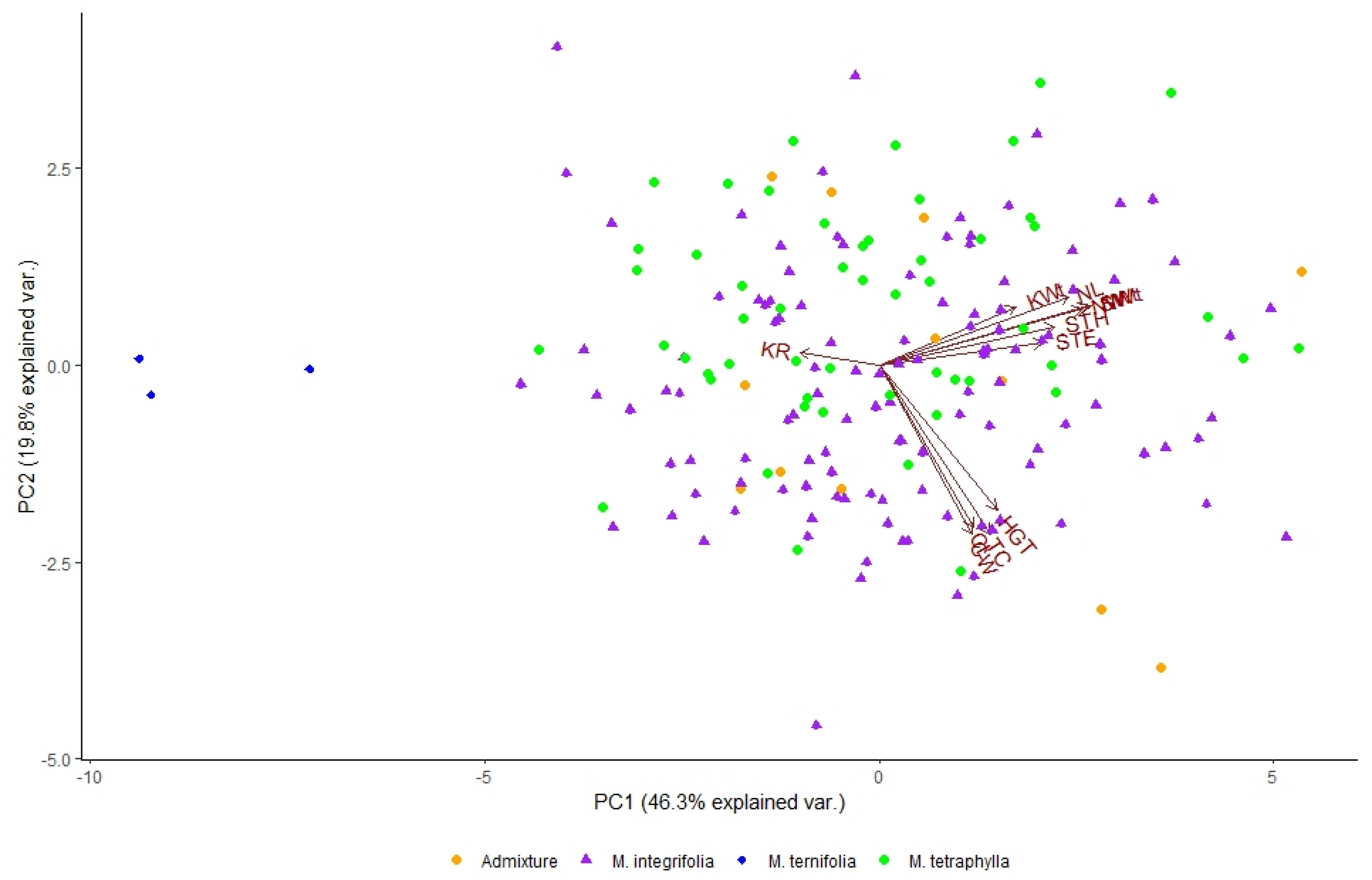
3.3. Principal Component Analysis
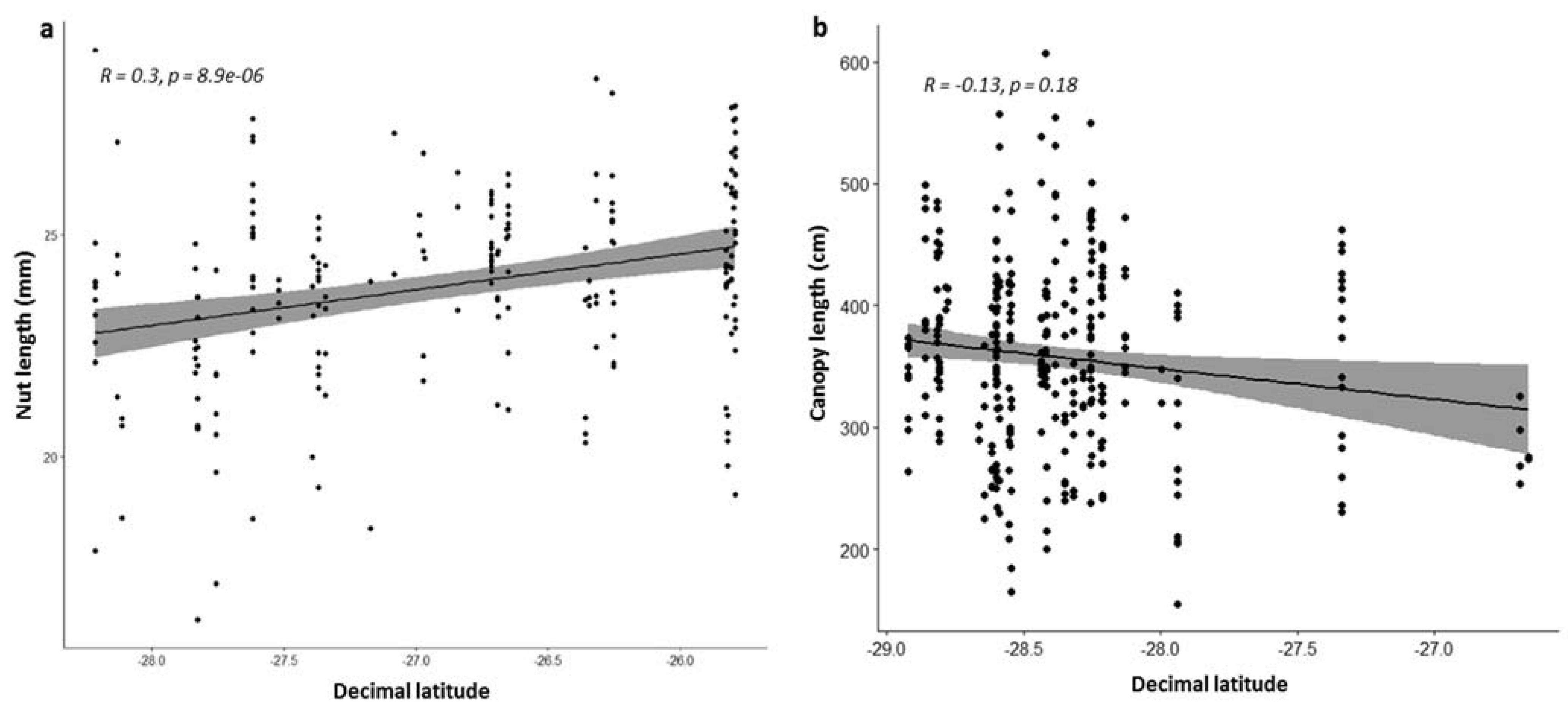
3.4. Effect of Geographic Distribution on Morphological Traits
4. Discussion
4.1. Variation in Growth and Nut Traits
4.2. Correlations among Phenotypic Traits
4.3. Relationship between Phenotypic Traits and Geographic Distribution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardner, C.M.; Peace, C.; Lowe, A.J.; Neal, J.; Pisanu, P.; Powell, M.; Schmidt, A.; Spain, C.; Williams, K. Genetic resources and domestication of macadamia. In Horticultural Reviews; Janick, J., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; Volume 35, pp. 1–125. [Google Scholar]
- Gross, C.L. Elaeagnaceae, Proteaceae 1. In Flora of Australia; Australian Biological Resources Study: Canberra, Australia, 1995; Volume 16, pp. 419–425. [Google Scholar]
- Gross, C.L.; Weston, P.H. Macadamia jansenii (Proteaceae), a new species from central Queensland. Aust. Syst. Bot. 1992, 5, 725–728. [Google Scholar] [CrossRef]
- Topp, B.L.; Nock, C.J.; Hardner, C.M.; Alam, M.; O’Connor, K.M. Macadamia (Macadamia spp.) Breeding. In Advances in Plant Breeding Strategies: Nut and Beverage Crops; Al-Khayri, J., Jain, S.D.J., Eds.; Springer: Cham, Switzerland, 2019; pp. 221–251. [Google Scholar]
- Dahler, J.; McConchie, C.; Turnbull, C. Quantification of Cyanogenic Glycosides in Seedlings of Three Macadamia (Proteaceae) Species. Aust. J. Bot. 1995, 43, 619–628. [Google Scholar] [CrossRef]
- Peace, C.P. Genetic Characterisation of Macadamia with DNA Markers. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 2004. [Google Scholar]
