1. Introduction
Today, we face global problems. such as vitamin and micronutrient deficiency, which are related with existing obesity and chronic diseases [
1]. Thus, the cultivation of Swiss chard should be encouraged because it is an excellent source of mineral nutrients and different phytochemicals and, at the same time, is a cheap food crop [
2]. Swiss chard (
Beta vulgaris ssp.
cicla L.), green leafy vegetables of the
Chenopodiaceae family are rich in bioactive compounds, such as phytopigments (chlorophylls and carotenoids), phenolic acids (syringic, caffeic, and p-coumaric), flavonoids (kaempferol, quercetin, and glycosides derived from apigenin) vitamins C, A, and B, and minerals (iron, potassium, calcium, magnesium, and manganese) with antioxidant and immunomodulating properties. They also contain dietary fibers and proteins [
3,
4].
The use of agronomic strategies to increase the content of mineral elements in the edible parts of crops is often known as agronomic biofortification. Zn biofortification is a promising approach to accumulating high concentrations of Zn in various edible plants to address human health problems associated with insufficient consumption of this microelement [
5]. Zinc (Zn) is an essential plant nutrient belonging to the micronutrients that face the problem of poor bioavailability, though this element plays a critical role in all vital functions of plants [
6]. In plants, Zn deficiency can cause severe crop production problems, as Zn is required for the detoxification of toxic oxygen free radicals, photosynthesis, protein synthesis, growth regulation, pollination, and disease defense mechanisms [
7].
Currently, nanoparticles (NPs) have received rapidly growing attention and find a wide range of applications in different economic sectors, such as food technology and modern agricultural production systems [
8,
9]. The application of NPs in agriculture is an effective strategy to improve agricultural practices aiming to minimize nutrient losses in fertilization, reduce the plant protection products for sustainable crop cultivation, and increase food security to meet the nutrition needs of the rapidly growing global population [
10,
11]. The success of using NPs for plant growth depends on many factors, such as plant species, their growth stage, and growth medium. In addition, the size, concentration, surface structure, physicochemical properties, and stability of NPs have a significant influence [
12]. But to date, there is still lacking knowledge about the uptake capacity, permissible limit, and ecotoxicity of different NPs in plants. Understanding the interactions between NPs and plants is important for comprehending nanotechnology’s impact on agriculture, with a focus on plant toxicity concerns and risks to human health.
Zinc oxide (ZnO) NPs are inorganic metal oxide compounds with nanoparticle sizes and large surface areas, and are commonly used in several industrial products [
13,
14,
15,
16,
17], as well as in agricultural and environmental sectors [
8,
9]. ZnO NPs have been effectively used in plant cultivation, due to their multifunctionality, which includes their importance as an essential element for nutrition, participation in metabolic functions, and abilities for simultaneous use as fertilizers and pesticides. [
9,
10]. Agricultural usage of ZnO NPs has shown positive and negative effects on plant growth [
8], depending on NP size, concentration, exposure time, and plant species [
18,
19,
20]. For instance, improved photosynthesis, induced generation of antistress enzymes, and affected nutritional quality were observed in cilantro with ZnO NPs exposures at concentrations of 0, 100, 200, and 400 mg kg
−1 [
21]. The treatment of radish sprouts with ZnO NPs (60 ppm) had stimulative effects on growth, quantity, and quality [
22]. The foliar application of 500–1000 ppm of ZnO NPs expressed the increased leaf length, width, surface area, and color, as well as higher protein and dietary fiber content values in spinach compared to control plant leaves [
18]. Recently, Sharifan with co-authors [
23] published the data regarding ZnO NPs’ effects on spinach (
Spinacia oleracea L.), parsley (
Petroselinum sativum L.), and cilantro (
Coriandrum sativum L.). They noted that the impacts of ZnO NPs at the 100 mg L
−1 level on the growth and yield of all three species were positive concerning increases in the nutritional values and reductions in the heavy metals in roots. However, in spring barley, all the tested concentrations (300 and 2000 mg L
−1) of ZnO NPs showed toxic effects. Suspensions affected cellular and sub-cellular organelles’ photosynthetic efficiency, anatomy, and ultrastructure [
24]. The harmful impact on the growth and development of the
Arabidopsis thaliana plants was also revealed when exposed to lower concentrations of ZnO NPs [
25,
26]. In another study, 400 mg L
−1 of ZnO NPs resulted in phytotoxic effects on Pisum sativum L., which induced oxidative stress [
27]. Thus, a review of the literature reveals inconsistent conclusions about the effects of ZnO NPs on plants and requires more profound research of these NPs on various aspects of plant growth and physiology. Even though the interaction between plants and ZnO NPs has been the focus of various publications, the NPs’ size-specific effects have not been well elucidated. Also, the knowledge about the impacts of Zn NPs on leafy greens is still limited. The targeted increase of NPs in leafy greens requires appropriate risk assessment and regulation. The use of NPs raises concern regarding the bioaccumulation of potentially toxic levels of heavy metals from metallic nanomaterials in edible plant tissues.
