Abstract
Proline (Pro) is not only an important osmotic adjustment substance, but it also plays an important role in regulating plant abiotic stress resistance. The maize varieties, Xinxin 2 (low temperature insensitive) and Damin 3307 (low temperature sensitive), were chosen as materials, setting a normal temperature for germination (22 °C/10 °C, 9d), low temperature germination (4 °C/4 °C, 5d) and normal temperature recovery (22 °C/10 °C, 4d), combined with a proline (15 mmol·L−1) soaking treatment, to reveal the seed germination and regulation mechanism in maize embryos. The results showed that proline significantly improved the germination potential, germination rate, germination index and vigor index of seeds under low temperature stress, increasing the length of the coleoptile and radicle, increasing the dry and fresh weight of young buds (coleoptile + radicle), and increasing the activity of α-amylase. Proline enhanced the activities of Δ1-pyrroline-5-carboxylic acid synthetase (P5CS) and ornithine aminotransferase (OAT) in maize embryos under low temperature stress, enhanced the proline synthesis pathways, and further enhanced proline accumulation. Proline induced the activity of proline dehydrogenase (ProDH) in the early stage of low temperature stress and stress relief. Under low temperature stress, the activities of hexokinase (HXK), phosphofructokinase (PFK), pyruvate kinase (PK), isocitrate dehydrogenase (IDH), succinate dehydrogenase (SDH), malate dehydrogenase (MDH) and glucose-6-phosphate dehydrogenase (G-6-PDH) and glucose-6-phosphate dehydrogenase (G-6-PDH and 6-P-GDH) in maize embryos were decreased, resulting in a decrease in the glycolysis (EMP) pathway, tricarboxylic acid (TCA) cycle and pentose phosphate pathway (PPP). Proline alleviated the inhibition of key enzyme activities of the EMP pathway, TCA cycle and PPP of maize embryos under low temperature stress, increased the activities of cytochrome oxidase (COX) and alternative oxidase (AOX), increased the ATP content, alleviated the inhibition of low temperature stress on main cytochrome pathway activity (ρVcyt), while further increasing the total respiratory activity (Vt) and the actual operational activity of the alternative pathway (ρValt) during seed germination at the initial stage of low temperature stress, as well as improving the inhibition of the Vt and ρValt in the middle and late stages of low temperature stress. Under low temperature stress, the content of abscisic acid (ABA) increased significantly, while gibberellin (GA), auxin (IAA) and zeatin nucleoside (ZR) decreased significantly. Proline alleviated the decrease in IAA, ZR and GA contents in maize embryos under low temperature stress, reduced the increase in the ABA content, and reduced the inhibition of low temperature on seed germination.
1. Introduction
The world population is expected to exceed 9.7 billion in 2050, and the current food growth rate has been unable to meet the needs brought by global population growth [1]. China is the most populous country in the world and the second largest maize producer in the world. The high and stable yield of maize plays an extremely important role in ensuring food security and the sustainable development of animal husbandry [2]. Temperature is closely related to plant growth and development [3,4]. Under the background of global warming, abnormal temperature changes also occur at the same time, the frequency of extreme weather intensifies, and the probability of crops suffering from low temperature stress increases, and these problems may become more and more serious in the 21st century [5,6,7,8,9]. Maize may suffer from low temperature stress in all growth stages [10,11,12,13]. The major maize growing areas around the world, such as the United States, China, Canada, Europe, Central Asia, Russia and other maize growing areas at high latitudes and altitudes, are facing different degrees of low temperature threat [14,15]. Low temperature stress has been one of the main environmental factors restricting the high yield of maize in these areas [16]. Previous studies have shown that severe low temperature stress occurs every 3–5 years in China, especially in Northeast China [17,18]. The frequency and intensity of low-temperature injury tend to increase, and the yield can be reduced by more than 15% in serious years [19,20]. In order to ensure the normal germination, emergence, growth and development of maize and to finally achieve a stable yield, it is imperative to improve the low temperature tolerance of maize [21,22].
Under low temperature, plants will accumulate a series of osmoregulatory substances to resist the effects of stress, especially proline. Under abiotic stress, proline can account for up to 80% of the cellular amino acid pool [23], while proline accumulation can be detected in various tissues undergoing rapid cell division and elongation [24], for example, proline accumulated in immature broad bean seeds and the hypothesis was put forward that proline may be important for the development of reproductive organs [25]. Genes encoding the key enzymes of proline metabolism have been successively found in Arabidopsis, tomato, alfalfa and rice, which are expressed during seed development [26,27]. The expression of antisense AtP5CS1, for example, led to the delay of the radicle in the process of Arabidopsis seed germination, which proved the evidence of proline synthesis in seed germination [28]. Exogenous proline can promote cell division and alleviate abnormal embryonic development by up-regulating the expression of the cyclin CYCB1:1 gene [29,30]. The homozygous grains of the mutant not only showed the collapsed form of a powdery endosperm, but also showed lethality at the two-leaf stage. The exogenous addition of proline can restore its normal development, therefore it is named proline responding 1(pro1). The Pro1 gene encodes P5CS2 through positional cloning and the functional identification of the maize Pro1 gene [31]. The loss of P5CS2 function in the maize mutant reduces the synthesis of proline and the lack of proline causes the accumulation of uncharged tRNApro AGG in cells, resulting in a general reduction in protein synthesis and proline deficiency affecting the accumulation of plant protein [32]. These studies point out that proline plays an important regulatory role in protein synthesis and cell cycle transformation in higher plants. Exogenous proline can improve the germination rate of cotton, pea and wheat seeds, reduce the germination time and promote the growth of seedlings under osmotic stress [33]. Meanwhile, exogenous proline has promoted wheat seed germination, root growth and seedling development under cadmium stress [34].
Respiration provides power for plant biosynthesis, cell maintenance and active transportation. Respiration is driven by many biochemical pathways, including glycolysis (EMP, located in the cytoplasm), the tricarboxylic acid cycle (TCA, located in the mitochondrial matrix), the pentose phosphate pathway (PPP, located in the cytoplasm), and the electron transport chain (ETC, located in the mitochondrial intima). Respiration and energy metabolism are particularly important for seed germination and both the EMP pathway and PPP are active during seed germination. When mitochondria become active, the PPP is dominant. On the contrary, when the production of mitochondrial ATP is limited by hypoxia, the substrate in the seed will enter the EMP pathway for catabolism after phosphorylation [35]. The plant ETC mainly includes the cytochrome pathway (CP) and alternative pathway (AP). The change of respiratory rate can be divided into two stages; the first is the short-term stress response, and then the change after long-term low temperature stress [36]. A former study showed that after wheat was exposed to a low temperature, the total respiratory activity (VT) initially increased, and then decreased with the extension of the low temperature duration [37]. Previous studies on Arabidopsis and plantain found that the decrease in Vt caused by low temperature is related to the damage of respiratory organs such as mitochondria [38,39]. Hexokinase (HXK), phosphofructokinase (PFK) and pyruvate kinase (PK) are the key enzymes of the EMP pathway. Low temperature stress will reduce the activity and gene expression of HXK, PFK and PK, slow down the decomposition of soluble sugar, improve the content of the plant osmotic regulator and strengthen a plant’s low temperature tolerance [40]. The decrease in SDH activity under low temperature stress is related to the transfer of carbon from the TCA cycle to the PPP, and the enhancement of the PPP can improve the tolerance of plants to low temperature stress [41]. Low temperature stress in the imbibition stage of soybean seeds will reduce the expression of MDH and imbalance mitochondrial metabolism [42]. Low temperature stress reduces the activity of key enzymes involved in the EMP-TCA cycle and promotes the transport of sugar from the endosperm to the embryo to prevent low temperature damage [43].
A low temperature can trigger the synthesis and accumulation of ABA, and then close the stomata by defending the signal transduction on the cytoplasmic membrane, reducing the water loss caused by transpiration and maintaining the water content of cells [44]. When exposed to low temperature stress, the ABA content in leaves of cold tolerant maize seedlings was more significant [45]. Exogenous ABA treatment can increase plant cold tolerance by increasing the content of soluble sugar and proline, reducing ROS accumulation, reducing the membrane lipid peroxidation level and cell membrane damage [46]. Gibberellin (GA) is involved in regulating seed germination, leaf expansion, stem elongation and the flowering of many plant species [47]. Low temperature treatment at 6 °C for four days could significantly reduce the GA3 content in maize seedling leaves and tomatoes [48,49]. Spraying a 500 mg L−1 GA3 solution on the leaf can promote bolting and the flowering of blood tonic grass plants under 6 °C low temperature stress [50]. Exogenous GA3 can also improve the chilling tolerance of peach fruit and reduce chilling injury by activating the antioxidant enzyme system and increasing the expression of GSK-3 [51]. Previous studies have shown the IAA content in cotton seeds germinated under low temperature stress [52]; however, the IAA content remained high in tea leaves with a naturally low temperature [53]. This indicates that the response of IAA under low temperature stress is related to the plant species and stress degree. Maintaining a high level of IAA in cells in a low temperature environment is conducive to improving the cold tolerance of plants [54]. Zeatin nucleoside (ZR) is activated during seed germination, which can promote cell proliferation, cell division, seed germination and inhibit leaf senescence [55]. During a low temperature treatment, the content of cytokinin decreased and the activity of cytokinin oxidase increased [56], while a low temperature treatment in the early stage of grain filling significantly reduced the ZR content, resulting in a reduction in the grain storage capacity. The above results show that a low temperature can change the level of plant hormones and delay the process of the cell cycle [57].
Proline is a common, soluble, small molecule organic matter in plants, and it is also one of the components of plant protein [58]. Under stress conditions, proline will accumulate in large quantities [59,60,61] and proline is generally considered to be a beneficial osmoregulation substance, which can protect the membrane integrity and stabilize the protein and subcellular structure [62]. Studies have shown that proline can also protect cell function by maintaining an appropriate NADP +/NADPH ratio and reducing reactive oxygen species (ROS) [63]. After stress relief, proline catabolism can also provide carbon and nitrogen sources and energy for plants [64]. Proline is easy to obtain and cheap, which shows that proline has practical value in protecting plants from the impact of a stress environment and accelerating the recovery of growth and development after stress [65]; however, the relationship between the changes of proline metabolism and plant low temperature tolerance and its regulation mechanism on plants have not been studied. Therefore, the study on the mitigation effect of exogenous proline on maize seed germination under low temperature stress, as well as the regulation mechanism of proline metabolism and the mechanism of improving maize low temperature tolerance can provide a theoretical basis and practical value for the application of proline in seed treatment.
2. Materials and Methods
2.1. Experimental Varieties
The experiment was carried out in the key laboratory of cold region grain crop variety improvement and physiological ecology of the ministry of education, Northeast Agricultural University. Based on the results of preliminary experiments, we selected the maize varieties, Xinxin 2 (low temperature insensitive, XX 2) and Damin 3307 (low temperature sensitive, DM 3307), as the experimental materials. The proline was purchased from Sigma (C5H9NO2, CAS No.147-85-3).
2.2. Experimental Design
2.2.1. Proline Concentration Screening Experiment
Two varieties of maize seeds with similar plumpness and uniformity were used in the experiment. After disinfecting with a 0.1% HgCl2 solution for 10 min, the seeds were washed repeatedly with distilled water, and then the water on the seed surface was wiped dry with filter paper. The seeds were placed in a 500 mL beaker, submerged in 0, 5, 10, 15 and 30 mmol·L−1 proline solution, respectively, and soaked in 22 °C artificial intelligence incubators (HPG-280HX) for 24 h. After soaking, the seeds were taken out and washed with distilled water. After natural drying, they were arranged in a 15 cm Petri dish paved with two layers of filter paper. Each Petri dish was covered with 30 seeds and a layer of filter paper. Each Petri dish was sprayed with 20 mL of distilled water.
The processing consisted of two parts. (1) Low temperature treatment: the seeds soaked in 0, 5, 10, 15 and 30 mmol·L−1 proline solutions were placed in a 4 °C/4 °C artificial intelligence incubator for low temperature treatment for 5 d, and then germinated in a 22 °C/10 °C artificial incubator. There were five treatments, which were recorded as L, L + P1, L + P2, L + P3 and L + P4, respectively; (2) In the normal temperature treatment, the seeds soaked in 0, 5, 10, 15 and 30 mmol·L−1 proline solutions were always placed in a 22 °C/10 °C artificial intelligence incubator for germination. There were five treatments, which were recorded as CK, P1, P2, P3 and P4, respectively. All germination tests were conducted in dark conditions, repeated every three times, supplemented with an appropriate amount of distilled water every day to ensure that the seeds were wet, and the filter paper was changed every two days. On the 3rd, 5th, 7th, 9th, 11th and 13th days of treatment, we counted the number of germinated seeds (the germination standard is that the radicle length exceeds the seed length and the germ length exceeds half of the seed length), the low temperature treatment was counted to 13 d and the normal temperature treatment was counted to 9 d. The complete seed embryo, radicle and coleoptile were taken for the determination of relevant physiological indexes after low temperature treatment and normal temperature treatment for 13 d and 9 d, respectively.
2.2.2. Germination Experiment of Seeds Soaked with Proline under Low Temperature Treatment
The seed soaking solution was a 0 mmol·L−1 (distilled water) and 15 mmol·L−1 proline solution. The other operations were the same as Section 2.2.1. The seeds after soaking were used in different treatments including (1) control group (CK), which germinated at 22 °C/10 °C for 9 days after soaking in distilled water (the 0th day); (2) proline seed soaking treatment (P), the 15 mmol·L−1 proline solution always germinated at 22 °C/10 °C for 9 days; (3) low temperature stress treatment (L), after soaking seeds in distilled water, with the seeds germinated at 4 °C/4 °C for 5 days, and then transferred to 22 °C/10 °C for 4 days; (4) low temperature stress and proline combined treatment (L + P), after soaking seeds in 15 mmol·L−1 proline solution, with the seeds germinated at 4 °C/4 °C for 5 days, and then transferred to 22 °C/10 °C for 4 days. All germination was carried out under dark and humid conditions, and the treatment was repeated every 3 times. An appropriate amount of distilled water was added every day, and the filter paper was changed every 2 days. At the 0, 1, 3, 5, 7 and 9 days of each treatment, we carefully peeled the complete seed embryo for the determination of relevant physiological indexes.
2.3. Measurement and Methods
2.3.1. Determination of Germination Potential, Germination Rate, Germination Index and Vigor Index
Germination potential = total number of germinated seeds/total number of tested seeds × 100% (number of germinated seeds on the 9th day of low temperature treatment and the 5th day of normal temperature treatment).
Germination rate = total number of germinated seeds/total number of tested seeds × 100% (number of germinated seeds on the 13th day of low temperature treatment and the 9th day of normal temperature treatment).
Germination Index = , where Gt is the number of germinated seeds on day t, and Tt is the number of days.
where S is the fresh weight of seedlings (g).
Vigor Index = GI × S
2.3.2. Determination of Coleoptile Length, Radicle Length and Dry-Fresh Weight of Young Buds
On the 9th day of normal temperature treatment and the 13th day of low temperature treatment, we randomly took 10 germinated seeds and measured the length of the coleoptile and radicle with a ruler. After analyzing and measuring the fresh weight of the bud (coleoptile + radicle), we overheated the bud at 105 °C for 30 min, and then dried it at 80 °C to constant weight to measure the dry weight.