- Nock, C.J.; Hardner, C.M.; Montenegro, J.D.; Termizi, A.A.A.; Hayashi, S.; Playford, J.; Edwards, D.; Batley, J. Wild Origins of Macadamia Domestication Identified through Intraspecific Chloroplast Genome Sequencing. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Vahdati, K.; Karimi, R.; Ershadi, A. Genetic Structure of Some Wild Walnut Populations in Iran. Acta Hortic. 2015, 125–128. [Google Scholar] [CrossRef]
- Karimi, R.; Ershadi, A.; Vahdati, K.; Woeste, K. Molecular Characterization of Persian Walnut Populations in Iran with Microsatellite Markers. HortScience 2010, 45, 1403–1406. [Google Scholar] [CrossRef] [Green Version]
- Vahdati, K.; Pourtaklu, S.M.; Karimi, R.; Barzehkar, R.; Amiri, R.; Mozaffari, M.; Woeste, K. Genetic diversity and gene flow of some Persian walnut populations in southeast of Iran revealed by SSR markers. Plant Syst. Evol. 2015, 301, 691–699. [Google Scholar] [CrossRef]
- Nematollahi, A.; Golein, B.; Vahdati, K. Analysis of the Genetic Diversity in Citrus (Citrus spp.) Species Using SSR Markers. J. Plant Physiol. Breed. 2013, 3, 41–49. [Google Scholar]
- Bhandari, H.R.; Bhanu, A.N.; Srivastava, K.; Singh, M.N.; Shreya, A.H. Assessment of Genetic Diversity in Crop Plants—An Overview. Adv. Plants Agric. Res. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Karimi, R.; Ershadi, A.; Ehtesham Nia, A.; Sharifani, M.; Rasouli, M.; Ebrahimi, A.; Vahdati, K. Morphological and Molecular Evaluation of Persian Walnut Populations in Northern and Western Regions of Iran. Int. J. Nuts Relat. Sci. 2015, 5, 21–31. [Google Scholar]
- Mai, T.; Alam, M.; Hardner, C.; Henry, R.; Topp, B. Genetic Structure of Wild Germplasm of Macadamia: Species Assignment, Diversity and Phylogeographic Relationships. Plants 2020, 9, 714. [Google Scholar] [CrossRef]