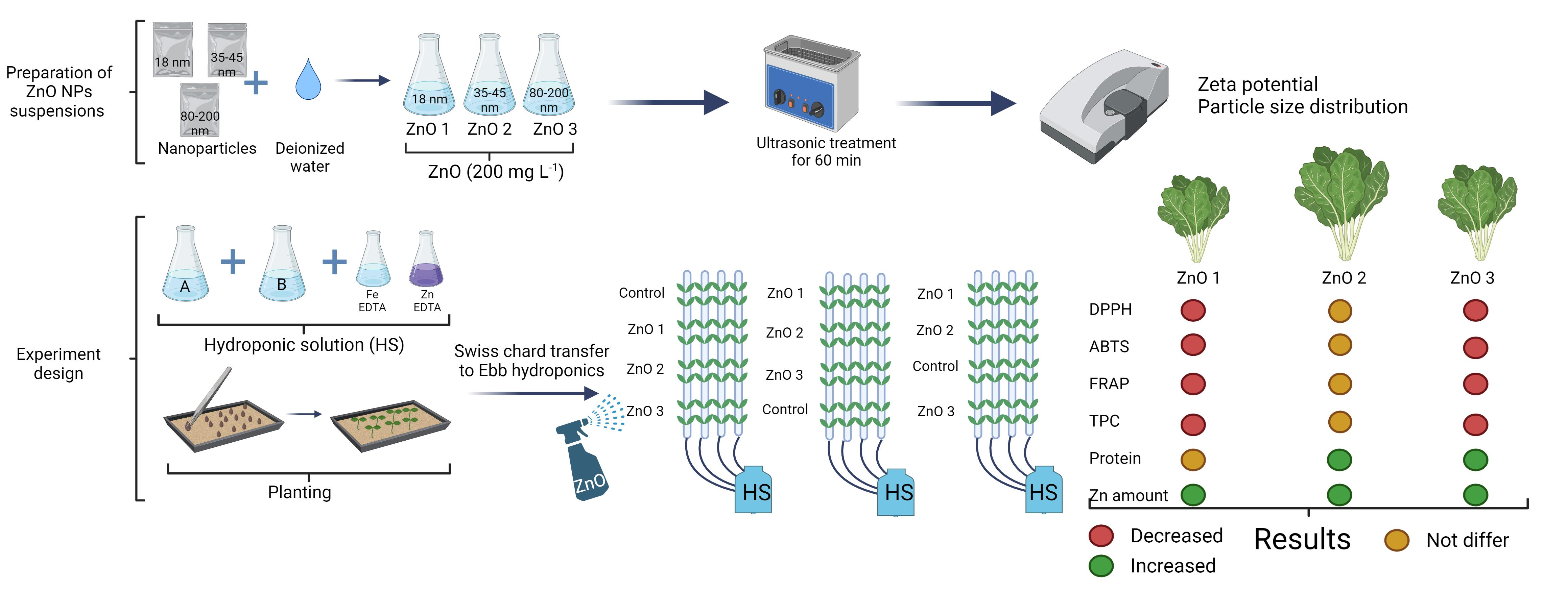
To understand the interactions between ZnO nanoparticles and Swiss chard plant physiology, this investigation was aimed: (i) to compare plant growth responses of Swiss chard plants exposed via foliar spray to a varying particle size of ZnO-NPs (ii) to analyze the physiological changes and nutritional quality of Swiss chard leaves treated with ZnO nanoparticles and (iii) to assess the accumulation of mineral nutrients on human health.
2. Materials and Methods
2.1. Growth Conditions
Experiments were performed in a walk-in controlled environment plant growth chamber (4 × 6 m, h = 3.2 m) at the Institute of Horticulture, in the Lithuanian Research Centre for Agriculture and Forestry. Seeds of the Swiss chard type ‘Barese’ (Beta vulgaris L.; CN Seeds, Ely, UK) were sown in Rockwool cubes (2.5 cm × 2.5 cm× 3.0 cm) and presoaked in deionized water with an adjusted pH of 5.0 using diluted sulfuric acid, and placed in a plastic tray. Two hundred seedlings were germinated in a growth chamber with a day/night temperature range of 21 ± 2/17 ± 2 °C, 12 h photoperiod, light intensity—220 µmol m−2 s−1, and the relative humidity was controlled to 60 ± 5%. After five days, a hydroponic solution was supplied in the tray with the following average concentrations of nutrients (mg L−1): N (120), Ca (88), P (20), K (128), Mg (40), S (53), B (0.16), Mo (0.2), Mn (0.08), Cu (0.08), Fe (1.6), Zn (0.8). The pH and the electrical conductivity (EC) of the nutrient solution were measured daily using a portable meter (GroLine HI9814, Hanna Instruments, Woonsocket, RI, USA), and were adjusted to a pH of 6.0 and an EC of 1.2 mS cm−1 using sulfuric acid or sodium bicarbonate. The 14-day-old Swiss chard seedlings of equal size were placed in the Ebb hydroponic systems, with 80 L containers containing a hydroponic solution composed of deionized water and macro- and microelements (composition described above) which was recirculated and oxygenated with an air stone. The pH (6.0 ± 0.2) and EC (1.4 ± 0.1 mS cm−1) of the nutrient solution tanks were measured as described previously, and adjusted using sulfuric acid or sodium bicarbonate daily.
The microclimate in the growth chamber was autonomously and independently controlled using the Phytotron Microclimate Control System developed in IH, based on separate microcontrollers (AL-2-24MR-D, Mitsubishi Electric, Tokyo, Japan). The air temperature was measured with resistance temperature detectors (P-100; OMEGA Engineering Ltd., Norwalk, CT, USA), and data for these measurements were transmitted to the microcontrollers. The relative humidity and CO2 concentration were measured by capacitive sensors (CO2RT(-D); Regin, Kållered, Sweden) and controlled by additional humidifiers. Data were collected every minute, processed, and stored on the operator panel (E1000, Mitsubishi Electric, Tokyo, Japan).