2.3.3. Determination of α-Amylase Activity
We took a 0.5 g seed embryo and placed it into a pre-cooled mortar and added a 0.1 M sodium citrate buffer (pH 5.5) to grind it into a homogenate. Then the homogenate was centrifuged at the speed of 10,000× g at 4 °C for 15 min, and the supernatant collected after centrifugation was used for the determination of α-amylase activity. The reaction mixture contained 0.5 mL of 1% starch, 0.25 mL of a 0.1 M citrate buffer (pH 5.5) and 100 μL supernatant, and then the reaction mixture was bathed in water at 50 °C for 20 min. Finally, we added 3 mL of a 1% (v/v) DNS solution, shaken well and boiled in a boiling water bath for 5 min. The absorbance value of the cooled solution was read with a spectrophotometer at 550 nm, and the unit of enzyme activity was mg·g−1·min−1 [66].
2.3.4. Determination of Proline Content
We weighed 0.2 g of seed embryo and placed it into a mortar, added 5 mL of 3% sulfosalicylic acid, ground it into a homogenate, placed it into a test tube, covered it with a boiling water bath for 10 min and cooled it for 11,500× g centrifugation for 15 min. We took 2 mL of the supernatant in the test tube, mixed it with 2 mL of acid ninhydrin and 2 mL of glacial acetic acid in the test tube, shook it well, then boiled it again for 1 h, and then cooled it naturally to room temperature. We added 4 mL of toluene into the test tube, shook it fully, took the upper toluene extract in the cuvette to measure the absorbance in OD520, and finally calculated the content of proline [67].
2.3.5. Determination of Enzymes Activities of P5CS, OAT and ProDH
The activity of the P5CS was determined according to the improved method of Garcia Rios et al. [68]. The whole extraction process of crude enzyme solution was carried out at 4 °C. We took a 0.2 g seed embryo sample, added liquid nitrogen and ground it into a powder in a pre-cooled mortar. Then we added a 2 mL 100 mm Tris HCl buffer (pH 7.5), 10 mm into the mortar β-Mercaptoethanol, 2% PVPP, 10 mM MgCl2, 1 mm PMSF and 4 mm DTT. After mixing it evenly, we used 12,000× g centrifuged for 20 min, and the supernatant was taken for enzyme activity determination. The reaction mixture (4 mL) contained a 50 mm Tris HCl buffer (pH 7.0), 50 mM glutamate, 20 mM MgCl2, 1 mm NADPH, and 10 mM ATP. We started the reaction by adding the enzyme extract, first reacted at 37 °C for 20 min, and then added 2 mL TCA to stop the reaction. Finally, we measured the absorbance of the supernatant OD338, and the unit of P5CS activity was nmol·mg−1·min−1 protein.
We took a 0.2 g seed embryo sample, added liquid nitrogen into the pre-cooled mortar and ground it into powder. Then, we added 2 mL of extract to the mortar, including 100 mM potassium phosphate buffer (pH 7.4), 1 mm pyridoxal 5-phosphate, 1 mm EDTA, 10 mm β-mercaptoethanol, 1% PVP, 5 mM MgCl2 and 0.6 M KCl. After the above solutions were mixed evenly, we used a 12,000× g centrifuge for 20 min. The supernatant obtained after centrifugation was used to determine the activities of the ProDH and OAT. We measured the OAT activity according to Vogel and Kopac’s method with slight changes. A 1 mL amount of the reaction mixture consisted of 50 mM ornithine, 20 mM α-ketoglutarate, 1 mM pyridoxal phosphate, 100 mM phosphate buffer (pH 8.0) and enzyme extract. The mixture was reacted at 37 °C for 30 min and the reaction was stopped by adding 0.5 mL of 10% trichloroacetic acid. Finally, 0.5 mL of chromogenic solution (0.5% o-aminobenzaldehyde, 95% ethanol) was added in this tube, and a 12,000× g centrifuge was completed for 10 min, with the transparent supernatant measured at OD440. The unit of OAT activity was μmol·mg−1·min−1 protein [69].
The activity of the ProDH was determined according to the method described by Yang et al. [70]. The reaction mixture (3 mL) contained a 100 mM Na2CO3-HCl buffer (pH 10.3), 50 mm proline, 0.4 mM NADP+ and enzyme extract. The absorbance value of the OD340 was measured, and the activity of the ProDH was expressed by NADP+ consumption. The unit of ProDH activity was nmol·mg−1·min−1 protein.
2.3.6. Determination of Vt, ρValt, and ρVcyt
A 0.1 g embryo sample was added to 10 mL of reaction medium (50 mM pH 6.8 phosphate buffer containing 0.1 M sucrose). The oxygen consumption rate in the solution was measured with a Clark oxygen electrode at 25 °C. The inhibitors used were 5 mM potassium cyanide (KCN) and 10 mM salicylhydroxamic acid (SHAM). Total respiratory activity (Vt) was measured without adding any respiratory inhibitors. The alternative pathway actual operation activity (ρValt) was calculated by subtracting the respiratory activity of sham only from the Vt. Cytochrome main pathway activity (ρVcyt) was calculated by subtracting the respiratory activity when only SHAM was added from the respiratory activity when the KCN and SHAM were added at the same time. The above respiratory rate was expressed in nmol·O2·g−1· FW·h−1 [71].
2.3.7. Determination of Enzymes Activities of COX and AOX
The extraction process of the mitochondria was as follows. A 1 g embryo sample was added to 5 mL pre-cooling extract (60 mM sucrose, 5 mM EDTA, 0.2% (w/v) BSA, 0.3% β-mercaptoethanol, 50 mM Tris-HCl buffer, pH 7.4), ground into a homogenate, and then a 3000× g centrifuge completed for 10 min (4 °C) before the supernatant was collected. We added 5 mL of pre-cooling extract to the precipitate in a 10,000× g centrifuge for 10 min (4 °C) and collected the supernatant. The supernatant was collected twice, 11,000× g centrifuged for 10 min (4 °C), and the precipitate was added into a 1 mL pre-cooling extract to a 11,000× g centrifuge for 10 min (4 °C) then resuspended. Finally, we added a 3 mL preservation solution (50 mm pH 7.4 potassium phosphate buffer and 2.0% Triton X-100) to the sediment and stored it at −80 °C for standby. All extraction steps were carried out at low temperature.
COX activity was determined according to the following method. A 3 mL amount of the reaction mixture contained the following, 50 mM phosphate buffer (pH 7.1), 0.2 M sucrose, 20 M reduced cytochrome C and 25 μL enzyme solution. The reaction was started by adding the enzyme extract, and the absorbance value of the reaction mixture at 550 nm every 30 s was measured and recorded for 5 min. The unit of COX activity was nmol·mg−1·min−1 protein [72].
AOX activity was determined according to the following method. The reaction mixture contained 0.6 mM sucrose, 10 mM trimethylolmethylaminoethanesulfonic acid (pH 7.2), 5 mM KH2PO4, 10 mM NaCl, 2 mM MgSO4 and 0.1% (w/v) BSA. All measurements were carried out at 25 °C, and the oxygen consumption was measured with a Clark oxygen electrode. We added 0.1 mM ATP, 0.5 mm KCN and 20 μL crude enzyme solution to the reaction mixture in advance. After the reading in the reaction system remained stable, 10 mM sodium succinate, 0.5 mm pyruvate, 1 mm ADP and 10 mm DTT were added, and the 5-min oxygen consumption rate was read quickly. The unit of AOX activity was nmol O2·mg−1·min−1 protein [73].
2.3.8. Determination of Enzymes Activities of HXK, PFK and PK
The extraction process of crude enzyme solution was as follows. A 1.0 g embryo sample was added to 10 mL 50 mM potassium phosphate buffer (pH 7.4) and ground into a homogenate with a pre-cooled pestle and mortar under ice bath conditions. The homogenate was then homogenized at 7000× g and 4 °C for 10 min, the supernatant was centrifuged again for 10 min under the same conditions, and the supernatant was then stored at 4 °C for testing. The reaction mixture contained 0.8 mL 0.1 M Tris HCl (pH 8.5) buffer (containing 5 mM DTT, 10 mM MgCl2, 10 mL glucose), 1U glucose-6-phosphate dehydrogenase, 20 μL 100 mM ATP, 20 μL 500 mM NADP+ and 0.15 mL crude enzyme solution. We started the reaction by adding the enzyme solution and measured the absorbance at OD340. We then poured the colorimetric solution into the test tube again, quickly returned it to the cuvette after 2 min of a water bath at 37 °C and measured the absorbance at OD340 again. An amount of 1 mmol NADPH was generated per minute as one HXK activity unit (U) [74]. Phosphofructose kinase (PFK) can catalyze fructose-6-phosphate (F6P) and ATP to produce fructose-1,6-diphosphate (FDP) and ADP, and then pyruvate kinase (PK) catalyzes NADH to finally produce NAD +, which reflects the PFK activity by measuring the decline rate of the NADH. The absorbance reduction value was measured in OD340 within 10 min, and the consumption of 1 mmol NADH per minute was taken as a PFK activity unit (U). Pyruvate kinase (PK) can catalyze phosphoenolpyruvate (PEP) and ADP to produce pyruvate. The PK activity was reflected by measuring the conversion of pep to pyruvate [73,74].
2.3.9. Determination of Enzymes Activities of IDH, SDH and MDH
The 1 g embryo samples were placed in a pre-cooled mortar, and 4 mL extract was added for ice bath grinding. The extract contained 50 mM HEPES-NaOH(pH 7.5), 5 mM MgCl2, 5 mM EDTA-Na2, 10% (v/v) glycerol, 0.1% (v/v) TritonX-100 and 2% (w/v) PVP-10, while 12,000× g was centrifuged for 5 min at 4 °C, and the supernatant was then taken for the determination of the IDH and MDH activities [75].
The IDH activity analysis refers to Gajewska’s method. The reaction mixture (1 mL) consisted of 100 mM MOPS(pH 7.4), 0.5 mM MgCl2, 0.5 mM NADP, 2.5 mM isocitric acid and 0.25 mL enzyme extract. The reaction was initiated by adding the enzyme solution, and the change of absorbance at 340 nm within 2 min was recorded. The activity of one unit was defined as the amount of enzyme (U) producing 1 nmol NADPH [76].
MDH activity was determined according to the method of Kumar et al. [77]. The reaction mixture (1 mL) consisted of a 100 mM Tris-HCl buffer (pH 7.8, containing 5 mM MgCl2 and 1 mM EDTA), 5 mM oxaloacetic acid, 0.8 mM NADH and enzyme extract. The initial absorbance at 340 nm and the absorbance at 1 min were recorded, and the amount of enzyme oxidizing 1 nmol NADH per minute was one activity unit (U) of MDH.
The activity of succinate dehydrogenase (SDH) was determined according to the method of Cooper et al. [78]. The extraction method of the enzyme solution was the same as that of the COX enzyme solution, but the temperature was controlled at 25 °C. SDH activity was determined by a reduction in Dichlorophenol indophenol sodium (DCPIP) resulting in a decrease in absorbance at 600 nm. The reaction mixture (3 mL) contained a 30 mM potassium phosphate buffer (pH 7.8), 1 mM NaN3, 10 mM sodium succinate, 0.1 mM 5-methylphenazine methyl sulfate (PMS), 0.008 mM DCPIP and 0.1 mL enzyme solution. The reaction was initiated by adding the enzyme solution. The change of absorbance value of the reaction mixture at 600 nm was measured at 25 °C. SDH activity was defined as reduction 1 μmol DCPIP within 1 min (U).
2.3.10. Determination of Enzymes Activities of G-6-PDH and 6-P-GDH
The extraction method of crude enzyme solution was consistent with that in Section 2.3.9. The reaction solution contained a 100 mM Tris HCl buffer (pH 7.4), 5 mM NADPNa2, 5 mM D-glucose-6-phosphate disodium and 5 mM NADPNa2. We added 1.35 mL of reaction solution to 0.1 mL of crude enzyme solution, immediately measured the absorbance value at 340 nm, and then recorded it every 1 min for a total of 5 times. In the calculation, the OD change of 0.1 was taken as an enzyme activity unit (U), and the enzyme activity unit was U·g−1 FW [79].
2.3.11. Determination of ATP Content
The ATP content was determined by spectrophotometry. ATP and creatine phosphocreatine were catalyzed by creatine kinase. The content of ATP was determined by phosphomolybdic acid colorimetry at OD700.
2.3.12. Determination of Contents of ABA, IAA, GA and ZR
The contents of ABA, IAA, GA and Zr were determined by an enzyme-linked immunosorbent assay (ELISA). A 0.5 g maize embryo was ground into powder with liquid nitrogen, and then 2 mL of the sample extract (containing 80% methanol and 1 mM butyl hydroxytoluene mixture) was added to the mortar and ground into a homogenate under an ice bath. We transferred the homogenate obtained above into a 10 mL test tube, rinsed the mortar with 2 mL of extract, and transferred it into the test tube together. After shaking the test tube, it was placed in a 4 °C refrigerator for 4 h. The test tube sample was centrifuged at 10,000× g for 8 min, and then the supernatant was taken. After centrifuging for 1 h, 1 mL of the waste solution was added to the refrigerator and we recorded the volume of the waste solution, then added it to the test tube for extraction. The collected supernatant was passed through a C-18 solid phase extraction column. The specific steps were as follows: balanced the column with 80% methanol (1 mL), loaded the sample, collected the sample, washed the column with 100% methanol (5 mL), washed the column with 100% ether (5 mL), washed the column with 100% methanol (5 mL), and recycled again. Then, the sample was transferred after passing the column into a 5 mL centrifuge tube, concentrated and dried in a vacuum, excess methanol removed, and the volume fixed with a sample diluent. We transferred the sample after passing the column into a 10 mL plastic centrifuge tube, concentrated and dried it in a vacuum or blow dried it with nitrogen to remove the methanol in the extract, and then fixed the volume with the sample diluent (the culture containing 0.50 mL Tween-20, 0.50 g gelatin and 500 mL PBS buffer). The ELISA kit was provided by China Agricultural University, and the hormone content was determined by a bio Tek elx-800 automatic microplate reader [80].
2.4. Data Analysis
According to the analysis of variance, the data were statistically analyzed following standard methods using Microsoft Excel 2010 and SPSS 12.0. Differences between the treatments were determined by a posteriori Tukey’s test at a significance level of 0.05 and 0.01.
3. Results
3.1. Germination Potential, Germination Rate, Germination Index and Vitality Index
It can be seen from Table 1 that compared with the normal temperature control treatment, the low temperature treatment significantly reduced the germination potential, germination rate, germination index and vitality index of the two varieties of maize seeds. The germination potential, germination rate, germination index and vigor index of the DM 3307 seeds decreased by 43.41%, 33.15%, 48.42% and 64.53%, respectively. The germination potential, germination rate, germination index and vigor index of the XX 2 seeds decreased by 25.99%, 18.46%, 26.28% and 51.10%, respectively. Soaking the seeds with different concentrations of proline at room temperature can improve the germination potential, germination rate, germination index and vigor index of the two varieties of maize seeds. Under the conditions of low temperature, with the gradual increase in proline concentration, the germination potential, germination rate, germination index and vigor index of the two varieties of maize seeds increased first and then decreased and reached the highest value under the P3 treatment (5 mmol·L−1). Among them, the germination potential, germination rate, germination index and vigor index of the DM 3307 increased by 55.85% and 25.23%, respectively, compared with the low temperature treatment of 50.67% and 103.68%, respectively. The germination potential, germination rate, germination index and vigor index of the XX 2 increased by 20.12%, 11.84%, 25.25% and 71.62%, respectively, compared with the low temperature treatment. It can be seen that an appropriate concentration of proline treatment can significantly improve the growth state of maize seeds, and each index of the low temperature sensitive maize variety DM 3307 is more sensitive to low temperature. At the same time, the interaction between the varieties and treatments had a very significant impact on the germination potential, germination rate, germination index and vigor index (Table 1).