- Alam, M.; Neal, J.; O’Connor, K.; Kilian, A.; Topp, B. Ultra-high-throughput DArTseq-based silicoDArT and SNP markers for genomic studies in macadamia. PLoS ONE 2018, 13, e0203465. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, K.; Kilian, A.; Hayes, B.; Hardner, C.; Nock, C.; Baten, A.; Alam, M.; Topp, B. Population structure, genetic diversity and linkage disequilibrium in a macadamia breeding population using SNP and silicoDArT markers. Tree Genet. Genomes 2019, 15, 1–16. [Google Scholar] [CrossRef]
- Hardner, C.; Winks, C.; Stephenson, R.; Gallagher, E. Genetic parameters for nut and kernel traits in macadamia. Euphytica 2001, 117, 151–161. [Google Scholar] [CrossRef]
- Leverington, R.E. Evaluation of macadamia nut varieties for processing. Qld. J. Agric. Sci. 1961, 19, 33–46. [Google Scholar]
- Gitonga, L.; Kahangi, E.; Muigai, A.; Ngamau, K.; Gichuki, S.; Cheluget, W.; Wepukhulu, S. Assessment of phenotypic diversity of macadamia (Macadamia spp) germplasm in Kenya using leaf and fruit morphology. Afr. J. Plant Sci. 2008, 2, 86–93. [Google Scholar]
- Wright, S. Evolution in mendelian populations. Bull. Math. Biol. 1990, 52, 241–295. [Google Scholar] [CrossRef] [Green Version]
- Gram, W.K.; Sork, V.L. Association between Environmental and Genetic Heterogeneity in Forest Tree Populations. Ecology 2001, 82, 2001. [Google Scholar] [CrossRef]
- De Woody, J.; Trewin, H.; Taylor, G. Genetic and morphological differentiation in Populus nigra L.: Isolation by colonization or isolation by adaptation? Mol. Ecol. 2015, 24, 2641–2655. [Google Scholar] [CrossRef] [Green Version]
- Arteaga, M.C.; Bello-Bedoy, R.; La Luz, J.L.L.-D.; Delgadillo, J.; Dominguez, R. Phenotypic variation of flowering and vegetative morphological traits along the distribution for the endemic species Yucca capensis (Agavaceae). Bot. Sci. 2015, 93, 765. [Google Scholar] [CrossRef]