Three-week-old plants were exposed via foliar spray to a varying particle size of ZnO-NPs (18, 35–45, and 80–200 nm) at a concentration of 200 ppm, and control plants were sprayed with distilled water. The plants were sprayed until thoroughly wetted with aqueous suspensions. During the foliar application of ZnO-NPs, the surfaces of the Ebb hydroponic systems were covered with a polyethylene sheath to avoid contamination. Once exposed to ZnO-NP suspensions, Swiss chard leaves were allowed to grow for seven days.
The ZnO-NPs used for plant exposure in this study were from a commercial product (US Research Nanomaterials, Inc., Houston, TX, USA). Three suspensions with different sizes of ZnO NPs (particle size: 18, 35–45, 80–200 nm; purity: 99%; US Research Nanomaterials, Inc., Houston, TX, USA) were prepared with deionized water. The NPs, with a concentration of 200 ppm, were suspended using an ultrasonic disperser for 60 min. The NPs size and suspension stability were measured using a Delsa™ Nano Submicron Particle Size (Beckman Coulter Instruments Corporation, Fullerton, CA, USA) and a Zeta Potential device (Dispersion Technology Inc., Bedford Hills, New York, NY, USA).
Table 1 shows the positive particle surface charge of the 35–45 nm ZnO NP suspension and the neutral particle surface charge of the 18 and 80–200 nm ZnO NP suspensions. The stable system of ZnO NPs were found in the 35–45 nm ZnO suspension, according to zeta potential. Besides, the polydispersity index (PDI) showed that NP suspensions were monodisperse.
2.2. Growth Characteristics and Non-Destructive Measurements
At the end of the experiment, the shoot weight (fresh and dry) and leaf area were measured for ten representative plants per treatment (n = 10). The fresh biomass was measured with an electronic scale (Mettler Toledo, ML104T/00; Mettler-Toledo, Columbus, OH, USA), and subsequently, the shoots were dried at +70 °C in an oven (Venti cell 222, Medcenter Einrichtungen, Gräfeling, Germany) for 48 h before measuring dry weight. The leaf area was measured using a leaf area meter (CI-202 Laser Area Meter; CID BioScience, Camas, WA, USA).
Non-destructive measurements of leaf chlorophyll and flavonol indexes in the fully developed leaves (10 plants per treatment, n = 10) were performed using a chlorophyll and flavonoid meter (Force-A DualexR 4 Scientific, Ocala, FL, USA).
2.3. Antioxidant Properties and Total Phenolic Compounds
The fresh Swiss chard leaves were immediately frozen in liquid N2 and lyophilized for antioxidant activity and phenolic compound analysis. Extracts were prepared by mixing the 100 mg of lyophilized plant material with 5 mL of 80% methanol (Sigma-Aldrich, St. Louis, MO, USA) and transferred to a 15 mL polypropylene conical centrifuge tube (Labbox Labware S.L., Barcelona, Spain). After 24 h, the samples were centrifuged for 10 min at 3000 rpm (Hermle Z300K, Baden-Württemberg, Germany), extracts were filtered through cellulose filters, and the supernatant was used for further analyses. Each of the three biological replicates consisted of at least eight conjugated plants.
The antioxidant properties of Swiss chard leaves were evaluated as the DPPH (2-diphenyl-1-picrylhydrazyl) assay, the ABTS (2,20-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid)) the diammonium salt radical scavenging activities, and the Fe2+ reducing antioxidant power assay (FRAP); also, the total contents of phenolic compounds were determined. in three analytical replications.
For the DPPH assay, the 126.8 μM DPPH (100% purity; Sigma-Aldrich, Burlington, MA, USA) solution was prepared in methanol [
28]. Subsequently, 290 μL of the DPPH solution was transferred to a test tube and mixed with 20 μL of the Swiss chard leaves’ extract. The absorbance was scanned at 515 nm (SPECTROstar Nano, BMG Labtech microplate reader, Ortenberg, Germany) while reacting for 16 min. The free radical scavenging capacity was expressed as μmol of DPPH radicals scavenged per 1 g of leaf fresh weight (mmol g
−1 DW). A calibration curve was determined using Trolox (6-hydroxy-2,5,7,8-tetramethychroman-2-carboxylic acid; 97% purity; Sigma-Aldrich, Burlington, MA, USA) as an external standard, with a range of concentrations from 0.1 to 0.6 mM (R
2 = 0.99).
The ABTS radical cation was obtained by incubating a 7 mM ABTS stock solution with 2.45 mM potassium persulfate (K
2S
2O
8; 99% purity; Sigma-Aldrich, Burlington, MA, USA) and allowing the mixture to stand in the dark at room temperature for 12–16 h before use [
29]. Thereafter, 20 μL of the prepared sample was mixed with 290 μL of ABTS solution (ABTS stock solution was diluted 1:7), and the absorbance was measured after 11 min (plateau phase) at 734 nm (SPECTROstar Nano, BMG Labtech microplate reader, Ortenberg, Germany). The ABTS scavenging activity of Swiss chard leaves extracts was calculated as the difference between the initial absorbance and the absorbance after reacting for 10 min. A calibration curve was determined using Trolox (6-hydroxy-2,5,7,8-tetramethychroman-2-carboxylic acid; 97% purity; Sigma-Aldrich, Burlington, MA, USA) as an external standard, with a range of concentrations from 0.1 to 0.8 mM (R
2 = 0.99). It was expressed as ABTS µmol scavenged per 1 g of leaf fresh weight (mmol g
−1 DW).