Table 1.
Effects of soaking seeds with different concentrations of proline solution on maize germination energy, germination rate, germination index, vitality index and the relative value of four indicators above under low temperature stress.
3.2. Length of Coleoptile and Radicle, Weight of Young Buds and α-Amylase Activity
It can be seen from Table 2 that compared with the normal temperature control treatment, the low temperature treatment significantly inhibited the coleoptile length, radicle length, fresh weight and dry weight of young buds of the DM 3307 and XX 2 seeds, and the decrease in the low temperature sensitive maize variety DM 3307 was more significant than that of XX 2. Under the normal temperature treatment, with the gradual increase in proline concentration, the germination indexes of the two varieties of maize were higher than those under the normal temperature treatment. Under low temperature conditions, with the gradual increase in proline concentration, the coleoptile length, radicle length, fresh bud weight, and dry bud weight of the two varieties of maize seeds showed a single peak trend and reached the extreme value under the P3 treatment (15 mmol·L−1). Compared with the low temperature treatment, the radicle length of the P1, P2, P3 and P4 treatment of the DM 3307 increased by 26.37%, 38.46%, 48.35% and 46.15%, respectively; the XX 2 increased by 11.11%, 24.79%, 31.62% and 24.79%, respectively. The dry weight of buds treated with P1, P2, P3 and P4 of the DM 3307 increased by 43.75%, 56.25%, 66.67% and 61.11%, respectively; the XX 2 increased by 23.68%, 30.53%, 35.26 and 31.58%, respectively. Meanwhile, under the treatment of P1, P2, P3 and P4, α-Amylase activities of the DM 3307increased by 15.80%, 30.16%, 57.62% and 36.14%, respectively; the XX 2 increased by 2.17%, 15.95%, 26.86% and 19.51%, respectively. It can be seen that the application of 15 mmol·L−1 proline can significantly promote the growth of maize buds, and the parameters of the DM 3307 are more sensitive to low temperature. At the same time, the interaction between the varieties and treatments had a very significant effect on coleoptile length, radicle length, fresh weight and dry weight of young buds. Low temperature treatment significantly inhibited the α-amylase activity of the DM 3307 and XX 2. Under the normal temperature treatment, with the gradual increase in seed soaking concentration of proline, the α-amylase activity was higher than that of the normal temperature treatment in varying degrees. At low temperature, with the gradual increase in proline concentration, the α-Amylase activity showed a single peak trend and reached the extreme value under the P3 treatment (15 mmol·L−1). Compared with the low temperature treatment, the radicle length of the P1, P2, P3 and P4 treatment of the DM 3307 increased by 15.80%, 30.16%, 57.62% and 36.14%, respectively; the XX 2 increased by 2.17%, 15.95%, 26.86% and 19.51%, respectively (Table 2). Combined with the experimental results of Table 1, our experiment shows that the experimental design has a certain accuracy and representativeness, can truly reflect the differences between the different concentrations, and can further study the effect of proline on maize embryo under normal and low temperature conditions (Table 1 and Table 2).
Table 2.
Effects of soaking seeds with different concentrations of proline solution on the length of coleoptile and radicle, fresh and dry weight of young buds, α-amylase activity and the relative value of the four indicators above under low temperature stress.
3.3. Proline Contents and Activities of P5CS, OAT and ProDH
It can be seen from Figure 1 that with the extension of low temperature stress time, the proline content, P5CS activity and OAT activity in the DM 3307 and XX 2 embryos gradually increased, while the ProDH activity showed a gradual decreasing trend and reached the extreme value on the fifth day of treatment. On the fifth day of treatment, compared with the normal temperature control treatment, the low temperature stress treatment significantly increased the proline content, P5CS activity and OAT activity of the two maize varieties, but inhibited the ProDH activity. Under the condition of low temperature stress, the proline soaking treatment further promoted the proline content, P5CS activity and OAT activity of the two maize varieties. The decrease in ProDH activity was alleviated, but the change was not significant and this promoting effect was more significant in the DM 3307. On the fifth day of low temperature stress, compared with the low temperature stress treatment, the proline content, P5CS activity, OAT activity and ProDH activity in the DM 3307 embryos increased by 48.33%, 62.13%, 14.90% and 23.94%, respectively; the proline content, P5CS activity, OAT activity and ProDH activity in the XX 2 embryos increased by 27.63%, 38.35%, 9.38% and 19.87%, respectively. At the same time, after the maize embryo was relieved of low temperature stress, the proline content, P5CS activity and OAT activity of the two maize varieties decreased, while the ProDH activity increased, and the promoting effect of the proline soaking was still obvious (Figure 1).
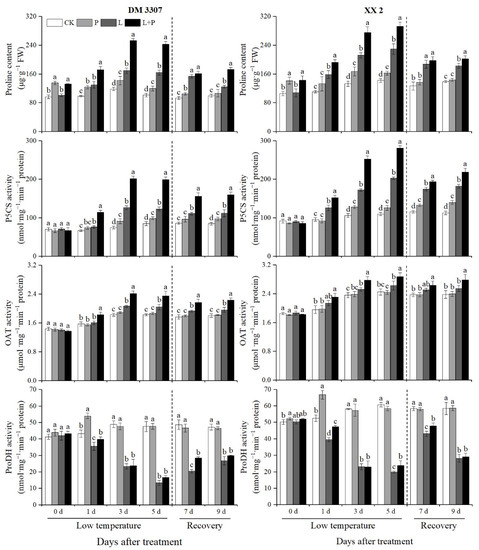
Figure 1.
Effect of exogenous proline on the proline contents and activities of P5CS, OAT and ProDH in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
3.4. Vt, ρValt and ρVcyt
As can be seen from Figure 2, the Vt and ρValt increased rapidly in the early stage of low temperature stress (day 1). With the extension of the low temperature stress time, the two indexes continued to decline and recovered after restoring normal temperature. Compared with the control, the Vt and ρValt of the DM 3307 increased by 18.18% and 56.42%, respectively, on the first day and decreased by 64.33% and 59.70%, respectively, on the fifth day; the Vt and ρValt of the XX 2 increased by 35.98% and 95.6%, respectively, on the first day, and decreased by 41.34% and 39.58%, respectively, on the fifth day.
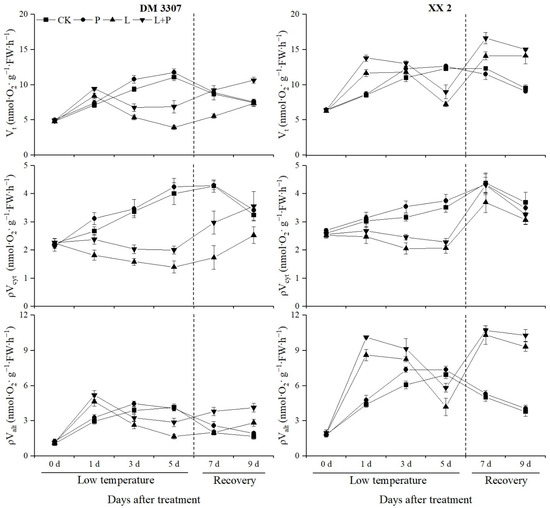
Figure 2.
Effect of exogenous proline on the Vt, ρValt and ρVcyt ratio of embryos of maize under low temperature stress.
However, the application of exogenous proline alleviated the decrease in the Vt and ρValt caused by low temperature to a certain extent. Compared with the low temperature treatment, the Vt and ρValt of the DM 3307 embryos treated with proline soaking under low temperature stress increased by 74.79% and 72.56% on the fifth day, and 44.49% and 45.54% on the fourth day of recovery, respectively; the Vt and ρValt of the XX 2 embryos increased by 24.21% and 38.87%, respectively, on the fifth day, and 6.56% and 10.29%, respectively, on the fourth day. During low temperatures, the ρVcyt continued to decline and recovered after the low temperature was relieved. Compared with the control, the ρVcyt of the DM 3307 and XX 2 after the low temperature treatment decreased by 65.00% and 41.14% on day 5, respectively. Exogenous proline partially alleviated the ρVcyt decrease caused by low temperature and recovered significantly after the release of low temperature. Compared with the low temperature treatment, the ρVcyt of the DM 3307 and XX 2 were increased by 43.03% and 10.25%, respectively, on the fifth day, and 40.65% and 6.07%, respectively, on the fourth day. The results showed that the exogenous proline treatment could reduce the damage of the electron transfer pathway of a seed embryo under low temperature stress and provide energy for seed germination. Under normal temperature, the Vt, ρValt and ρVcyt trend of the DM 3307 and XX 2 was similar, and both of them showed the trend of increasing first and then decreasing slowly. Exogenous proline treatment can improve the Vt, ρValt and ρVcyt of the maize embryos of the two varieties. The Vt and ρValt of the DM 3307 and XX 2 reached a significant level on day 3, the ρValt of the DM 3307 reached a significant level on day 7 and EEE on day 1, and the ρVcyt of the XX 2 embryos reached a significant level on day 3 (Figure 2).
3.5. Activities of COX and AOX
As the key enzyme of the electron transport chain, COX is the key to maintaining mitochondrial function and cell energy metabolism. Figure 3 shows that under low temperature stress, the activity of COX in maize embryos of the two varieties showed a downward trend and recovered rapidly after restoring normal temperature. The decreasing rate of COX activity in the DM 3307 was higher than that in the XX 2. Compared with the control, the COX activity of DM 3307 and XX 2 decreased by 65.18% and 45.35%, respectively, on the fifth day. Under low temperature stress, exogenous proline alleviated the decrease in COX activity of the seed embryos caused by low temperature. After restoring normal temperature, the COX activity of the seed embryos recovered faster, and had a stronger effect on the decrease in COX activity of the DM 3307 embryos under low temperature stress. Compared with the low temperature treatment, the COX activity of the DM 3307 and XX 2 embryos treated by proline soaking under low temperature stress increased by 53.83% and 27.06% on the fifth day, and 63.46% and 13.74% on the second day of recovery, respectively. Under normal temperature, the change trend of COX activity in the DM 3307 and XX 2 embryos was similar, both of which increased first and decreased slowly. Exogenous proline treatment could improve the COX activity of the maize embryo of the two varieties, but it did not reach a significant level.
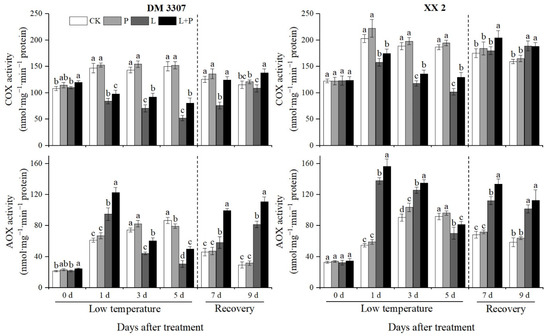
Figure 3.
Effect of exogenous proline on the activities of COX and AOX in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
AOX is the terminal oxidase of the alternative respiration pathway. The AOX electron transport pathway plays an important role in seed germination. Figure 3 shows that under the low temperature stress treatment, the activity of the AOX in maize embryos of the two varieties increased first and then decreased and then recovered after restoring normal temperature. The increase in the AOX activity in the XX 2 was higher than that in the DM 3307. Compared with the control, the activity of AOX in the DM 3307 and XX 2 embryos increased by 55.17% and 152.21%, respectively, on the first day, and decreased by 64.70% and 23.64%, respectively, on the fifth day. The application of exogenous proline under low temperature stress alleviated the decrease in AOX activity of the seed embryos caused by low temperature to a certain extent. After restoring normal temperature, the AOX activity of the seed embryos recovered faster, and the decrease in AOX activity of the DM 3307 seed embryos under low temperature stress was alleviated more significantly. Compared with the low temperature treatment, the AOX activity of the DM 3307 and XX 2 embryos increased by 62.34% and 16.00%, respectively, on the fifth day and 35.81% and 10.76%, respectively, on the fourth day of recovery. Under normal temperature, the exogenous proline treatment significantly increased the activity of AOX in the maize embryo of the two varieties on the fifth day (Figure 3).
3.6. Activities of PFK, HXK and PK
Hexokinase (HXK), phosphofructokinase (PFK) and pyruvate kinase (PK) catalyze three irreversible reactions in the EMP pathway. They are important regulatory enzymes in the EMP pathway and provide energy for the process of seed germination. During the period of low temperature stress, the changes of PFK activity in the embryos of the DM 3307 and XX 2 were different. The PFK activity of the embryos of DM 3307 showed a downward trend and XX 2 showed an upward trend, but the PFK activity of the two varieties was significantly lower than that of the control. Compared with the control, the activity of PFK in the DM 3307 and XX 2 embryos decreased by 71.27% and 33.53% on the fifth day, respectively. The application of exogenous proline under low temperature stress alleviated the decline of PFK activity of the seed embryos caused by low temperature to a certain extent. The PFK activity of the seed embryos recovered faster after restoring normal temperature, and the remission of PFK activity of the DM 3307 embryos under low temperature stress was more significant. Compared with the low temperature treatment, the PFK activity of the DM 3307 and XX 2 embryos in the proline soaking treatment under low temperature stress increased by 114.60% and 20.03% on the fifth day, and 58.48% and 1.74% on the fourth day of recovery, respectively (Figure 4).
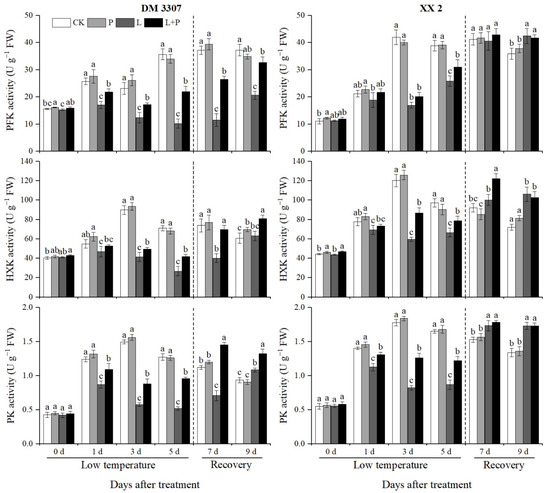
Figure 4.