- Król, K.; Gantner, M. Morphological Traits and Chemical Composition of Hazelnut from Different Geographical origins: A Review. Agriculture 2020, 10, 375. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, X.; Zhou, Q.; Deng, J. Morphological Variability between Geographical Provenances of Walnut Fruit (Juglans mandshurica) in the Eastern Liaoning Province, PR China. Pol. J. Environ. Stud. 2021. [Google Scholar] [CrossRef]
- Townsend, A.M. Geographic Variation in Fruit Characteristics of Acer rubrum. Bull. Torrey Bot. Club 1972, 99, 122. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Lai, L.; Jiang, L.; Zhuang, P.; Zhang, L.; Zheng, Y.; Baskin, J.M.; Baskin, C.C. Geographic variation in seed traits within and among forty-two species of Rhododendron (Ericaceae) on the Tibetan plateau: Relationships with altitude, habitat, plant height, and phylogeny. Ecol. Evol. 2014, 4, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Hardner, C.M.; Pisanu, P.C.; Boyton, S.J. National Macadamia Germplasm Conservation Program; Australian Macadamia Society: Lismore, Australia, 2004; pp. 8–62. [Google Scholar]
- Prichavudhi, K.; Yamamoto, A.H.Y. Effect of drying temperature on chemical composition and quality of macadamia nuts. Food Technol. 1965, 19, 1153–1156. [Google Scholar]
- Butler, D.G. Asreml: Fits the Linear Mixed Model R Package Version 4.1.0.110; VSN International Ltd.: Hemel Hempstead, UK, 2019. [Google Scholar]
- Brian, G.P.; Carl, A.P. PerformanceAnalytics: Econometric Tools for Performance and Risk Analysis. R Package Version 2.0.4. 2020. Available online: https://rdrr.io/cran/PerformanceAnalytics/ (accessed on 30 May 2021).
- Vu, V.Q. ggbiplot: A ggplot2 Based Biplot. R Package Version 0.55. 2011. Available online: https://rdrr.io/github/vqv/ggbiplot/ (accessed on 30 May 2021).
- Kassambara, A. ggpubr: “ggplot” Based Publication Ready Plots. R Package Version 0.2.5. 2020. Available online: https://rdrr.io/cran/ggpubr/ (accessed on 30 May 2021).