The FRAP method is based on reducing ferric ions (Fe
3+) to ferrous ions (Fe
2+). The fresh reaction mixture was prepared by mixing a 300 mM, pH 3.6 acetate buffer, 10 mM TPTZ (2,4,6-tripyridyl-s-triazine) solution in 40 mM of HCl and 20 mM of FeCl
3 × 6H
2O at 10:1:1 (
v/
v/
v) [
30]. Subsequently, 20 µL of the sample was mixed with 290 μL of the reaction mixture and incubated in the dark for 30 min. Readings of the colored product (ferrous tripyridyl-triazine complex) were then taken at 593 nm with a SPECTROstar Nano BMG Labtech microplate reader (Ortenberg, Germany). A calibration curve was determined using Fe
2(SO
4)
3 (iron (III) sulfate; 97% purity; Sigma-Aldrich, Burlington, MA, USA) as an external standard, with a range of concentrations from 0.005 to 0.5 mM (R
2 = 0.99). The antioxidant power was expressed as Fe
2+ antioxidant capacity (Fe
2+ µmol g
−1 DW).
The total content of phenolic compounds was determined as gallic acid equivalents. A 20 µL aliquot of the sample extract was mixed with 20 µL of 10% (
w/v) Folin–Ciocalteu reagent and 160 µL of a 1 M Na
2CO
3 solution [
31]. After incubation for 20 min in the dark, the absorbance was measured at 765 nm (SPECTROstar Nano, BMG Labtech microplate reader, Ortenberg, Germany). The total phenolic compounds’ quantity in mg g
−1 of leaf dry weight was calculated from the calibration curve of the gallic acid (0.01–0.1 mg mL
−1, R
2 = 0.99).
2.4. Measurement of Soluble Protein Content
The extracts used to determine the soluble protein content in the Swiss chard leaves were prepared by grinding 0.5 g of fresh sample with liquid nitrogen and diluting within 5 mL of a 100 mM potassium-phosphate buffer (pH 7.8, 0.1 mM EDTA). After centrifugation for 10 min at 3000 rpm (Hermle Z300K, Baden-Württemberg, Germany), the supernatant was collected and used for total soluble protein measurement. All steps in the preparation of the extract were carried out at 4 °C. The dye-binding method and bovine serum albumin were used for soluble protein determination as the standard. A volume of 20 µL of enzyme extract was mixed with 290 µL of Bradford reagent diluted to 1:5 with DI water. Absorbance was read after 2 min using a spectrophotometer (SPECTROstar Nano, BMG Labtech microplate reader, Ortenberg, Germany) at 595 nm [
32].
2.5. Determination of Mineral Nutrients
The contents of macro- and microelements in the Swiss chard leaves were determined using the microwave-assisted digestion technique, combined with inductively coupled plasma optical emission spectrometry, as described by Araújo et al. [
33] and Barbosa et al. [
34]. The complete digestion of 0.3 g of dry plant material was achieved with 8 mL of 65% HNO
3 using a microwave digestion system (Multiwave GO; Anton Paar GmbH, Graz, Austria) that followed a two-step heating program: (1) 170 °C reached within 3 min and held for 10 min, and (2) 180 °C reached within 10 min and held for 10 min, followed by cooling. After complete digestion, samples were diluted to 50 mL with ultrapure DI water, filtered with Whatman Grade 1 qualitative filter paper, and stored at 4 °C until analysis. The elemental profile was analyzed by an ICP-OES spectrometer (SPECTRO Genesis, Analytical Instruments GmbH, Kleve, Germany). The contents of mineral nutrients (mg L
−1) were evaluated according to analytical wavebands of 766.491 nm for K, 445.478 nm for Ca, 324.754 nm for Cu, 279.079 nm for Mg, 257.611 nm for Mn, 259.941 nm for Fe, 213.856 nm for Zn, 249.773 nm for B, 213.618 nm for P, and 182.034 nm for S. The operating conditions employed for the ICP-OES were: 1.3 kW RF power, 1.0 L min
−1 auxiliary argon (Ar) flow, 0.80 L min
−1 nebulizer Ar flow, 12 L min
−1 coolant Ar flow, and axial plasma configuration. Each sample was analyzed in triplicate. The calibration standards of mineral nutrients were prepared by diluting a stock multi-element standard solution (1000 mg L
−1) in 6.5% HNO
3 and by diluting phosphorus and standard sulfur solutions (1000 mg L
−1) in ultrapure DI (Merck KGaA). The calibration curves for all the studied elements were in the range of 0.01–400 mg L
−1. The content of each macro- and microelement in the dry weight of the Swiss chard (mg g
−1 DW) was presented.
2.6. Intake Risk Assessment
The average daily intake (mg kg
−1 day
−1) of potentially toxic metals by consuming leaves of Swiss chard after foliar application of ZnO-NPs was calculated by Equation (1): [
35]:
where
Cm means the metal concentration in a plant (mg kg
−1) on a dry weight basis,
Cf is the the conversion factor (0.085) to convert the fresh to dry weight,
IR is the ingestion rate of vegetables, and
Bw is the average body weight for an adult at 70 kg. The average daily intake of Swiss chard was estimated at 100 g (0.1 kg person
−1 day
−1).
The risk of non-carcinogenic health effects is often evaluated from hazard quotients (
HQs), which are the ratio of the daily intake (often the average daily intake,
ADI) to a toxicological reference dose (RfD) according to Equation (2) [
35]:
The oral reference dose (
RfD) for Zn and Fe is 0.3 mg kg
−1 day
−1 [
36].