Effect of exogenous proline on the activities of PFK, HXK and PK in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
Under low temperature stress, the activities of HXK and PK in the maize embryos of the two varieties increased first and then decreased. The activity of HXK and PK in the DM 3307 decreased more than that in the XX 2. Compared with the control, the activities of HXK and PK in the DM 3307 embryos reached the peak on the fifth day, and decreased by 62.30% and 59.20%, respectively; the activities of HXK and PK in the XX 2 embryos reached the peak on the third day, decreased by 50.50% and 53.63%, respectively, and increased slightly on the fifth day. Under low temperature stress, the application of exogenous proline alleviated the decline of HXK and PK activities of the seed embryos caused by low temperature to a certain extent. The HXK activity of the seed embryos recovered rapidly after restoring normal temperature. Compared with the low temperature treatment, the HXK activity of the DM 3307 and XX 2 embryos increased by 56.00% and 17.99%, respectively, on the fifth day, and 73.94% and 21.69%, respectively, on the second day of recovery; the PK activity increased by 84.13% and 40.81% on day 5 and 105.45% and 2.70% on day 2, respectively, (Figure 4). Compared with the control, the activity of PK in the DM 3307 embryos after the low temperature treatment reached the peak on the fifth day and decreased by 62.30%. The activity of PK in the XX 2 PK embryo reached the peak on the third day, decreased by 50.50%, and increased slightly on the fifth day. The application of exogenous proline under low temperature stress alleviated the decrease in PK activity of the seed embryos caused by low temperature to a certain extent, and the PK activity of the seed embryos recovered faster after restoring normal temperature. Compared with the low temperature treatment, the PK activity of the DM 3307 and XX 2 embryos increased by 56.00% and 17.99%, respectively, on the fifth day, and 73.94% and 21.69%, respectively, on the second day of recovery. Compared with the low temperature treatment, the effect of exogenous proline on the activities of PFK, HXK and PK of the XX 2 embryos after stress relief did not reach a significant level, but the activities of PFK, HXK and PK of the DM 3307 embryos increased significantly, indicating that proline had a more significant effect on low temperature sensitive varieties (Figure 4).
3.7. Activities of IDH, SDH and MDH
It can be seen from Figure 5 that the low temperature stress treatment significantly reduced the activity of IDH in the embryos of the two varieties of maize, especially in the DM 3307 embryos. After restoring normal temperature, the activity of IDH in the embryos of the two varieties of maize partially recovered. Compared with the control, the activity of IDH in embryos of the DM 3307 treated with low temperature showed a downward trend, reached the peak on the fifth day, and decreased by 65.34%; the activity of IDH in the XX 2 embryos decreased first and then increased, reached the peak on the first day, and decreased by 45.97%. The application of exogenous proline under low temperature stress alleviated the decrease in seed embryo IDH activity caused by low temperature to a certain extent. After the stress was relieved, the seed embryo IDH activity recovered faster, and the remission of the DM 3307 seed embryo IDH activity under low temperature stress was more significant. Compared with the low temperature treatment, the IDH activity of the DM 3307 and XX 2 embryos treated with proline soaking under low temperature stress increased by 68.91% and 23.62%, respectively, on the fifth day, and increased by 16.53% and 5.00%, respectively, on the fourth day of recovery (Figure 5).
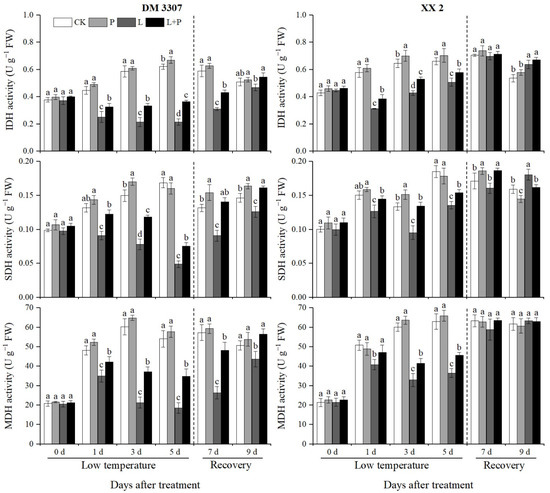
Figure 5.
Effect of exogenous proline on the activities of IDH, SDH and MDH in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
The low temperature stress treatment significantly reduced the activity of SDH in the maize embryos of the two varieties, and the activity of SDH partially recovered after restoring normal temperature. Compared with the control, the SDH activity of the DM 3307 embryos treated with low temperature showed a downward trend, reached the peak on the fifth day, and decreased by 71.09%; the SDH activity of the XX 2 embryos increased first, then decreased and then increased, reached the peak on the third day, then decreased by 28.60%. The application of exogenous proline under low temperature stress alleviated the decline of SDH activity of the seed embryos caused by low temperature to a certain extent. After the stress was relieved, the SDH activity of the seed embryos recovered faster, and the SDH activity of the DM 3307 embryos under low temperature stress was alleviated more significantly. Compared with the low temperature treatment, the SDH activity of the DM 3307 and XX 2 embryos increased by 54.72% and 13.41%, respectively, on the fifth day and 54.58% and 16.08%, respectively, on the second day of recovery. Under the low temperature stress treatment, the activity of MDH in the maize embryos of the two varieties increased first, then decreased, and recovered after the treatment was removed. Compared with the control, the MDH activity of the DM 3307 embryos reached the peak on the fifth day of the low temperature treatment; the activity of MDH in the XX 2 embryos reached the peak on the third day of the low temperature treatment. The application of exogenous proline under low temperature stress alleviated the decrease in MDH activity of the seed embryos caused by low temperature to a certain extent. After the stress was relieved, the MDH activity of the seed embryos recovered faster, and the MDH activity of the DM 3307 embryos under low temperature stress was alleviated more significantly. Compared with the low temperature treatment, the MDH activity of the DM 3307 and XX 2 embryos treated with proline soaking under low temperature stress increased by 86.76% and 25.22% on the fifth day, and 82.99% and 7.72% on the second day of recovery, respectively. Compared with the low temperature treatment, the application of exogenous proline had no significant effect on the activities of IDH and MDH of the XX 2 embryos after stress relief, but the activities of IDH and MDH of the DM 3307 embryo reached a significant level. This shows that SDH activity is greatly affected by low temperature, and the mitigation effect of proline on low temperature sensitive varieties is more significant (Figure 5).
3.8. Activities of G-6-PDH and 6-P-GDH
The key metabolic pathway of seed germination is the smooth progress of the pentose phosphate pathway (PPP). G-6-PDH and 6-P-GDH are two key enzymes in the pentose phosphate pathway. It can be seen from Figure 6 that during low temperature, the activities of the G-6-PDH and 6-P-GDH combined enzymes in the maize embryos of the two varieties first increased and then decreased, and then recovered after the stress was relieved. Compared with the control, the activities of the G-6-PDH and 6-P-GDH combined enzymes in the embryos of the DM 3307 treated with low temperature reached the peak on the fifth day, which decreased by 49.11%; the activities of the G-6-PDH and 6-P-GDH combined enzymes in the XX 2 embryos reached the peak on the third day and decreased by 37.98%. Under low temperature stress, the application of exogenous proline alleviated the decrease in G-6-PDH and 6-P-GDH combined enzyme activities of the seed embryos caused by low temperature to a certain extent. After the stress was relieved, the G-6-PDH and 6-P-GDH combined enzyme activities of the seed embryos recovered faster, and the G-6-PDH and 6-P-GDH combined enzyme activities of the DM 3307 embryos were alleviated more significantly. Compared with the low temperature treatment, the combined enzyme activities of the G-6-PDH and 6-P-GDH of the DM 3307 and XX 2 embryos treated by proline soaking under low temperature stress, increased by 41.96% and 11.63%, respectively, on the fifth day, and increased by 32.10% and 5.51%, respectively, on the second day of recovery (Figure 6).
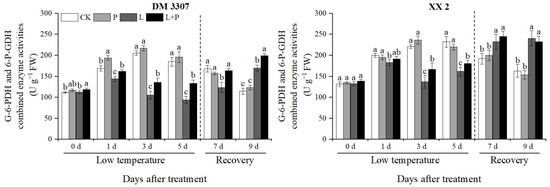
Figure 6.
Effect of exogenous proline on the activities of G-6-PDH and 6-P-GDH in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
3.9. ATP Contents
During the low temperature period, the ATP content in the embryos of the two varieties of maize first increased and then decreased, and then recovered after the stress was relieved, as shown in Figure 7. Compared with the control, the ATP content of the DM 3307 and XX 2 embryos reached the peak on the third day, which decreased by 86.94% and 56.79%, respectively; however, the application of exogenous proline under low temperature stress alleviated the decrease in ATP content and activity of the seed embryos caused by low temperature to a certain extent. After the stress was relieved, the ATP content and activity of the seed embryos recovered rapidly, and the ATP content of the DM 3307 embryos increased significantly. Compared with low temperature treatment, the ATP content and activity of the DM 3307 and XX 2 embryos were increased by 194.51% and 37.19%, respectively, on the fifth day and 236.45% and 0.28%, respectively, on the fourth day of recovery (Figure 7).
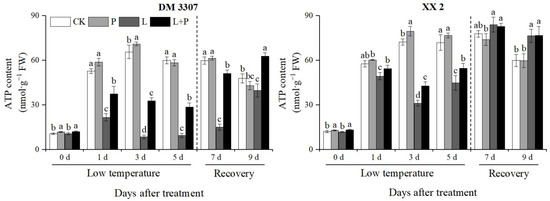
Figure 7.
Effect of exogenous proline on the ATP contents of embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
3.10. Contents of ABA, IAA, GA, ZR and GA/ABA Ratio
It can be seen from Figure 8 that on the third and fifth days of treatment, the low temperature stress treatment significantly increased the ABA content of the seed embryos of the two maize varieties, and proline inhibited the ABA content of the seed embryos of the two maize varieties under the normal temperature and low temperature treatments after seed soaking. The ow temperature stress treatment significantly inhibited the IAA content of DM 3307 and XX 2 embryos, and exogenous proline significantly increased the IAA content of all kinds of embryos of the two products, which was the most significant on the third and fifth days of low temperature. The IAA content of the DM 3307 increased by 90.04% and 61.79%, respectively; the IAA content of the XX 2 increased by 26.15% and 17.52%, respectively. It can be seen that the IAA content of the low temperature sensitive maize varieties was more sensitive to low temperature. Similarly, the contents of GA and ZR in the embryos of the two maize varieties increased first and then decreased with the extension of the low temperature stress time and reached the peak on the third day of treatment. Compared with the normal treatment, the low temperature treatment significantly inhibited the contents of GA and ZR in the embryos of the two varieties. On the third day of the low temperature treatment, compared with the simple low temperature treatment, the GA and ZR contents of the DM 3307 embryos increased by 34.01% and 14.98%, respectively the GA and ZR contents of the XX 2 embryos increased by 18.18% and 14.65%, respectively. At the same time, the low temperature stress treatment also significantly inhibited the GA/ABA ratio. On the fifth day of the low temperature treatment, the GA/ABA ratio of the DM 3307 and XX 2 embryos decreased by 44.69% and 39.94%, respectively. Seed soaking with proline could significantly increase the GA/ABA ratio in the embryo of the two maize varieties. After relieving the low temperature stress of the maize embryos, the contents of IAA, GA and ZR and the ratio of GA/ABA in the DM 3307 and XX 2 embryos increased in varying degrees, while the content of ABA decreased, and the exogenous proline treatment could also promote the accumulation of IAA, GA and ZR contents in embryos (Figure 8).
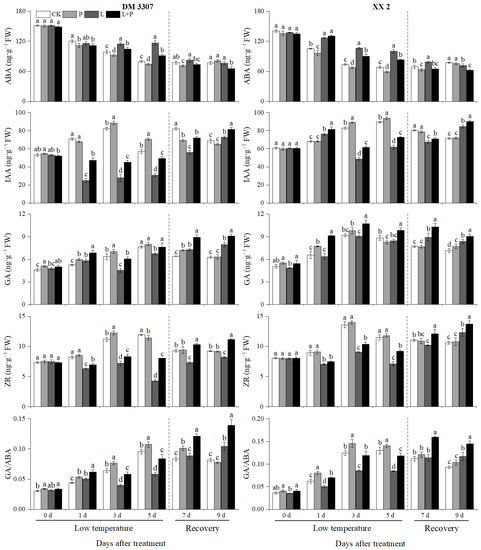
Figure 8.
Effect of exogenous proline on the contents of ABA, IAA, GA, ZR and GA/ABA ratio in embryos of maize under low temperature stress. Different letters within a column indicate significant differences at p < 0.05.
4. Discussion
Seed germination is the most vigorous and complex stage of physiological metabolism in the plant life cycle. The essence of the germination process is the process that the seed embryo recovers from a dormant state to an active growth state, that is, after the seed is imbibed, the seed embryo cells begin to elongate, the radicle breaks through the seed coat and enters the germination state, the cell division and elongation of the embryo accelerate, and the radicle and germ grow rapidly and enter the germination stage. The whole process of seed germination is particularly sensitive to the external environment and will be restricted by many factors such as low temperature, drought and hypoxia [81,82,83]. The quality of seed germination determines the strength of plant seedlings, and the duration of germination determines whether the seedlings can enter the growth environment corresponding to their development at the optimal period, which has a very important impact on the yield [84,85]. A low temperature will not only slow down the repair of the seed cell biofilm, but also cause membrane lipid phase change and membrane structure damage of the repaired membrane, resulting in solute extravasation, respiratory metabolism disorder, enzyme activity weakening, material transformation becoming blocked, toxic substance accumulation, and finally slow or even no germination. In this study, under low temperature stress, the germination rate, germination potential, germination index, vigor index, coleoptile length, radicle length, dry and fresh weight of two varieties of maize seeds decreased significantly. Our results are consistent with the research results of Chang Bowen [86] and Zheng Yunye [87]. At the same time, we found that the DM 3307 suffered more stress than the XX 2, indicating that the XX 2 had a stronger resistance to low temperature stress than the DM 3307. The regulation of exogenous substances on crop growth and development is one of the effective measures to resist adversity. Proline is the most effective compatible osmotic regulator in plants, which is produced in large quantities when plants are subjected to stress. The study on the regulation effect of proline on crops under different stresses shows that exogenous proline can effectively alleviate the inhibition of stress on plant growth and the inhibition of low temperature stress on seed germination [88]. Soaking seeds with different concentrations of proline can effectively alleviate the inhibition of low temperature stress on quinoa seed germination, and 20 mmol·L−1 proline can significantly improve the germination rate and root length of quinoa seeds under low temperature stress [89]. Proline increased the seed germination potential of Arabidopsis under low temperature stress and in addition to alleviating the inhibition of low temperature stress on seed germination, proline also alleviates a variety of stresses [28]. A 15 mmol·L−1 proline solution had the best promotion effect. The energy and substances needed for seed germination are completely derived from the oxidative decomposition of storage substances. The main storage substances in maize seeds are starch, protein and fat, and low temperature stress will inhibit α-amylase activity, which hinders the supply of materials and energy required for embryo growth, thus inhibiting seed germination. In this study, seed soaking with proline alleviated the inhibitory effect of low temperature on the α-amylase activity to a certain extent, thus restoring the supply of materials and energy required for seed embryo growth and promoting seed germination.