- Department of Agriculture and Fisheries. Macadamia Industry Benchmark Report 2009 to 2016 Seasons; Department of Agriculture and Fisheries: Wilsonton, Australia, 2017. [Google Scholar]
- Huett, D.O. Macadamia physiology review: A canopy light response study and literature review. Aust. J. Agric. Res. 2004, 55, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Hardner, C.M.; Winks, C.W.; Stephenson, R.A.; Gallagher, E.G.; McConchie, C.A. Genetic parameters for yield in macadamia. Euphytica 2002, 125, 255–264. [Google Scholar] [CrossRef]
- Toft, B.D.; Alam, M.; Topp, B. Estimating genetic parameters of architectural and reproductive traits in young macadamia cultivars. Tree Genet. Genomes 2018, 14, 50. [Google Scholar] [CrossRef]
- Zeng, H.; Zou, M.; Zhang, H.; Luo, L. Characters analysis of the Macadamia cv. Ikaika (333) germplasm resource. Afr. J. Biotechnol. 2011, 10, 17443–17450. [Google Scholar] [CrossRef]
- Storey, W.B. Macadamia Selection on the Basis of Nut Quality. Calif. Macad. Soc. 1960, 9, 69–74. [Google Scholar]
- O’Connor, K. Selection Strategies to Improve Yield in Macadamia Using Component Traits and Genomics. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2019. [Google Scholar]
- Simpson, E.H. The Interpretation of Interaction in Contingency Tables. J. R. Stat. Soc. Ser. B Methodol. 1951, 13, 238–241. [Google Scholar] [CrossRef]
- Thompson, T.; Baker, J. Heritability and Phenotypic Correlations of Six Pecan Nut Characteristics. J. Am. Soc. Hortic. Sci. 1993, 118, 415–418. [Google Scholar] [CrossRef]
- Sorkheh, K.; Shiran, B.; Khodambashi, M.; Moradi, H.; Gradziel, T.; Martinez-Gomez, P. Correlations between quantitative tree and fruit almond traits and their implications for breeding. Sci. Hortic. 2010, 125, 323–331. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Sarooghi, F.; Abbasi, F. Phenotypic variation of Prunus scoparia germplasm: Implications for breeding. Sci. Hortic. 2016, 207, 193–202. [Google Scholar] [CrossRef]
- Sorkheh, K.; Shiran, B.; Rouhi, V.; Asadi, E.; Jahanbazi, H.; Moradi, H.; Gradziel, T.; Martinezgomez, P. Phenotypic diversity within native Iranian almond (Prunus spp.) species and their breeding potential. Genet. Resour. Crop. Evol. 2009, 56, 947–961. [Google Scholar] [CrossRef]
- Bak, T.; Karadeniz, T. Effects of Branch Number on Quality Traits and Yield Properties of European Hazelnut (Corylus avellana L.). Agriculture 2021, 11, 437. [Google Scholar] [CrossRef]