If the value of HQ is less than 1, it is assumed to be safe from the risk of non-carcinogenic effects. Conversely, if the HQ is equal to or higher than 1, it indicates a potential risk for some of the exposed individuals to experience adverse health effects.
2.7. Statistical Analysis
The data were processed using XLStat software (Addinsoft, USA, 2022). Means were statistically tested using analysis of variance (ANOVA) along with the Tukey’s HSD test. Differences were considered to be significant at p = 0.05.
3. Results
The experimental results revealed that the foliar spray of ZnO-NPs caused no visual damage to Swiss chard leaves and improved fresh and dry biomass, leaf area, affected macro- and microelement uptake, and antioxidative properties. The most significant effect on biomass accumulation in the Swiss chard was achieved after foliar spray with a solution containing 35–45 nm ZnO-NPs (
Table 2). The lowest fresh and dry shoot biomass and leaf area were observed in the untreated plants. Meanwhile, plants exposed to solutions containing 18 nm or 80–200 nm ZnO NPs were similar in above-ground growth.
The chlorophyll index significantly increased in Swiss chard sprayed with ZnO NPs solutions compared to the plants not sprayed by NPs (
Figure 1a). In addition, the flavonols index differed significantly between ZnO NP size treatments; the 35–45 nm treatment resulted in the highest flavonols index, while the 18 nm treatment resulted in the lowest one (
Figure 2a).
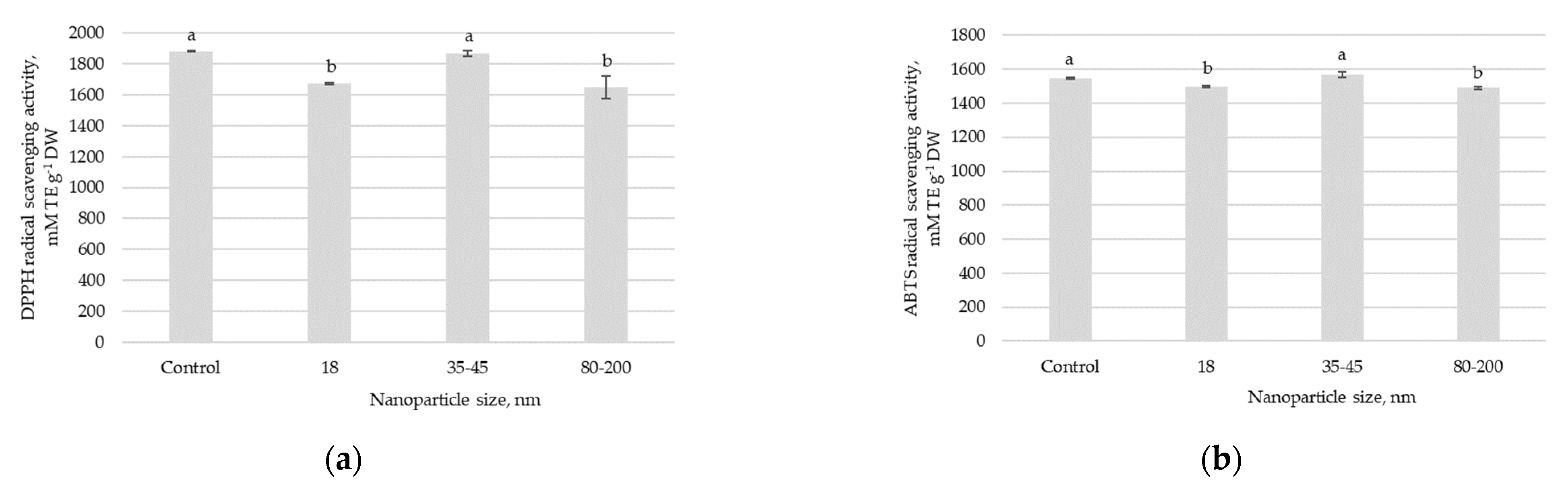
In the Swiss chard, total phenolic compounds showed statistically significant differences between plants sprayed with a solution containing different sizes of ZnO NPs (
Figure 2d). The higher value of total phenolic compounds was found for plants exposed to the 35–45 nm ZnO NPs solution, but there were no statistically significant changes compared to control plants. The foliar spray of 18 nm and 80–200 nm ZnO-NPs caused the reduction of total phenolic compounds in Swiss chard leaves, with the most significant decrease at 80–200 nm. The antioxidant properties of the Swiss chard measured by several methods showed similar responses of antioxidative activity to ZnO NP treatments. There were no statistically significant changes in 2-diphenyl-1-picrylhydrazyl (DPPH) and 2,20-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS) free radical scavenging activities, nor for the Fe
2+ reducing antioxidant power (FRAP) between control plants and plants sprayed with 35–45 nm ZnO NPs (
Figure 2a–c). Significant effects in reducing DPPH, ABTS, and FRAP in the Swiss chard were achieved after foliar spray with solutions containing 18 nm and 80–200 nm ZnO-NPs.