Proline is considered to be the most widely distributed and important osmotic regulator [90,91]. Studies have shown that the contents of osmotic regulators such as proline, soluble sugar and soluble protein in plants will be greatly increased after low temperature stress, so as to coordinate the osmotic pressure inside and outside cells, protect membrane stability and maintain ion balance. In this study, we found that under low temperature stress, the endogenous proline in a maize embryo increased significantly, the internal metabolism of the maize embryo was restored, and the respiration was strengthened. The results of this study are consistent with previous research results in cucumber, banana, quinoa and chrysanthemum. Exogenous proline can increase the content of endogenous proline, soluble sugar and soluble protein in a seed embryo. The increase in osmoregulation substances protect the structure of the cell membrane and protein, reduces membrane damage and enhances a plant’s cold resistance [92]. After the stress is relieved, the proline accumulated by seed embryos soaked with proline decreases rapidly and then tends to be stable. A large amount of proline can be quickly degraded as an energy substance after the stress is relieved, providing more carbon and nitrogen sources and energy for seed germination, which is also one of the important reasons for the rapid germination of seeds treated with exogenous proline after recovery.
Proline synthesis mainly has two pathways, the Glu pathway and the Orn pathway, which are regulated by P5CS and OAT, respectively; The degradation of proline is mainly regulated by ProDH [93]. Proline accumulation during seed germination mainly depends on the Orn pathway. Antisense inhibition of AtP5CS expression will lead to delayed radicle elongation during Arabidopsis seed development [94]. Proline biosynthesis and decomposition can be associated with many metabolic pathways, among which Orn and Arg are the synthetic precursors of polyamines, which play an important role in plant development and stress resistance [95,96]. Previous studies have shown that P5CS activity is affected by a variety of abiotic stresses. Moreover, overexpression of the P5CS gene can enhance the stress resistance of plants [97]. Compared with the control, the activity of P5CS in the DM 3307 embryos began to increase significantly on the third day of low temperature stress treatment; the P5CS activity of the XX 2 embryos increased significantly from the first day. The changes of ProDH activity of the two varieties were consistent and decreased continuously with the extension of the low temperature stress time; OAT activity increased significantly on day 3. This indicates that maize embryos accumulate proline mainly by enhancing the glutamate pathway and reducing proline degradation. Varieties with strong resistance can activate the Glu pathway earlier to accumulate proline and improve cold resistance [98]. This is consistent with the results of previous studies. The P5CS gene increased rapidly in the early stage of stress, while the OAT gene began to increase after 12 h. Exogenous proline treatment further increased the activities of P5CS and OAT in the maize embryo under low temperature stress, and the activity of ProDH increased significantly only in the early stage of low temperature stress and stress relief. The results showed that exogenous proline increased the accumulation of proline significantly by increasing the activities of P5CS and OAT under low temperature stress and enhancing the two proline synthesis pathways. The enhancement of ProDH activity in the early stage of low temperature stress and stress relief may be related to the induction of proline, which also explains the reason why proline can degrade rapidly after stress relief. The increase in ProDH activity can accelerate the degradation of proline and provide carbon and nitrogen sources and a large amount of reducing power for seed embryo recovery and seed germination. Proline also has a more significant mitigation effect on low temperature sensitive varieties [99]. The above research fully shows that exogenous proline is essential for seed embryo development after the release of low temperature stress.
Respiration is the main source of energy supply during seed germination and the core of material metabolism [100]. According to the different terminal oxidase, the electron transport pathway of plant mitochondria can be divided into the cytochrome pathway (CP) and the alternative pathway (AP), which are catalyzed by cytochrome oxidase (COX) and alternative oxidase (AOX), respectively. COX is the main terminal oxidase during plant growth, whereas CP is the main synthesis pathway of ATP, and a low temperature will reduce COX activity. Meanwhile the regulation of AOX activity is very complex. The energy produced by AP is mainly lost by heat, and the proportion of AP will gradually increase during seed germination. Studies have confirmed that the AOX gene will be expressed in response to stress, especially low temperature stress. AP is activated, the maximum capacity of the alternative pathway (Valt) is increased, and the adaptability of plants to low temperature is strengthened [101,102], but more evidence showed that ρValt is different from Valt, and ρValt can truly reflect the state and function of plants under specific physiological conditions [103]. This study showed that low temperature stress reduced the activity of the cytochrome main pathway operation (ρVcyt) and COX. After the stress was relieved, the XX 2 recovered significantly, and the DM 3307 recovered slowly, indicating that CP was blocked and the mitochondrial function of the seed embryo cells was damaged under low temperature stress, especially the DM 3307, which is not resistant to low temperatures. The activity of the Vt, ρValt and AOX increased rapidly on the first day, which was higher than that of normal temperature control; on the third day, the Vt and ρValt of the DM 3307 embryos decreased significantly, which was lower than that of the control; Vt and ρValt of the XX 2 decreased rapidly on the fifth day but recovered after the stress was relieved. Low temperature stress can lead to the decrease in COX, the obstruction of CP and the destruction of the respiratory electron transport chain. As an “overflow” pathway, the strengthening of the AP can not only accelerate the consumption of electrons and reduce the production of reactive oxygen species, but also ensure the operation of the respiratory metabolic pathway and the most basic demand for respiratory metabolism to maintain the life activities of seed embryos under low temperature stress [104]. The decline of ρValt in the later stage of low temperature stress may be related to the integrity of the mitochondrial membrane system [105]; however, proline treatment increased the activities of AOX and COX under low temperature stress and alleviated the decline of Vt, ρValt and ρVcyt. This may be because proline treatment increases the inner proline metabolism. Proline degradation provides a reduction equivalent for electron transport in the respiratory chain and ATP production, reduces the content of reactive oxygen species, protects the activities of the AOX and COX enzymes, alleviates the damage of membrane lipid peroxidation, maintains the integrity of the mitochondrial membrane, accelerates the repair and resynthesis of mitochondria and the recovery of seed embryo cells after stress relief, and promotes the repair of stress-induced injury [106,107].
There are many respiratory and metabolic pathways in plants, including the EMP, TCA and PPP. The EMP pathway is the first stage of respiratory metabolism, which can oxidize and decompose carbohydrates into pyruvate and release ATP. Three steps in the EMP pathway are irreversible reactions, involving three key enzymes, namely, hexokinase (HXK), phosphofructokinase (PFK) and pyruvate kinase (PK) [108]. In the early stage of seed germination, the EMP pathway will be activated rapidly and the activity of metabolic enzymes will increase rapidly. In this study, the activity changes of PFK in the DM 3307 and XX 2 embryos were different during low temperature stress. PFK in the DM 3307 embryos increased first and then decreased, and the XX 2 showed an upward trend. The activities of HXK and PK in the two varieties increased first and then decreased, but the activities of PFK, HXK and PK in the two varieties were significantly lower than those in the control after low temperature stress. This is because under low temperature stress, the seed’s own stress protection system will reduce the activities of PFK, HXK and PK, slow down the oxidative decomposition of sugar, and the accumulated soluble sugar will participate in cell osmotic regulation to improve its adaptability to low temperature. This is consistent with the results of previous studies on different POA varieties under low temperature stress [109]. Compared with the low temperature stress treatment, the proline treatment significantly increased the activities of PFK, HXK and PK under low temperature stress, especially the increase in PFK and PK. This shows that exogenous proline alleviated the cell osmotic stress, transformed the sugar accumulated under low temperature stress into the reaction substrate of subsequent material energy metabolism by improving the activity of PK, promoted the recovery of PFK activity, accelerated the operation of the EMP pathway, produced more ATP, enhanced the respiration of the maize embryo under low temperature stress, and alleviated the inhibition of low temperature on seed germination. The proline treatment increased the activities of PFK, HXK and PK under low temperature stress, which may also be because the proline accelerated the repair and resynthesis of mitochondria and promoted the TCA cycle; however, there are few studies on the regulation of exogenous proline on the plant EMP pathway under low temperature stress. Further experiments are needed to verify the regulation mechanism of proline on glycolysis.
The TCA cycle is the central pathway for the catabolism and transformation of sugar, fat and protein. It can not only provide significant of energy for plant life activities, but also serve as the carbon skeleton of the biological oxidation matrix. IDH, SDH and MDH are key enzymes in the TCA cycle and begin to be expressed in the early stage of seed germination [110]. MDH not only catalyzes the reversible transformation between malic acid and oxaloacetic acid, but also confirms that MDH is closely related to plant seed germination, sugar accumulation, fruit development and maturation, stress resistance and other physiological processes [111,112]. Previous studies have shown that stress induced high expression of the millet MDH gene can improve plant stress resistance, and the MDH gene plays an important role in the stress response signal system [113]. In this study, the activities of IDH, SDH and MDH of the two varieties under low temperature stress were significantly lower than those of the control, and the TCA cycle rate decreased as a whole, but the change trend was different. IDH was the first and most affected by low temperature stress in the DM 3307 embryos. Compared with day 0, IDH decreased significantly, SDH did not change significantly, and MDH increased significantly; however, with the extension of the stress time, the activities of the three enzymes decreased gradually. The activity of IDH in the XX 2 embryos decreased first and then increased with the extension of stress time, while the activities of SDH and MDH fluctuated first and increased. After the stress was relieved, the activities of IDH, SDH and MDH of the XX 2 recovered rapidly, which was basically consistent with the control, but the recovery of the DM 3307 was slow. The response of the TCA cycle to low temperature stress was significantly different among the different genotypes, and the varieties with strong resistance were less affected. The exogenous proline treatment significantly increased the activities of IDH, SDH and MDH in the maize embryos under low temperature stress and had the strongest effect on the activity of MDH in the DM 3307 embryos. This shows that proline can effectively promote the transformation rate of the TCA pathway, accelerate the decomposition of substrate, promote the progress of aerobic respiration under low temperature stress, and promote seed germination while improving seed stress resistance. The site of aerobic oxidation in the TCA cycle is in the mitochondria. The promoting effect of proline on the TCA cycle is also related to its protection of mitochondrial structural integrity and improvement of electron transport chain efficiency under low temperature stress [114]. In addition, the process of proline degrading into α-ketoglutarate in the mitochondria may provide a carbon source for the TCA cycle and electrons for the mitochondrial electron transport chain to produce ATP. The effect of proline on alleviating the damage of plants caused by low temperature stress is particularly prominent after the stress is relieved [115].
PPP can directly lead to oxidative decarboxylation and molecular recombination of glucose across the EMP-TCA cycle and release a large amount of reducing force. The whole reaction process uses NADP+ as the coenzyme of dehydrogenase to produce a large amount of NADPH, which does not involve the production and consumption of ATP. The intermediate products of PPP can participate in the synthesis of substances related to plant growth and stress resistance, such as nucleic acids, auxin, amino acids and phenols, and can also be connected with photosynthesis to realize the transformation between monosaccharides. PPP is not only closely related to the release of seed dormancy but is also a key pathway of respiratory metabolism during seed germination [116]. PPP also plays an important role in plants under stress. G-6-PDH and 6-P-GDH are key rate limiting enzymes in PPP. In this study, compared with that before stress, the activities of the G-6-PDH and 6-P-GDH combined enzymes in embryos of DM 3307 increased first and then decreased with the extension of low temperature stress, while the XX 2 fluctuated slightly and increased, but all treatments were significantly lower than normal temperature. Proline metabolism is related to PPP. NADPH generated by PPP is used to synthesize proline and produce NADP+, which supplements the electron receptor required for electron transfer. Compared with the low temperature stress treatment, the proline treatment significantly increased the combined enzyme activities of G-6-PDH and 6-PGDH in embryos from the first day, which is consistent with the results of previous studies. The above results show that the proline treatment alleviated the decline of PPP operation efficiency in the seed embryos due to the restriction of the G-6-PDH and 6-PGDH combined enzyme activity under low temperature and supplemented the NADP+ library through proline anabolism to improve PPP efficiency. The intermediate products generated by the operation of the PPP pathway can be used for the repair of seed embryo cell DNA, provide reducing power for seed embryo cell biosynthesis, promote the production of new ATP, maintain the material and energy metabolism demand of a seed embryo under low temperature stress, maintain the reducing environment in cells, reduce the damage of reactive oxygen species to proteins and membrane lipids, and further reduce the damage of low temperature stress to seed embryo cells. Exogenous proline treatment after stress relief can rapidly improve the activities of G-6-PDH and 6-PGDH. It may be that a large amount of proline decomposes after restoring normal temperature, provides electrons for the respiratory chain, and can further activate the PPP pathway. The decomposed products participate in material metabolism as a carbon and nitrogen source, accelerate the elongation of seed cells after low temperature stress relief, and promote seed germination [117]. Our experiment only discussed the effect of proline on aerobic respiration, and the research direction will focus on anaerobic respiration in the future.
A plant hormone is an organic substance that can regulate growth and development, which is metabolized and synthesized by plants under the induction of certain signals and the process of seed germination is regulated by a variety of hormones, rather than the result of a single hormone [118]. In this study, low temperature stress significantly increased the concentration of ABA in the embryo of two maize varieties and significantly decreased the concentration of IAA, GA and ZR. Additionally, low temperature stress significantly reduced the GA/ABA ratio of the two varieties. GA/ABA of the DM 3307 showed a fluctuating trend, which was an obvious imbalance of plant hormones, therefore, its germination was inhibited more obviously. At the same time, GA/ABA of the XX 2 gradually increased, showing a good adaptability and resistance to low temperature stress. The results of this study are basically consistent with those of previous studies [119]. ABA accumulation can significantly inhibit seed germination, but it is also an important intercellular signal, which can conduct signal transduction rapidly under stress conditions [120]. ABA and proline have an obvious interaction in determining stress tolerance [121]. During abiotic stress, ABA induced the expression of P5CS and P5 CR, but could not regulate the expression of ProDH [122]. In this study, exogenous proline treatment significantly reduced the concentration of ABA in the embryos under low temperature stress. Exogenous proline cannot promote the production of ABA under low temperature stress, but this does not mean that there is no crosstalk between the proline and ABA signals. The decrease in ABA concentration in this study may be due to the fact that exogenous proline improves the antioxidant capacity of embryos under low temperature stress, reduces the damage of the membrane system, alleviates the disorder of endogenous hormone metabolism, restores seed germination, and carries out ABA catabolism with germination. GA can improve the germination of barley, maize and other seeds under stress conditions [123], while GA and ABA are two antagonistic hormones regulating seed germination and dormancy. The ratio of GA/ABA determines whether the seed is dormant or germinated. Therefore, GA/ABA can be used to reflect the process of seed germination and reflect its resistance and adaptability to low temperature. Exogenous proline increased the concentration of GA and the ratio of GA/ABA in embryos under low temperature stress, alleviated the inhibitory effect of low temperature on embryo germination, and accelerated the process of seed germination after relieving low temperature, especially for DM 3307, which is not resistant to low temperature. In this study, the application of exogenous proline increased the contents of IAA and ZR in maize embryos under low temperature and after stress relief. This may be related to the proline protecting the IAA and other biological macromolecules from decomposition. It also explains the reason why the proline accelerated seed embryo germination after stress relief. Our results show that the application of proline can alleviate the inhibition of low temperature on seed embryo germination by increasing the accumulation of ZR, IAA and GA in maize embryos and reducing the concentration of ABA, and by promoting seed germination after the release of low temperature stress; however, the mechanism of proline regulating endogenous hormones in seed embryos under low temperature stress needs to be further studied in the future.