- Mast, A.R.; Willis, C.L.; Jones, E.H.; Downs, K.M.; Weston, P.H. A smaller Macadamia from a more vagile tribe: Inference of phylogenetic relationships, divergence times, and diaspore evolution in Macadamia and relatives (tribe Macadamieae; Proteaceae). Am. J. Bot. 2008, 95, 843–870. [Google Scholar] [CrossRef]
- Pan, Y.; Li, S.; Wang, C.; Ma, W.; Xu, G.; Shao, L.; Li, K.; Zhao, X.; Jiang, T. Early evaluation of growth traits of Larix kaempferi clones. J. For. Res. 2018, 29, 1031–1039. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characters | Admixture | M. integrifolia | M. tetraphylla | M. ternifolia | M. jansenii | |
|---|---|---|---|---|---|---|
| Growth traits: | N | 13 | 119 | 107 | 5 | N/A |
| HGT (cm) | Mean | 481.5 a | 476.7 a | 446.4 b | 329.2 c | N/A |
| SE | 19.6 | 7.1 | 7.6 | 32.1 | N/A | |
| Min | 353.2 | 302.5 | 349.6 | 287.1 | N/A | |
| Max | 613.5 | 629.3 | 597.3 | 366.2 | N/A | |
| TC (cm) | Mean | 54.0 a | 52.8 a | 46.3 b | 27.5 c | N/A |
| SE | 2.8 | 1.0 | 1.1 | 4.6 | N/A | |
| Min | 35.4 | 31.9 | 26.9 | 23.1 | N/A | |
| Max | 78.4 | 71.8 | 66.3 | 30.0 | N/A | |
| CL (cm) | Mean | 456.2 a | 383.1 b | 355.5 c | 245.3 d | N/A |
| SE | 20.5 | 7.5 | 8.0 | 33.7 | N/A | |
| Min | 332.9 | 221.7 | 256.1 | 189.7 | N/A | |
| Max | 622.6 | 503.3 | 527.9 | 295.8 | N/A | |
| CW (cm) | Mean | 407.1 a | 387 a | 352.6 b | 244.5 c | N/A |
| SE | 18.1 | 6.6 | 7.1 | 29.8 | N/A | |
| Min | 317.1 | 252.6 | 253.7 | 173.3 | N/A | |
| Max | 515.4 | 522.5 | 471.8 | 306.8 | N/A | |
| Nut traits: | N | 12 | 117 | 53 | 3 | 3 |
| NWt (g) | Mean | 7.08 a | 6.68 a | 6.33 a | 1.32 b | 1.99 b |
| SE | 0.50 | 0.19 | 0.25 | 0.97 | 0.89 | |
| Min | 5.89 | 4.3 | 4.58 | 1.17 | 1.81 | |
| Max | 9.96 | 9.15 | 9.22 | 1.49 | 2.22 | |
| NL (mm) | Mean | 25.7 a | 23.78 b | 25.7 a | 14.67 d | 18.61 c |
| SE | 0.66 | 0.26 | 1.16 | 1.29 | 0.34 | |
| Min | 23.75 | 20.91 | 22.73 | 13.56 | 17.35 | |
| Max | 29.44 | 25.98 | 29.94 | 15.23 | 19.58 | |
| NW (mm) | Mean | 24.65 a | 23.3 b | 23.59 ab | 13.69 c | 15.46 c |
| SE | 0.64 | 0.26 | 0.32 | 1.24 | 1.09 | |
| Min | 23.42 | 20.59 | 21.63 | 12.70 | 15.21 | |
| Max | 26.83 | 25.26 | 26.48 | 14.26 | 15.71 | |
| STH (mm) | Mean | 5.49 a | 5.89 a | 6.04 a | 2.31 b | 3.72 b |
| SE | 0.28 | 0.11 | 0.14 | 0.54 | 0.49 | |
| Min | 4.85 | 4.59 | 4.71 | 2.16 | 3.18 | |
| Max | 6.78 | 8.02 | 8.12 | 2.41 | 4.12 | |
| STE (mm) | Mean | 2.54 b | 2.96 a | 2.68 b | 1.51 c | 1.68 c |
| SE | 0.17 | 0.07 | 0.09 | 0.33 | 0.31 | |
| Min | 1.76 | 2.10 | 2.14 | 1.24 | 1.52 | |
| Max | 3.10 | 4.49 | 3.75 | 1.78 | 1.87 | |