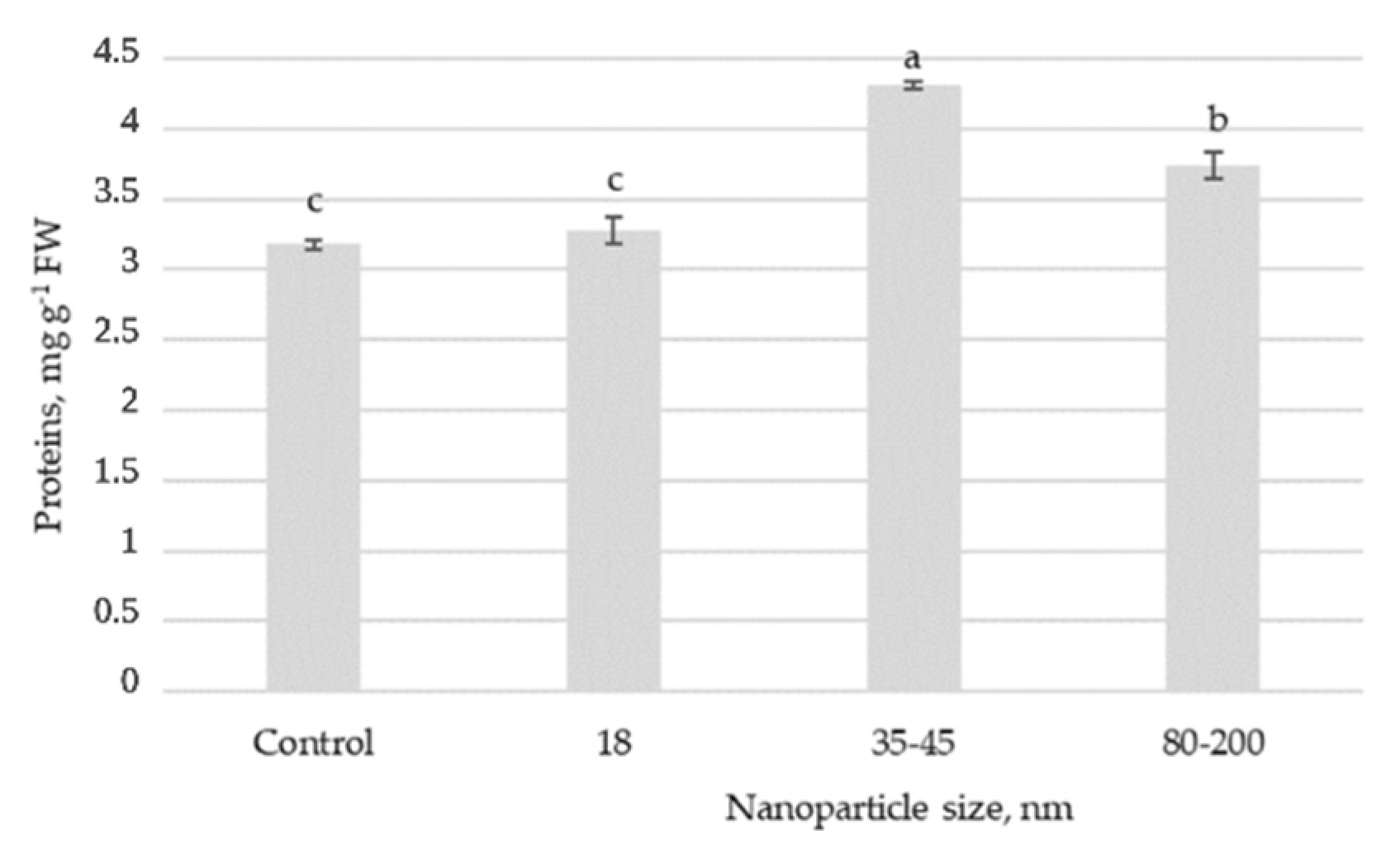
The results of soluble protein content showed statistically significant differences between plants sprayed with solutions containing different sizes of ZnO NPs (
Figure 3). The significantly highest content of soluble proteins in Swiss chard leaves was determined when plants were sprayed with a solution containing 35–45 nm ZnO NPs. Plants sprayed with the 80–200 nm ZnO NP solution also accumulated a higher content of soluble proteins. In comparison, the solution with 18 nm ZnO NPs had no significant effect compared to the untreated plants.
The results from assaying mineral content show that changes in the elemental composition in Swiss chard leaves depended on the size of the ZnO-NP used in the suspensions (
Table 3). Of the macroelements, the level of P significantly increased in the 35–45 nm ZnO NP treatment, as well as Ca, and S under the influence of all ZnO NP sizes, while there was no significant effect on K and Mg. The ZnO NPs of all sizes had a strong positive impact on Zn accumulation in the Swiss chard leaves, increasing Zn levels by 92%, 126%, and 77% with 18 nm, 35–45 nm, and 80–200 nm ZnO NP treatments, respectively. Also, plants exposed to 35–45 nm and 80–200 nm ZnO NPs accumulated significantly higher Fe in their leaves. The content of Mn increased with exposure to 35–45 nm ZnO NPs, while the level of B decreased with any exposure to ZnO NPs.
The foliar treatment of Swiss chard with ZnO-NPs of different particle sizes showed considerable variation in the risk assessment indexes (
Table 4). The results indicated that the Zn ADI in the edible parts of the plants varied, with the highest value in the 35–45 nm treatment, followed by the 18 nm and 80–200 nm. The calculated Zn HQ values were about two times higher for ZnO-NP-treated plants than controls. A health risk assessment was also performed, as Fe is classified as a potentially toxic metal, due to the increased concentration in Swiss chard leaves after treatment with ZnO NPs. Higher Fe ADI values were observed in the Swiss chard after foliar spraying with 35–45 nm and 80–200 nm ZnO NP solutions. HQ values were also higher in these plants compared to untreated plants. Despite this, the HQ values of Zn and Fe were lower than 1, which supports the safe consumption of Swiss chard after ZnO NP treatment.