5. Conclusions
Proline significantly improved the germination potential, germination rate, germination index, vigor index and α-amylase activity of maize seeds under low temperature stress, and effectively alleviated the inhibitory effect of low temperature stress. In this study, the optimum concentration of proline soaking to promote maize seed germination under low temperature stress was 15 mmol·L−1. Exogenous proline can regulate the metabolism of endogenous proline in maize embryos by regulating the activities of enzymes related to the synthesis and degradation of endogenous proline. At the same time, proline promotes the germination embryo to accumulate more endogenous proline, which not only improves the adaptability of maize seeds to low temperature stress during germination, but also provides material and an energy source for seed germination after stress is relieved. Exogenous proline can inhibit the decrease in the EMP pathway, TCA cycle and PPP key enzyme activities caused by low temperature stress, increase the activity of terminal oxidase and ATP content, effectively improve the respiratory rate during seed germination under low temperature stress, and then accelerate mitochondrial repair and resynthesis. Exogenous proline can improve the low temperature tolerance of maize embryos and alleviate the inhibitory effect of low temperature stress on maize germination by regulating the balance of endogenous plant hormones (Figure 9).
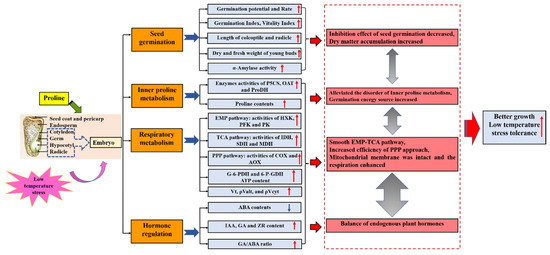
Figure 9.
Schematic representation of the positive role of exogenous proline alleviated low temperature stress by optimizing seed germination, inner proline metabolism, respiratory metabolism and hormone regulation mechanism in maize embryos. The red arrows (↑) and the blue arrows (↓) represent the positive and passive roles of spermidine, respectively.
Author Contributions
Conceptualization, J.L., W.G. and S.W.; methodology, S.Z.; software, S.Z.; validation, J.L.; formal analysis, S.Z.; investigation, S.Z.; resources, S.W.; data curation, S.Z. and J.L.; writing original draft preparation, S.Z.; writing—review and editing, S.Z. and S.W.; visualization, J.L., S.W. and W.G.; supervision, S.W; project administration, S.W. and W.G; funding acquisition, J.L. and W.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (3217150962) and the Special Project of Collaborative Innovation and Extension System of Green Organic Agriculture in Heilongjiang Province (GOAS2022).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Calicioglu, O.; Flammini, A.; Bracco, S.; Lorenzo Bellù, L.; Sims, R. The future challenges of food and agriculture: An integrated analysis of trends and solutions. Sustainability 2019, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.W. Characteristics and Reduction Potential of Losses and Waste of Major Grains of the Food Supply in China. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2018. [Google Scholar]
- Ahuja, I.; De Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Pereira, A. Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Synthesis Report; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Allen, D.J.; Ort, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Shannon, H.D.; Motha, R.P. Managing weather and climate risks to agriculture in North America, Central America and the Caribbean. Weather. Clim. Extrem. 2015, 10, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Vogel, E.; Donat, M.G.; Alexander, L.V.; Meinshausen, M.; Frieler, K. The effects of climate extremes on global agricultural yields. Environ. Res. Lett. 2019, 14, 054010. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Grzybowski, M.; Adamczyk, J.; Jończyk, M.; Sobkowiak, A.; Szczepanik, J.; Frankiewicz, K.; Fronk, J.; Sowiński, P. Increased photosensitivity at early growth as a possible mechanism of maize adaptation to cold springs. J. Exp. Bot. 2019, 70, 2887–2904. [Google Scholar] [CrossRef] [Green Version]
- Turk, H.; Erdal, S.; Dumlupinar, R. Carnitine-induced physio-biochemical and molecular alterations in maize seedlings in response to cold stress. Arch. Agron. Soil Sci. 2019, 66, 925–941. [Google Scholar] [CrossRef]
- Li, M.; Sui, N.; Lin, L.; Yang, Z.; Zhang, Y.H. Transcriptomic profiling revealed genes involved in response to cold stress in maize. Funct. Plant Biol. 2019, 46, 830–844. [Google Scholar] [CrossRef]
- Miedema, P. The effects of low temperature on Zea mays. Adv. Agron. 1982, 35, 93–128. [Google Scholar] [CrossRef]
- Trzcinska-Danielewicz, J.; Bilska, A.; Fronk, J.; Zielenkiewicz, P.; Jarochowska, E.; Roszczyk, M.; Jonczyk, M.; Axentowicz, E.; Skoneczny, M.; Sowinski, P. Global analysis of gene expression in maize leaves treated with low temperature. I. Moderate chilling (14 °C). Plant Sci. 2009, 177, 648–658. [Google Scholar] [CrossRef]
- Jończyk, M.; Sobkowiak, A.; Trzcinska-Danielewicz, J.; Skoneczny, M.; Solecka, D.; Fronk, J.; Sowinski, P. Global analysis of gene expression in maize leaves treated with low temperature. II.combined effect of severe cold (8 A degrees C) and circadian rhythm. Plant Mol. Biol. 2017, 95, 279–302. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, V.M.; Romay, M.C.; Ordás, A.; Revilla, P. Evaluation of European maize (Zea mays L.) germplasm under cold conditions. Genet. Resour. Crop Evol. 2010, 57, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.X.; Yu, M.; Li, H.R.; Yin, S.P.; Li, Y.G.; Ji, Y.H. The Lowest Temperature Index in Spring and Its Effects on Crop Yield in Heilongjiang. Chin. Agric. Sci. Bull. 2018, 34, 97–103. [Google Scholar]
- Wang, C.Y. Study on Low Temperature Chilling Injury of Crops in Northeast China; China Meteorological Press: Beijing, China, 2008. [Google Scholar]
- Shi, Z.Z.; Ben, X.M.; Zhang, J.T.; Tanik, C.; Song, G.Y. The emerging pattern and preventive measure of maize cold damage in the Sanjiang River Plain. Heilong Jiang Agricutural Sci. 2003, 2, 7–10. [Google Scholar]
- Yang, R.Z.; Zhou, G.S. A Comprehensive risk assessment of the main maize agrometeorological disasters in three provinces of northeast China. Acta Meteorol. Sin. 2015, 73, 1141–1153. [Google Scholar] [CrossRef]
- Waqas, M.A.; Wang, X.K.; Zafar, S.A.; Noor, M.A.; Hussain, H.A.; Nawaz, M.A.; Farooq, M. Thermal stresses in maize: Effects and management strategies. Plants 2021, 10, 293. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.X.; Yu, Q.Q.; Ma, Y.; Gu, W.R.; Yang, D.G. Physiological changes associated with enhanced cold resistance during maize (Zea mays) germination and seedling growth in response to exogenous calcium. Crop. Pasture Sci. 2020, 71, 529–538. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. Available online: https://www.jstor.org/stable/24110209 (accessed on 7 April 2022).
- Sun, C.H.; Li, X.H.; Hu, Y.L.; Zhao, P.Y.; Gao, X.L. Proline, sugars, and antioxidant enzymes respond to drought stress in the leaves of strawberry plants. Korean J. Hortic. Sci. Technol. 2015, 33, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Venekamp, J.H.; Koot, J. The distribution of free amino acids, especially of proline, in the organs of field bean plants, Vicia faba L. during development in the field. J. Plant Physiol. 1984, 116, 343–349. [Google Scholar] [CrossRef]
- Ku, H.M.; Tan, C.W.; Su, Y.S.; Chiu, C.Y.; Chen, C.T.; Jan, F.J. The effect of water deficit and excess copper on proline metabolism in Nicotiana benthamiana. Biol. Plant. 2012, 56, 337–343. [Google Scholar] [CrossRef]
- Armengaud, P.; Thiery, L.; Buhot, N.; Grenier-de March, G.; Savoure, A. Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol. Plant. 2010, 120, 442–450. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Van Staden, J.V. A regulatory role for proline metabolism in stimulating Arabidopsis thaliana seed germination. Plant Growth Regul. 2003, 39, 41–50. [Google Scholar] [CrossRef]
- May, G.P.; Maldon, M.D. Instance of spontaneous perforation of the stomach. Lancet 1840, 34, 41–43. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, R.; Falasca, G.; Sabatini, S.; Maddalena Altamura, M.; Maurizio Trovato, M. The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development. Physiol. Plant. 2010, 137, 72–85. [Google Scholar] [CrossRef]
- Gavazzi, G.; Nava-Racchi, M.; Tonelli, C. A mutation causing proline requirement in Zea mays. Theor. Appl. Genet. 1975, 46, 339–345. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.S.; Wang, G.F.; Fan, X.Y.; Sun, X.; Qin, H.L.; Xu, N.; Zhong, M.Y.; Qiao, Z.Y.; Tang, Y.P.; et al. Proline responding1 plays a critical role in regulating general protein synthesis and the cell cycle in maize. Plant Cell 2014, 26, 2582–2600. [Google Scholar] [CrossRef] [Green Version]
- Heikal, M.D.; Shaddad M, A. Alleviation of osmotic stress on seed germination and seedling growth of cotton, pea and wheat by proline. Phyton 1982, 22, 275–287. [Google Scholar]
- Karalija, E.; Selovic, A.; Dahija, S.; Demir, A.; Samardzic, J.; Vrobel, O.; Zeljkovic, S.C.; Paric, A. Use of seed priming to improve Cd accumulation and tolerance in Silene sendtneri, novel Cd hyper-accumulator. Ecotoxicol. Environ. Saf. 2021, 210, 111882. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Balmer, Y. Redox regulation: A broadening horizon. Annu. Rev. Plant Biol. 2005, 56, 187–220. [Google Scholar] [CrossRef]
- Graham, D.; Patterson, B.D. Responses of plants to low, nonfreezing temperatures: Proteins, metabolism, and acclimation. Annu. Rev. Plant Biol. 1982, 33, 347–372. [Google Scholar] [CrossRef]