| SWt (g) | Mean | 4.81 a | 4.75 a | 4.56 a | 0.82 b | 1.22 b |
| SE | 0.40 | 0.16 | 0.20 | 0.78 | 0.72 | |
| Min | 3.70 | 3.10 | 2.94 | 0.67 | 1.10 | |
| Max | 6.45 | 7.13 | 7.20 | 0.96 | 1.43 | |
| KWt (g) | Mean | 2.02 a | 1.55 b | 1.62 b | 0.52 c | 0.58 c |
| SE | 0.13 | 0.05 | 0.06 | 0.25 | 0.23 | |
| Min | 1.60 | 0.92 | 1.03 | 0.48 | 0.53 | |
| Max | 2.85 | 2.36 | 2.53 | 0.57 | 0.66 | |
| KR (%) | Mean | 29.0 a | 24.3 b | 25.2 b | 34.7 a | 30.9 a |
| SE | 1.4 | 0.5 | 0.7 | 2.7 | 2.5 | |
| Min | 24.0 | 18.5 | 16.6 | 32.5 | 29.8 | |
| Max | 32.7 | 32.4 | 33.1 | 37.8 | 33.0 | |
| Variable | VG ± SE * | VE ± SE | H |
|---|---|---|---|
| HGT | 3811.9 ± 444.0 | 2528.3 ± 156.3 | 0.60 |
| TC | 72.6 ± 9.0 | 60.9 ± 3.8 | 0.54 |
| CL | 3973.1 ± 492.1 | 3187.6 ± 198.4 | 0.55 |
| CW | 2929.5 ± 380.1 | 2851.7 ± 177.0 | 0.51 |
| NWt | 1.7 ± 0.3 | 1.2 ± 0.2 | 0.59 |
| NL | 2.7 ± 0.5 | 2.7 ± 0.3 | 0.50 |
| NW | 2.2 ± 0.5 | 2.8 ± 0.3 | 0.43 |
| STH | 0.5 ± 0.1 | 0.4 ± 0.0 | 0.58 |
| STE | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.64 |
| SWt | 1.2 ± 0.2 | 0.7 ± 0.1 | 0.62 |
| KWt | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.61 |
| KR | 13.6 ± 2.3 | 9.4 ± 1.2 | 0.59 |
| HGT | TC | CL | CW | NWt | NL | NW | STH | STE | SWt | KWt | KR | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TC | 0.76 | |||||||||||
| CL | 0.49 | 0.65 | ||||||||||
| CW | 0.57 | 0.65 | 0.71 | |||||||||
| NWt | 0.38 | 0.36 | 0.30 | 0.29 | ||||||||
| NL | 0.23 | 0.19 | 0.18 | 0.14 ns | 0.73 | |||||||
| NW | 0.35 | 0.36 | 0.31 | 0.27 | 0.88 | 0.87 | ||||||
| STH | 0.32 | 0.27 | 0.15 | 0.22 | 0.68 | 0.73 | 0.68 | |||||
| STE | 0.35 | 0.32 | 0.16 | 0.26 | 0.67 | 0.37 | 0.49 | 0.68 | ||||
| SWt | 0.35 | 0.32 | 0.24 | 0.28 | 0.95 | 0.69 | 0.81 | 0.72 | 0.75 | |||
| KWt | 0.22 | 0.20 | 0.24 | 0.15 | 0.74 | 0.65 | 0.76 | 0.29 | 0.11 ns | 0.60 | ||
| KR | −0.21 | −0.21 | −0.07 ns | −0.21 | −0.28 | −0.19 | −0.22 | −0.57 | −0.7 | −0.43 | 0.32 |
| Traits | Variance Component | |||||
|---|---|---|---|---|---|---|
| Site | Site (Accession) | Site: Year | ||||
| Estimate ± SE | % Variation | Estimate ± SE | % Variation | Estimate ± SE | % Variation | |
| M. integrifolia | ||||||
| HGT | 437.2 ± 404.7 | 5.9 | 2887.2 ± 620.4 | 38.9 | 0.0 ± NA | 0.0 |
| TC | 14.0 ± 9.6 | 9.8 | 47.2 ± 11.6 | 32.9 | 0.0 ± NA | 0.0 |
| CL | 58.1 ± 339.3 | 0.8 | 3059.0 ± 672.6 | 42.0 | 0.0 ± NA | 0.0 |
| CW | 61.2 ± 324.9 | 0.9 | 2900.2 ± 633.7 | 43.4 | 0.9 ± NA | 0.0 |