4. Discussion
Nanotechnology has received considerable attention in recent decades to increase agricultural production rates and plant quality. One example could be using metal nanoparticles (NPs) to biofortify certain trace elements in plants. Of course, it is essential to study the effects of NPs on plants, soil, and humans, as they can be beneficial or toxic. Since the impact of NPs depends on the physicochemical properties of nanoparticles, on the ways of their entry into the plant, and on the selected plant species, this study investigated the effect of spraying different sizes of ZnO NPs on Swiss chard. The previous research demonstrated that using NPs for plant growth depended on the NP characteristics and plant species [
12].
Regarding ZnO NP treatments, various studies in the literature confirm different plant growth responses to ZnO NP application. It usually depends on NP size, concentration, exposure time, and plant species [
18,
19,
20]. The results above confirm that the Swiss chard response to the foliar application of ZnO NPs depended on the NP size used in solutions. Small sized and easily soluble ZnO NPs with high specific surface areas allowed plants to quickly absorb the Zn released from them, thus meeting plant nutritional needs and promoting their growth and development [
37,
38].
Literature data also showed that NPs smaller than the pores could enter and function in plant tissues. For the large size of NPs, there are two potential ways to enter into the plant; one is that the NPs may induce the formation of new pores, which may be a bit larger than usual [
39]. Different characteristics and surface charges of ZnO NPs could affect their further interaction with cells and impact various physiological functions in plants. In addition, the cell wall has hydrophobic and hydrophilic components with unequally distributed fixed negative charges (cellulose fiber (−15 mV) and lignin (−45 mV)) [
40,
41]. Such an ion-exchange surface potentially favors the penetration of cationic rather than anionic NPs. As we can see in our results obtained by measuring the parameters of suspensions, particles with a size of 35–45 nm had the most robust cationic surfaces. Therefore, their suspension was the most related to the surface charge of the leaf and could more easily enter through the Swiss chard leaves. Naturally, Zn binds to insoluble complexes such as Zn-phytate or soluble compounds with proteins, which move in the xylem. Unbound Zn ions remain cationic, can form metal complexes, and move easily in the phloem [
42]. The experiment revealed that the foliar spraying of ZnO-NPs caused superior growth performance, affected leaf chlorophyll and flavonol indexes, increased total soluble protein content, and shifted macro- and microelement accumulations in the Swiss chard leaves. The total phenolic compounds and antioxidant properties in the Swiss chard were dependent on the applied solution containing different ZnO NP particle sizes.
Thus, in this study, different particle size treatments of ZnO NPs significantly impacted Swiss chard biomass accumulation compared to untreated plants. The most significant effect on biomass accumulation in the Swiss chard was achieved after foliar spray with a solution containing 35–45 nm ZnO-NPs. The growth parameters of these plants, in terms of fresh leaves and dry biomass, assimilating leaf area, and fresh roots and dry biomass showed up to 80%, 110%, 76%, 123% and 92% higher values compared to untreated plants, respectively. In agreement with our results, several reports have shown a positive effect of ZnO NPs on radish sprout growth [
22], spinach leaf morphology, biomass and color [
23], parsley, cilantro, and spinach growth and yield production [
20]. Zn plays a vital role in activating enzymes related to protein biosynthesis and carbohydrate metabolism, leading to higher plant biomass. Furthermore, Zn is necessary for activating many enzymes in photosynthetic tissues and is essential for chlorophyll formation [
22,
43]. This is confirmed by our results, because we found a high chlorophyll index in Swiss chard leaves after spraying with solutions containing ZnO NPs. This suggests that the growth and yield promotion of Swiss chard supplied with ZnO NPs could be related to leaf photosynthetic pigments and the photosynthesis rate [
22], as chlorophyll pigments acting as light-capturing centers play an essential role in photosynthesis [
21] and are an important indicator of photosynthetic efficiency for excellent plant growth. Our results are consistent with previous results reported that Zn positively affected the photosynthetic leaf pigments and photosynthesis parameters [
21,
22,
44]. The observed increases in chlorophyll content could be attributed to the essential role of Zn in plant metabolism by affecting the activity of key enzymes, such as carbonic anhydrase, which are Zn metalloenzymes that facilitate the utilization of carbon dioxide at plant carboxylation sites [
21,
44]. In addition, the regulation of photosynthetic pigments and total photosynthesis is associated with reduced oxidative damage caused by abiotic stress [
45,
46].
Understanding the plant response to ZnO NP treatment is of practical importance for plant growth and nutritional quality. Our results showed that, foliar spraying of ZnO NPs disrupted the secondary metabolism of Swiss chard plants and initiated phytochemical changes in leaf tissues. The effects of ZnO NPs depended on the particle size; 18 nm- and 80–200 nm-sprayed plants showed significantly reduced phenols, while 35–45 nm had no significant effect on total phenolic compounds in Swiss chard leaves compared to untreated plants. Applying ZnO NPs caused decreased phenolic compounds in Swiss chard leaves, resulting in reduced antioxidant activities in terms of DPPH, ABTS, and FRAP. This possibly indicates that the treatment with ZnO NPs (particle size 18 nm and 80–200 nm) reduced the induction of non-enzymatic antioxidant compounds in Swiss chard leaves. It is also noticeable that the surface charge of these NPs in the suspension was neutral, which could have determined that they reacted differently than cationic particles when they got inside the plant and thus caused a negative effect on the antioxidant activity of plants. Our findings agree with previous studies reporting the negative effect of ZnO NPs on the total phenols, flavonoid, β-carotene and lycopene contents in tomato fruits [
47]. Results obtained in other studies showed that applying ZnO NPs increased the non-enzymatic antioxidant molecules, such as phenolic compounds and flavonoids, in
Brassica nigra [
48], which generated the highest accumulation of phenolic compounds, and increased the antioxidant activities in
Capsicum chinense Jacq. [
44] and
Capsicum annuum L. [
49], as well as increased the total phenolic compounds in potato plants [
50]. The increases were related to NP-induced oxidative stress, which led to greater activation of secondary metabolism in plant tissues. Plants have different defense mechanisms to reduce oxidative damage caused by ROS through the production of antioxidants such as phenols, carotenoids, and antioxidant enzymes [
2]. In our study, an adequate level of ZnO NPs did not induce oxidative stress in Swiss chard tissues, leading to stable metabolism, resulting in increased production of secondary metabolites and antioxidant activity. However, we cannot compare them, as no study provides a zeta potential to support our results. In addition, most studies do not consider that in suspension or soil, NPs can react to change their shape, charge, and surface area. Therefore, it is essential to not only measure pure and dry NPs.