- Mizuno, N.; Sugie, A.; Kobayashi, F.; Takumi, S. Mitochondrial alternative pathway is associated with development of freezing tolerance in common wheat. J. Plant Physiol. 2008, 165, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Covey-Cump, E.M.; Attwood, R.G.; Atkin, O.K. Regulation of root respiration in two species of Plantago that differ in relative growth rate: The effect of short-and long-term changes in temperature. Plant Cell Environ. 2002, 25, 1501–1513. [Google Scholar] [CrossRef]
- Matos, A.R.; Hourton-Cabassa, C.; Cicek, D.; Reze, N.; Arrabaca, J.D.; Zachowski, A.; Moreau, F. Alternative oxidase involvement in cold stress response of Arabidopsis thaliana fad2 and FAD3+ cell suspensions altered in membrane lipid composition. Plant Cell Physiol. 2007, 48, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.J. Effects of ABA on Sugar Metabolism and Expression of Antifreeze Genes of Winter Wheat. Ph.D. Thesis, Northeast Agricultural University, Harbin, China, June 2013. [Google Scholar]
- Sarkar, D.; Bhowmik, P.C.; Young-In-Kwon; Shetty, K. Cold acclimation responses of three cool-season turfgrasses and the role of proline-associated pentose phosphate pathway. J. Am. Soc. Hortic. Sci. 2009, 134, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.K.; Sun, H.M.; Xin, X.; Qin, G.Z.; Liang, Z.; Jing, X.M. Mitochondrial damage in the soybean seed axis during imbibition at chilling temperatures. Plant Cell Physiol. 2009, 50, 1305–1318. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Cui, Y.T.; Hu, G.H.; Wang, X.D.; Chen, H.Z.; Shi, Q.H.; Xiang, J.; Zhang, Y.K.; Zhu, D.F.; Zhang, Y.P. Reduced bioactive gibberellin content in rice seeds under low temperature leads to decreased sugar consumption and low seed germination rates. Plant Physiol. Biochem. 2018, 133, 1–10. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Janowiak, F.; Maas, B.; Dörffling, K. Importance of abscisic acid for chilling tolerance of maize seedlings. J. Plant Physiol. 2002, 159, 635–643. [Google Scholar] [CrossRef]
- Huang, X.B.; Shi, H.Y.; Hu, Z.R.; Liu, A.; Amombo, E.; Chen, L.; Fu, J.M. ABA is involved in regulation of cold stress response in bermudagrass. Front. Plant Sci. 2017, 8, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J. 2004, 37, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.H.; Zhang, Y.P.; Xiang, J.; Wu, H.; Chen, H.Z.; Zhang, Y.K.; Zhu, D.F. Effects of chilling tolerance induced by spermidine pretreatment on antioxidative activity, endogenous hormones and ultrastructure of indica-japonica hybrid rice seedlings. J. Integr. Agric. 2016, 15, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sheng, J.P.; Li, S.Y.; Nie, Y.; Zhao, J.H.; Zhu, Z.; Wang, Z.D.; Tang, X.M. The role of gibberellins in the mitigation of chilling injury in cherry tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2015, 101, 88–95. [Google Scholar] [CrossRef]
- Dalla, G.C.; Scordo, E.; Allera, C.; Farina, E. Effects of low temperatures and gibberellic acid on flowering of Limonium gmelinii. Acta Hortic. 2000, 541, 323–326. [Google Scholar] [CrossRef]
- Jiao, C.; Duan, Y. The role of glycogen synthase kinase-3 in gibberellic acid-induced chilling tolerance and defense response in postharvest peach fruit. Food Bioprocess Technol. 2019, 12, 1733–1740. [Google Scholar] [CrossRef]
- Xia, J.; Shi, X.J.; Hao, X.Z.; Li, N.N.; Tian, Y.; Li, J.H.; Luo, H.H. Effects of low temperature on enzyme activity and hormone content in germination of different genotypes of cotton seeds. Plant Physiol. J. 2019, 55, 1291–1305. [Google Scholar] [CrossRef]
- Zeng, G.H.; Ma, Q.P.; Wang, W.D.; Zhou, L.; Yin, Y.; Li, X.H. Effect of Natural Low-temperature on Endogenous Hormones of Camellia sinensis (L.) Kuntze Plant. J. Tea Sci. 2016, 36, 85–91. [Google Scholar] [CrossRef]
- Yu, X.F.; Qiu, X.L.; Zhang, S.H.; Zhang, Y.F.; Zeng, M.; Zeng, L.Q.; Yang, S.M. Effects of low temperature and nitrogen coupling on endogenous hormone content of rice. J. Northwet AF Univ. 2020, 48, 73–80. [Google Scholar] [CrossRef]
- Yao, J.F.; An, J.P.; You, C.X.; Wang, X.F.; Hao, Y.J. Molecular Cloning and Tolerance Identification of Apple Cytokinin Oxidase Gene MdCKX7. 2. Acta Hortic. Sin. 2019, 46, 409–420. [Google Scholar] [CrossRef]
- Veselova, S.V.; Farhutdinov, R.G.; Veselov, S.Y.; Kudoyarova, G.R.; Veselov, D.S.; Hartung, W. The effect of root cooling on hormone content, leaf conductance and root hydraulic conductivity of durum wheat seedlings (Triticum durum L.). J. Plant Physiol. 2005, 162, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, P.; Chen, L.J.; Tao, H.B.; Wang, P. Effect of Grain Filling and Endogenous Hormones Changes on Maize Grains in Vitro Culture under Low Temperature Stress during Grain Filling Stage. J. Maize Sci. 2017, 25, 49–54. [Google Scholar] [CrossRef]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.J.; Huang, L.; Lin, X.Y.; Sun, C.L. Hydrogen peroxide alleviates salinity-induced damage through enhancing proline accumulation in wheat seedlings. Plant Cell Rep. 2020, 39, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Garcia, P.C.; López-Lefebre, L.R.; Rivero, R.M.; Ruiz, J.M.; Romero, L. Proline metabolism in response to nitrogen deficiency in French bean plants (Phaseolus vulgaris L. cv Strike). Plant Growth Regul. 2002, 36, 261–265. [Google Scholar] [CrossRef]
- Wei, M.; Zhuang, Y.; Li, H.; Li, P.H.; Huo, H.Q.; Shu, D.; Huang, W.Z.; Wang, S.H. The cloning and characterization of hypersensitive to salt stress mutant, affected in quinolinate synthase, highlights the involvement of NAD in stress-induced accumulation of ABA and proline. Plant J. 2020, 102, 85–98. [Google Scholar] [CrossRef]
- Hamilton, E.W.; Heckathorn, S.A. Mitochondrial adaptations to NaCl. complex I is protected by anti-oxidants and small heat shock proteins, whereas complex II is protected by proline and betaine. Plant Physiol. 2001, 126, 1266–1274. [Google Scholar] [CrossRef] [Green Version]
- Banu, N.A.; Hoque, A.; Watanabe-Sugimoto, M.; Matsuoka, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Proline and glycinebetaine induce antioxidant defense gene expression and suppress cell death in cultured tobacco cells under salt stress. J. Plant Physiol. 2009, 166, 146–156. [Google Scholar] [CrossRef]
- Naliwajski, M.; Skodowska, M. The relationship between the antioxidant system and proline metabolism in the leaves of cucumber plants acclimated to salt stress. Cells 2021, 10, 609. [Google Scholar] [CrossRef]
- Franco, A.C.; Ball, E.; Luttge, U. The influence of nitrogen, light and water stress on CO2 exchange and organic acid accumulation in the tropical C3-CAM tree, Clusia minor. J. Exp. Bot. 1991, 42, 597–603. [Google Scholar] [CrossRef]
- Wattanakulpakin, P.; Photchanachai, S.; Miyagawa, S.; Ratanakhanokchai, K. Loss of maize seed vigor as affected by biochemical changes during hydropriming. Crop Sci. 2012, 52, 2783. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Garcia-Rios, M.; Fujita, T.; Larosa, P.C.; Locy, R.D.; Clithero, J.M.; Csonka, B.L.N. Cloning of a polycistronic cDNA from tomato encoding γ-glutamyl kinase and γ-glutamyl phosphate reductase. Proc. Natl. Acad. Sci. USA 1997, 94, 8249–8254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, R.H.; Kopac, M.J. Some properties of ornithine δ-transaminase from Neurospora. Biochim. Biophys. Acta 1960, 37, 539–540. [Google Scholar] [CrossRef]
- Yang, S.L.; Lan, S.S.; Gong, M. Hydrogen peroxide-induced proline and metabolic pathway of its accumulation in maize seedlings. J. Plant Physiol. 2009, 166, 1694–1699. [Google Scholar] [CrossRef]
- Deng, Y.; Kohlwein, S.D.; Mannella, C.A. Fasting induces cyanide-resistant respiration and oxidative stress in the amoeba Chaos carolinensis: Implications for the cubic structural transition in mitochondrial membranes. Protoplasma 2002, 219, 160–167. [Google Scholar] [CrossRef]
- Stamp, P. Activities of phosphofructokinase and cytochrome-c-oxidase in leaves of maize seedlings (Zea mays L.) in relation to genotype and to temperature changes. Z. Für Pflanzenphysiol. 1981, 103, 131–137. [Google Scholar] [CrossRef]
- Umbach, A.L.; Siedow, J.N. Covalent and noncovalent dimers of the cyanide-resistant alternative oxidase protein in higher plant mitochondria and their relationship to enzyme activity. Plant Physiol. 1993, 103, 845–854. [Google Scholar] [CrossRef]
- Braithwaite, S.S.; Palazuk, B.; Colca, J.R.; Edwards, C.W., 3rd; Hofmann, C. Reduced expression of hexokinase II in insulin-resistant diabetes. Diabetes 1995, 44, 43–51. [Google Scholar] [CrossRef]
- Gajewska, E.; Niewiadomska, E.; Tokarz, K.; Słaba, M.; Skłodowska, M. Nickel-induced changes in carbon metabolism in wheat shoots. J. Plant Physiol. 2013, 170, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.C.; Song, J.H.; Weng, Q.Y.; Ma, H.L.; Wang, L.Y.; Liu, Y.H. Cloning and Character Analysis of NADP+-Dependent Isocitrate Dehydrogenase Gene in Maize. Plant Physiol. J. 2015, 51, 481–487. [Google Scholar] [CrossRef]
- Kumar, R.G.; Shah, K.; Dubey, R.S. Salinity induced behavioural changes in malate dehydrogenase and glutamate dehydrogenase activities in rice seedlings of differing salt tolerance. Plant Sci. 2000, 156, 23–34. [Google Scholar] [CrossRef]
- Cooper, T.G.; Beevers, H. Mitochondria and glyoxysomes from castor bean endosperm enzyme constitutents and catalytic capacity. J. Biol. Chem. 1969, 244, 3507–3513. [Google Scholar] [CrossRef]