| NWt | 0.85 ± 0.45 | 27.1 | 0.98 ± 0.27 | 31.5 | 0.36 ± 0.27 | 11.5 |
| NL | 0.57 ± 0.57 | 12.1 | 1.48 ± 0.45 | 31.6 | 0.53 ± 0.48 | 11.3 |
| NW | 0.83 ± 0.66 | 16.4 | 1.11 ± 0.45 | 21.9 | 0.64 ± 0.54 | 12.6 |
| STH | 0.18 ± 0.09 | 19.4 | 0.33 ± 0.1 | 35.6 | 0.00 ± NA | 0.0 |
| STE | 0.07 ± 0.04 | 17.3 | 0.19 ± 0.04 | 49.2 | 0.00 ± 0.02 | 0.5 |
| SWt | 0.55 ± 0.29 | 26.2 | 0.76 ± 0.19 | 36.5 | 0.21 ± 0.17 | 9.9 |
| KWt | 0.04 ± 0.02 | 23.8 | 0.06 ± 0.02 | 39.3 | 0.01 ± 0.01 | 3.2 |
| KR | 0.20 ± 1.83 | 1.0 | 13.54 ± 2.68 | 68.5 | 1.48 ± 1.40 | 7.5 |
| M. tetraphylla | ||||||
| HGT | 929.7 ± 567.5 | 14.5 | 2979.5 ± 630.0 | 46.5 | 0 ± NA | 0.0 |
| TC | 6.7 ± 9.0 | 5.7 | 64.7 ± 13.6 | 54.5 | 0 ± NA | 0.0 |
| CL | 137.2 ± 489.8 | 2.0 | 4111.7 ± 842.6 | 58.5 | 0 ± NA | 0.0 |
| CW | 193.2 ± 330.0 | 3.7 | 2481.4 ± 554.4 | 47.0 | 0 ± NA | 0.0 |
| NWt | 0.33 ± 0.45 | 10.1 | 0.77 ± 0.61 | 23.7 | 0 ± NA | 0.0 |
| NL | 0.96 ± 1.1 | 12.8 | 1.82 ± 1.45 | 24.3 | 0 ± NA | 0.0 |
| NW | 0 ± NA | 0.0 | 0.76 ± 1.09 | 12.9 | 0 ± NA | 0.0 |
| STH | 0.13 ± 0.19 | 13.0 | 0.48 ± 0.22 | 47.0 | 0 ± NA | 0.0 |
| STE | 0 ± NA | 0.0 | 0.14 ± 0.05 | 45.4 | 0 ± NA | 0.0 |
| SWt | 0 ± NA | 0.0 | 0.89 ± 0.38 | 45.1 | 0 ± NA | 0.0 |
| KWt | 0.04 ± 0.05 | 12.3 | 0.08 ± 0.06 | 28.0 | 0.02 ± 0.05 | 5.9 |
| KR | 4.11 ± 8.87 | 12.0 | 2.21 ± 4.29 | 6.5 | 9.58 ± 9.7 | 28.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, T.T.P.; Hardner, C.M.; Alam, M.M.; Henry, R.J.; Topp, B.L. Phenotypic Characterisation for Growth and Nut Characteristics Revealed the Extent of Genetic Diversity in Wild Macadamia Germplasm. Agriculture 2021, 11, 680. https://doi.org/10.3390/agriculture11070680
Mai TTP, Hardner CM, Alam MM, Henry RJ, Topp BL. Phenotypic Characterisation for Growth and Nut Characteristics Revealed the Extent of Genetic Diversity in Wild Macadamia Germplasm. Agriculture. 2021; 11(7):680. https://doi.org/10.3390/agriculture11070680
Chicago/Turabian StyleMai, Thuy T. P., Craig M. Hardner, Mobashwer M. Alam, Robert J. Henry, and Bruce L. Topp. 2021. "Phenotypic Characterisation for Growth and Nut Characteristics Revealed the Extent of Genetic Diversity in Wild Macadamia Germplasm" Agriculture 11, no. 7: 680. https://doi.org/10.3390/agriculture11070680
APA StyleMai, T. T. P., Hardner, C. M., Alam, M. M., Henry, R. J., & Topp, B. L. (2021). Phenotypic Characterisation for Growth and Nut Characteristics Revealed the Extent of Genetic Diversity in Wild Macadamia Germplasm. Agriculture, 11(7), 680. https://doi.org/10.3390/agriculture11070680