Compared to untreated plants, foliar spraying with ZnO NPs enhanced the total protein content by 36% and 20% when 35–45 nm and 80–200 nm ZnO NPs were applied, respectively. Similarly, ZnO NPs significantly increased the leaf total soluble protein content in cotton [
51], green peas [
14], pearl millet [
52] and cluster beans [
53]. These results indicate that Zn is required as a structural and catalytic component of proteins and enzymes for normal plant growth and development [
7]. Also, it was reported that proteins protected the cells from oxidative damage caused by NP deposition in plants [
51].
Macro- and microelements are vital nutrients in plant tissues involved in various activities that ensure normal plant growth and development. Our research results showed that the application of ZnO NPs significantly affected Ca, S, and B content in Swiss chard leaves. The positive interactions in response to increasing Zn levels were found for Ca and S, while they were negative for B. Zn is known to interfere with plant uptake of B. Zn has even been suggested as a means of reducing B toxicity in crops grown in boron-rich soils. For Ca and S, positive and negative interaction effects have been reported in crop plants, suggesting different mechanisms in various plant species [
54,
55]. Publications report that Ca blocks the uptake of Zn in plants through roots, as it is its antagonist. However, in this study, we treated plants with ZnO NPs through the leaves, thus reducing competition for their entry through the roots [
56]. This may have accounted for the higher Ca content in the Swiss chard tissues. Although different results have been reported for the association of S with Zn, the exact mechanism is unclear. For example, in rice seedlings, Zn content was reduced by S, and in the mustard study, Zn content was increased by S.
The presence of mineral elements in acceptable and safe amounts for human health significantly correlates with the nutritional quality of plants. The results of the current study revealed a considerable uptake of Zn in Swiss chard leaves via the foliar pathway. Previously, it has been shown that ZnO NPs easily penetrated the leaf surface, and plants accumulated a high content of Zn [
57,
58]. In Swiss chard leaves, ZnO NPs with a particle size of 35–45 nm had the most substantial effect on Zn accumulation compared to plants treated with solutions containing 18 nm or 80–200 nm ZnO NPs. The differences in zeta potential may have caused the different release of Zn ions and their translocation in the leaves [
27]. Furthermore, the high levels of Zn detected in the Swiss chard leaves may be associated with the alteration in gene expression corresponding to metal homeostasis and binding in plant leaves [
59]. Improvement in the adequate amount of essential minerals, especially Zn and Fe, provides a better source of these nutrients for human consumption and a critical factor in evaluating the food quality of leafy greens [
20]. Compared to untreated plants, the presence of ZnO NPs increased Fe content in edible Swiss chard tissues by about 80% and 45% when plants were sprayed with solutions containing 35–45 nm and 80–200 nm ZnO NPs, respectively. Sharifan with co-workers [
20] found that 100 mg L
−1 of ZnO NPs increased the Fe content in spinach and cilantro, but Fe accumulation was unaffected in parsley. The natural resistance-associated macrophage protein members (NRAMP3-4) and metal tolerance proteins (MTP1-8) are transporters of the essential metals, exporting them via vacuoles, which also regulate Fe translocation, and the sequestration of Zn content drives their function in the tonoplast [
60]. Whereas the sequestration of Zn occurs through ZIP, NRAMP, and YSL transporters, high levels of Zn may cause the overexpression of essential minerals transporters, leading to a change in the efflux. The concentration of Zn and Fe in the edible parts of the Swiss chard supplied with ZnO NPs did not reach the toxicity level according to the health risk index data (
Table 3). In addition, the leaf Zn and Fe health risk index did not exceed the allowable limit established by the Food and Agriculture Organization/World Health Organization (FAO/WHO) (less than 1). It is notable that all treated plants with ZnO NPs were safe for consumption, and there was no apparent risk associated with heavy metal toxicity. In addition, the increase of Zn and Fe in the edible tissue of Swiss chard were below tolerable daily intake limits of Zn (99.4 mg kg
−1) and Fe (425.5 mg kg
−1) according to the (FAO/WHO) guideline.
Over the past few decades, consumers have become more health-conscious and are consuming more vegetables, which are a source of various health-beneficial compounds, including minerals. However, it is estimated that over 2 billion humans of the world’s population are deficient in Fe and Zn [
61]. This situation is practically related to the cultivation of plants in regions with low mineral availability for growing plants. Also, food deprivation for humans is another crucial factor. In this context, using nanoparticles for various purposes to reduce the adverse effects of their deficiency in edible plant tissues could be considered a sustainable method to adjust their ratio in plants and improve nutritional quality.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}