- Pu, X.C.; Han, J.G.; Li, M.; Du, G.P.; Ni, X.Q. Studies on Respiratory Pathways of Zoysiagrass Seed When Breaking Dormancy. Acta Prataculturae Sin. 1996, 5, 57–61. [Google Scholar]
- Wan, Z.H.; Ren, B.Z.; Zhao, B.; Liu, P.; Dong, S.T.; Zhang, J.W. Grain filling, dehydration characteristics and changes of endogenous hormones of summer maize hybrids differing in maturities. Acta Agron. Sinica 2019, 45, 1446–1453. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Janas, K.M. Effects of seed hydropriming in presence of exogenous proline on chilling injury limitation in Vigna radiata L. seedlings. Acta Physiol. Plant. 2007, 29, 509–517. [Google Scholar] [CrossRef]
- Khajeh-Hossein, M.; Powell, A.A.; Bingham, I.J. The interaction between salinity stress and seed vigour during germination of soyabean seeds. Seed Sci. Technol. 2003, 31, 715–725. [Google Scholar] [CrossRef]
- Perruc, E.; Kinoshita, N.; Lopez-Molina, L. The role of chromatin-remodeling factor PKL in balancing osmotic stress responses during Arabidopsis seed germination. Plant J. 2010, 52, 927–936. [Google Scholar] [CrossRef]
- Rajjou, L.; Duva, L.M.; Gallardo, K.; Catusse, J.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Dodd, G.L.; Donovan, L.A. Water potential and ionic effects on germination and seedling growth of two cold desert shrubs. Am. J. Bot. 1999, 86, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.W.; Zhong, P.; Liu, J.; Tang, Z.H.; Gao, Y.B.; Yu, H.J.; Guo, W. Effect of low-temperature stress and gibberellin on seed germination and seedling physiological responses in peanut. Acta Agron. Sin. 2019, 45, 118–130. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Cao, D.D.; Zhang, S.; Guan, Y.J.; Hu, J. Effect of Polyamines on Chilling Tolerance in Seed Imbibition and Seed Germination in Maize. Acta Agron. Sin. 2008, 2, 261–267. [Google Scholar] [CrossRef]
- Nawaz, K.; Talat, A.; Iqra, K.H.; Majeed, A. Induction of salt tolerance in two cultivars of sorghum (Sorghum bicolor L.) by exogenous application of proline at seedling stage. World Appl. Sci. J. 2013, 10, 93–99. [Google Scholar]
- Yaqoob, H.; Akram, N.A.; Iftikhar, S.; Ashraf, M.; Khalid, N.; Sadiq, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Seed pretreatment and foliar application of proline regulate morphological, physio-biochemical processes and activity of antioxidant enzymes in plants of two cultivars of quinoa (Chenopodium quinoa Willd). Plants 2019, 8, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rady, M.M.; Kuşvuran, A.; Alharby, H.F.; Alzahrani, Y.; Kuşvuran, S. Pretreatment with proline or an organic bio-stimulant induces salt tolerance in wheat plants by improving antioxidant redox state and enzymatic activities and reducing the oxidative stress. J. Plant Growth Regul. 2019, 38, 449–462. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Kubala, S.; Wojtyla, L.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced expression of the proline synthesis gene P5CSA in relation to seed osmopriming improvement of Brassica napus germination under salinity stress. J. Plant Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Lu, Q.; Verma, D.P.S. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase, a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J. Biol. Chem. 1995, 270, 20491–20496. [Google Scholar] [CrossRef] [Green Version]
- Egert, A.; Keller, F.; Peters, S. Abiotic stress-induced accumulation of raffinose in Arabidopsis leaves is mediated by a single raffinose synthase (RS5, At5 g40390). BMC Plant Biol. 2013, 13, 218. [Google Scholar] [CrossRef] [Green Version]
- Li, L.J.; Gu, W.R.; Li, J.; Li, C.F.; Xie, T.L.; Qu, D.Y.; Meng, Y.; Li, C.F.; Wei, S. Exogenously applied spermidine alleviates photosynthetic inhibition under drought stress in maize (Zea mays L.) seedlings associated with changes in endogenous polyamines and phytohormones. Plant Physiol. Biochem. 2018, 129, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.J.; Zheng, Y.T.; Zheng, Y.Y.; Xu, S.C. Polyamines are involved in chilling tolerance in tobacco (Nicotiana tabacum) seedlings. Plant Growth Regul. 2019, 89, 153–166. [Google Scholar] [CrossRef]
- Choudhary, N.L.; Sairam, R.K.; Tyagi, A. Expression of Δ1-pyrroline-5-carboxylate synthetase gene during drought in rice (Oryza sativa L.). Indian J. Biochem. Biophys. 2005, 42, 366–370. [Google Scholar]
- Kishor, P.B.K.; Hong, Z.; Miao, G.H.; Hu, C.A.A.; Verma, D.P.S. Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol. 1995, 108, 1387–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.R.; Guo, S.R.; Sun, J.; Cheng, Y.J.; Liu, C.J.; Wang, L.P. Effects of Proline on Reactive Oxygen Metabolism and Content of Osmotic Adjustment Substances of Cucumber under High Temperature Stress. Acta Agric. Boreali-Occident. Sin. 2010, 19, 127–131. [Google Scholar]
- Plaxton, W.C.; Podestá, F.E. The functional organization and control of plant respiration. Crit. Rev. Plant Sci. 2006, 25, 159–198. [Google Scholar] [CrossRef]
- Dahal, K.; Vanlerberghe, G.C. Alternative oxidase respiration maintains both mitochondrial and chloroplast function during drought. New Phytol. 2016, 213, 560–571. [Google Scholar] [CrossRef]
- Thomas, Y.; Araújo, W.L.; Allen, A.E.; Fernie, A.R.; Bowler, C. Downregulation of mitochondrial alternative oxidase affects chloroplast function, redox status and stress response in a marine diatom. New Phytol. 2018, 221, 1303–1316. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.C. The Effects of Different Temperature Stresses and Salicylic Acid Treatment on Cyanide-Resistant Respiration and the Molecular Biology Studies on Expression of Alternative Oxidase Gene in Tobacco Callus. Master’s Thesis, Sichuan University, Chengdu, China, April 2002. [Google Scholar]
- Lam, E.; Kato, N.; Lawton, M. Programmed cell death, mitochondria and the plant hypersensitive response. Nature 2001, 411, 848–853. [Google Scholar] [CrossRef]
- Wang, H.; Huang, J.; Liang, X.; Bi, Y.L. Involvement of hydrogen peroxide, calcium, and ethylene in the induction of the alternative pathway in chilling-stressed Arabidopsis callus. Planta 2012, 235, 53–67. [Google Scholar] [CrossRef]
- Hao, J.; Li, X.; Xu, G.; Yang, H. Exogenous progesterone treatment alleviates chilling injury in postharvest banana fruit associated with induction of alternative oxidase and antioxidant defense. Food Chem. 2019, 286, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Rumiantsev, A.M.; Padkina, M.V.; Sambuk, E.V. Effect of nitrogen source on gene expression of first steps of methanol utilization pathway in Pichia pastoris. Russ. J. Genet. 2013, 49, 394–400. [Google Scholar] [CrossRef]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plant. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef]
- Dong, W.K.; Ma, X.; Zhang, Y.J.; Guo, R.; Zhang, Z.H.; Zhang, Y.; Ma, H.L. Effects of low-temperature stress on glycolysis metabolism and related gene expression of different Poa pratensis varieties. Acta Agrestia Sin. 2019, 27, 1503–1510. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Muller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Chen, L.; Zhou, Y.; Thomas, P.; Mawhinney, B. Functional characterization of Arabidopsis thaliana isopropylmalate dehydrogenases reveals their important roles in gametophyte development. New Phytol. 2010, 189, 160–175. [Google Scholar] [CrossRef]
- Venkat, S.; Gregory, C.; Sturges, J.; Gan, Q.; Fan, C. Studying the lysine acetylation of malate dehydrogenase. J. Mol. Biol. 2017, 429, 1396–1405. [Google Scholar] [CrossRef]
- Ying, J.; Lee, E.A.; Tollenaar, M. Response of maize leaf photosynthesis to low temperature during the grain-filling period. Field Crop. Res. 2000, 68, 87–96. [Google Scholar] [CrossRef]
- Sun, H.; Lin, L.; Xu, W.; Wu, S.; Wang, X. Ascorbate-glutathione cycle of mitochondria in osmoprimed soybean cotyledons in response to imbibitional chilling injury. J. Plant Physiol. 2011, 168, 226–232. [Google Scholar] [CrossRef]
- Bunik, V.I.; Fernie, A.R. Metabolic control exerted by the 2-oxoglutarate dehydrogenase reaction: A cross-kingdom comparison of the crossroad between energy production and nitrogen assimilation. Biochem. J. 2009, 422, 405–421. [Google Scholar] [CrossRef] [Green Version]
- Berley, J.D.; Black, M. Physiology and Biochemistry of Seeds; Spring: Berlin/Heidelberg, Germany, 1988. [Google Scholar]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A. Primary, preventive and permissive roles of hormones in plant systems. Bot. Rev. 1975, 41, 391–420. [Google Scholar] [CrossRef]
- Liu, Y.X.; Shi, K.M.; Lin, J.; Cui, L.J. Response of endogenous hormone to temperatures changes during the seed germination of Phoebe zhennan. J. Cent. South Univ. For. Technol. 2020, 40, 89–95. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Effects of exogenous abscisic acid on some physiological responses in a popular aromatic indica rice compared with those from two traditional non-aromatic indica rice cultivars. Acta Physiol. Plant. 2009, 31, 915–926. [Google Scholar] [CrossRef]
- Sharma, S.; Verslues, P.E. Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant Cell Environ. 2010, 33, 1838–1851. [Google Scholar] [CrossRef]
- Cadman, C.S.C.; Toorop, P.E.; Hilhorst, H.W.M.; Finch-Savage, W.E. Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism. Plant J. 2010, 46, 805–822. [Google Scholar] [CrossRef]
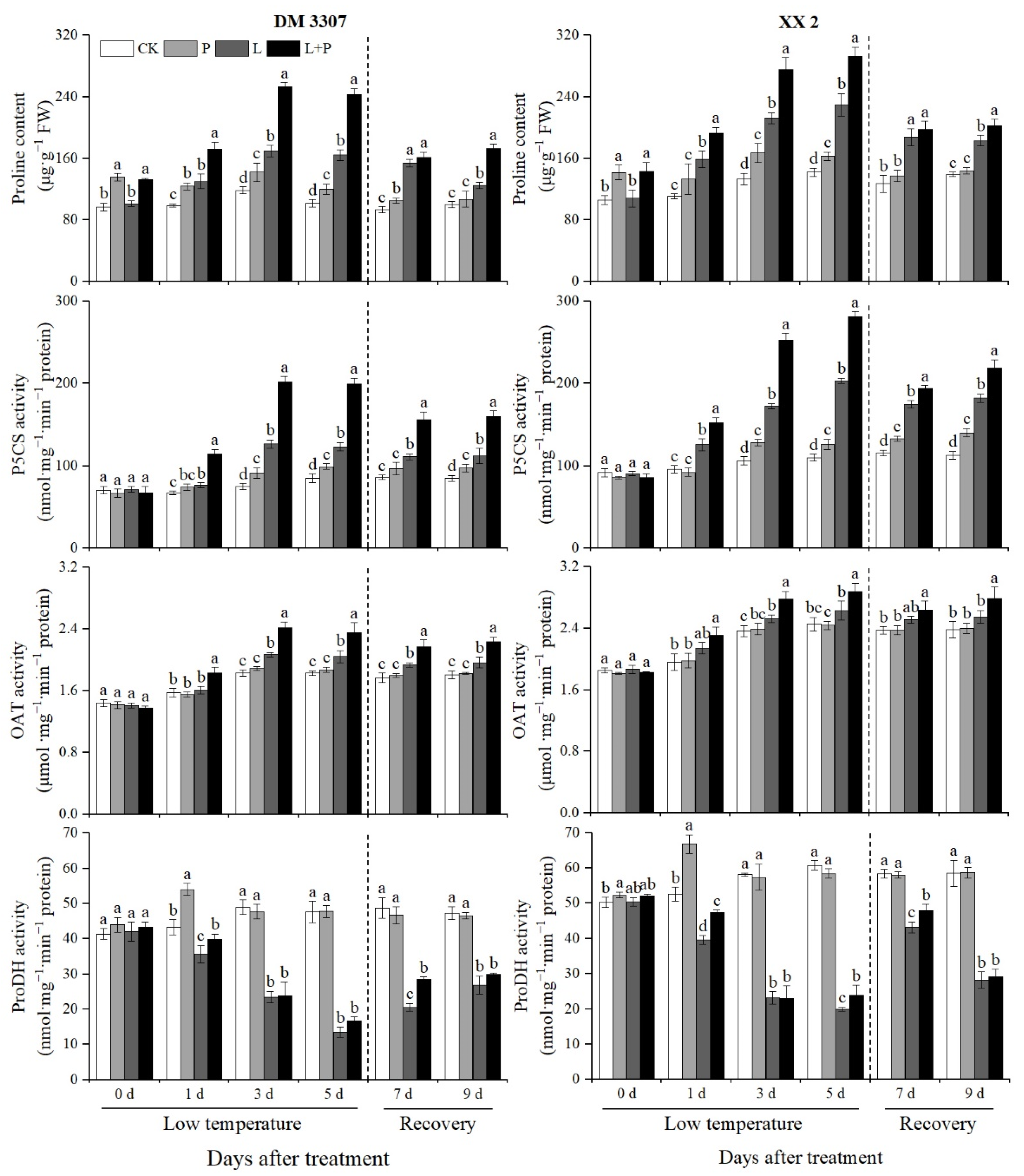
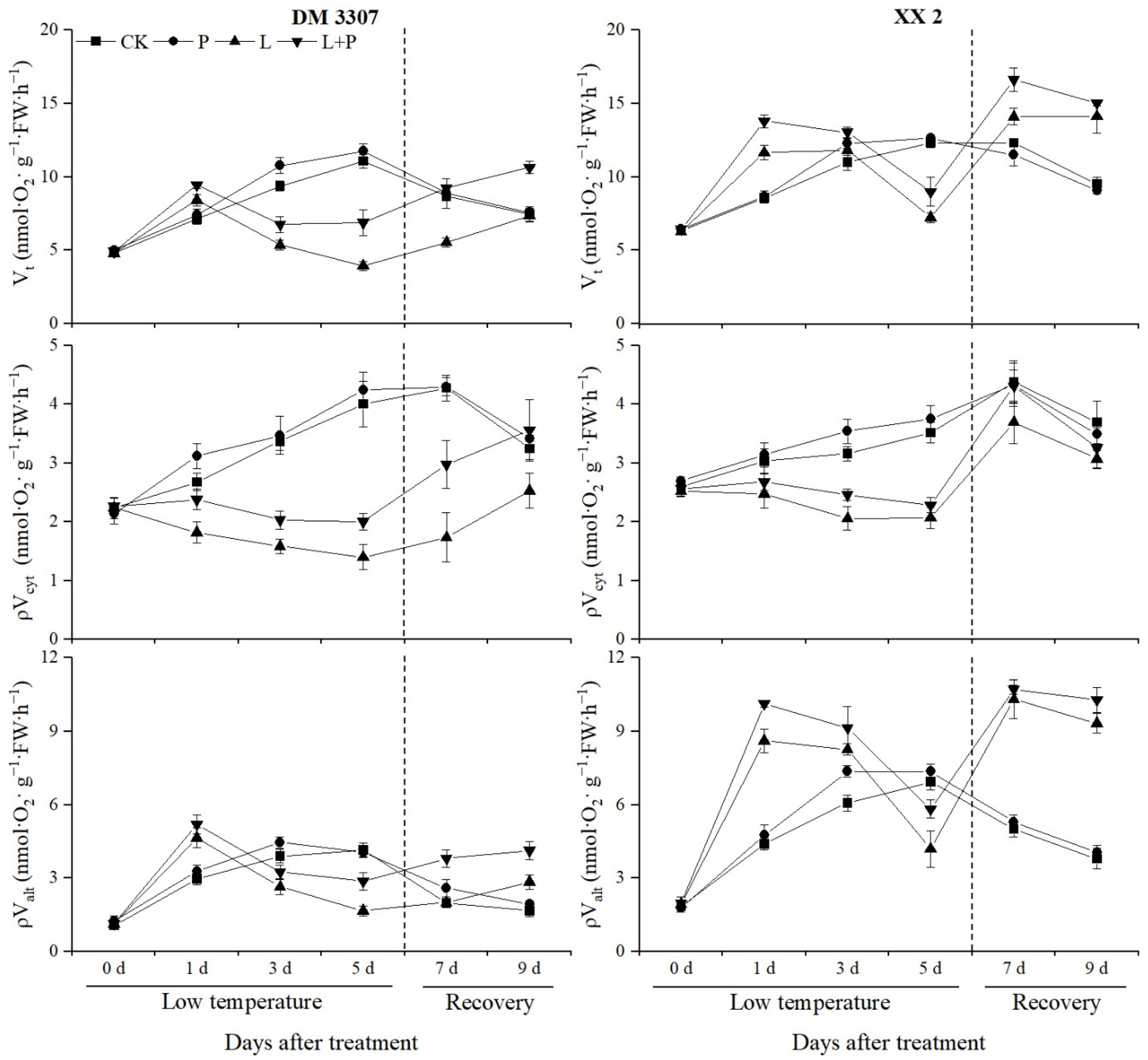
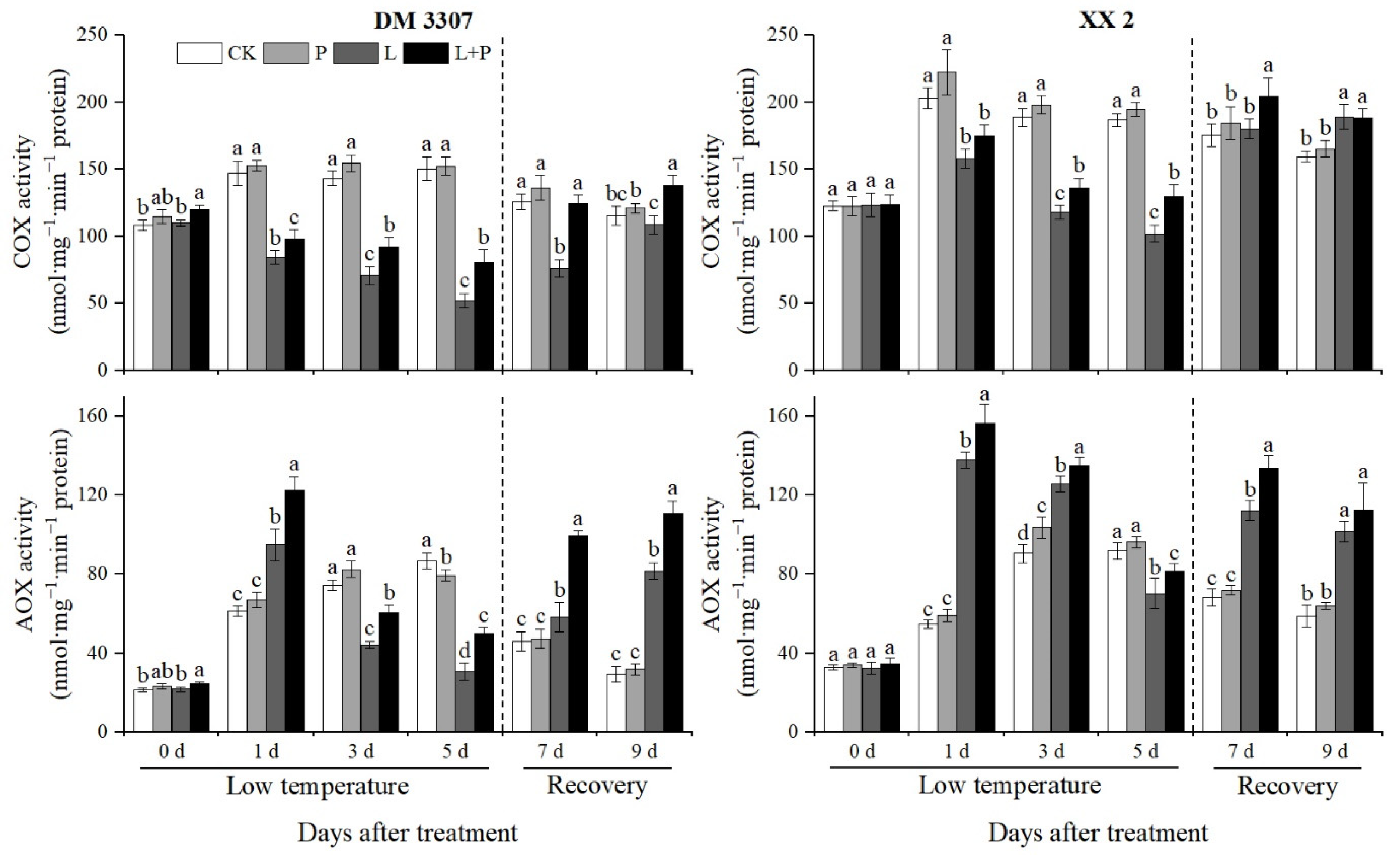
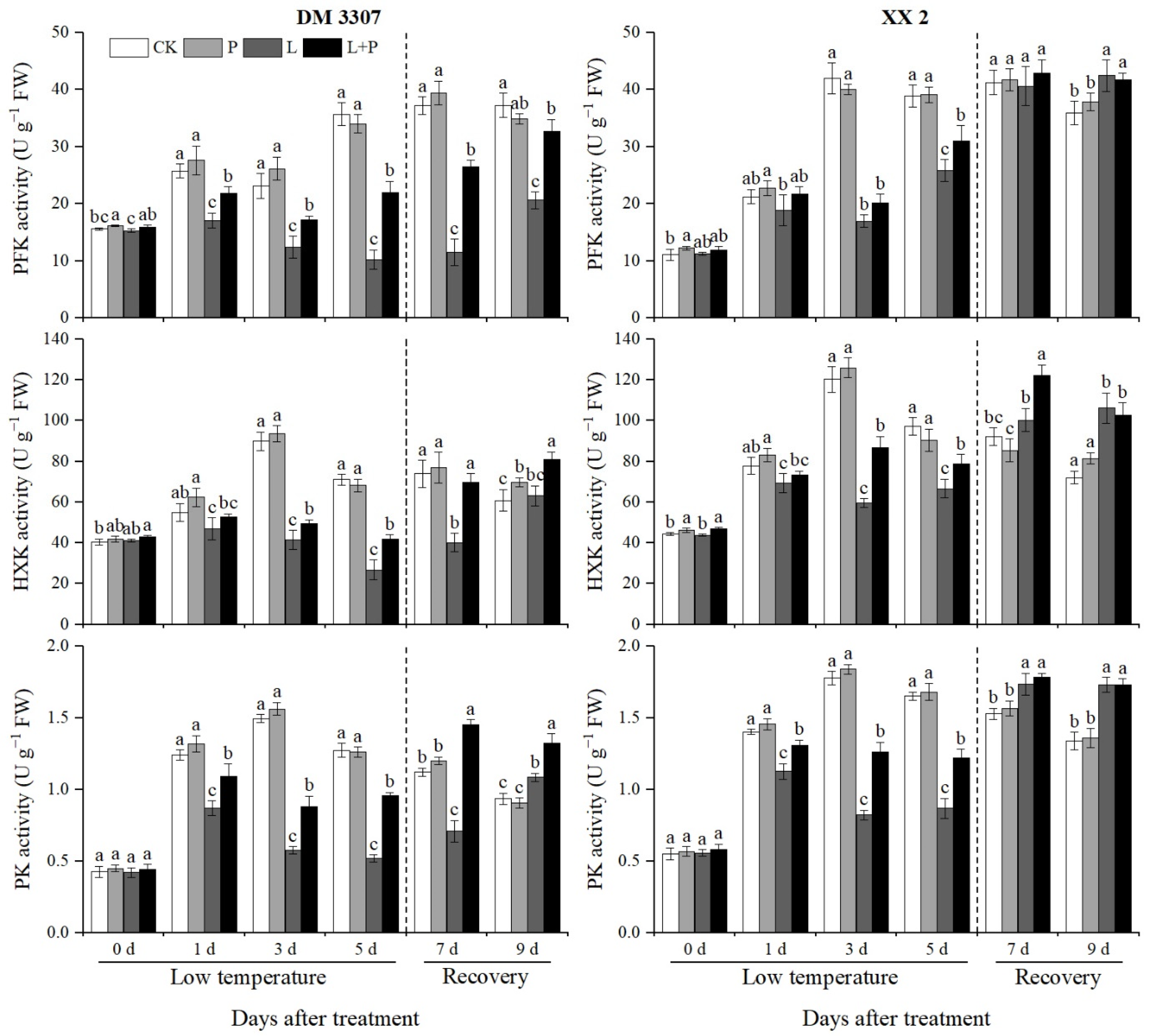
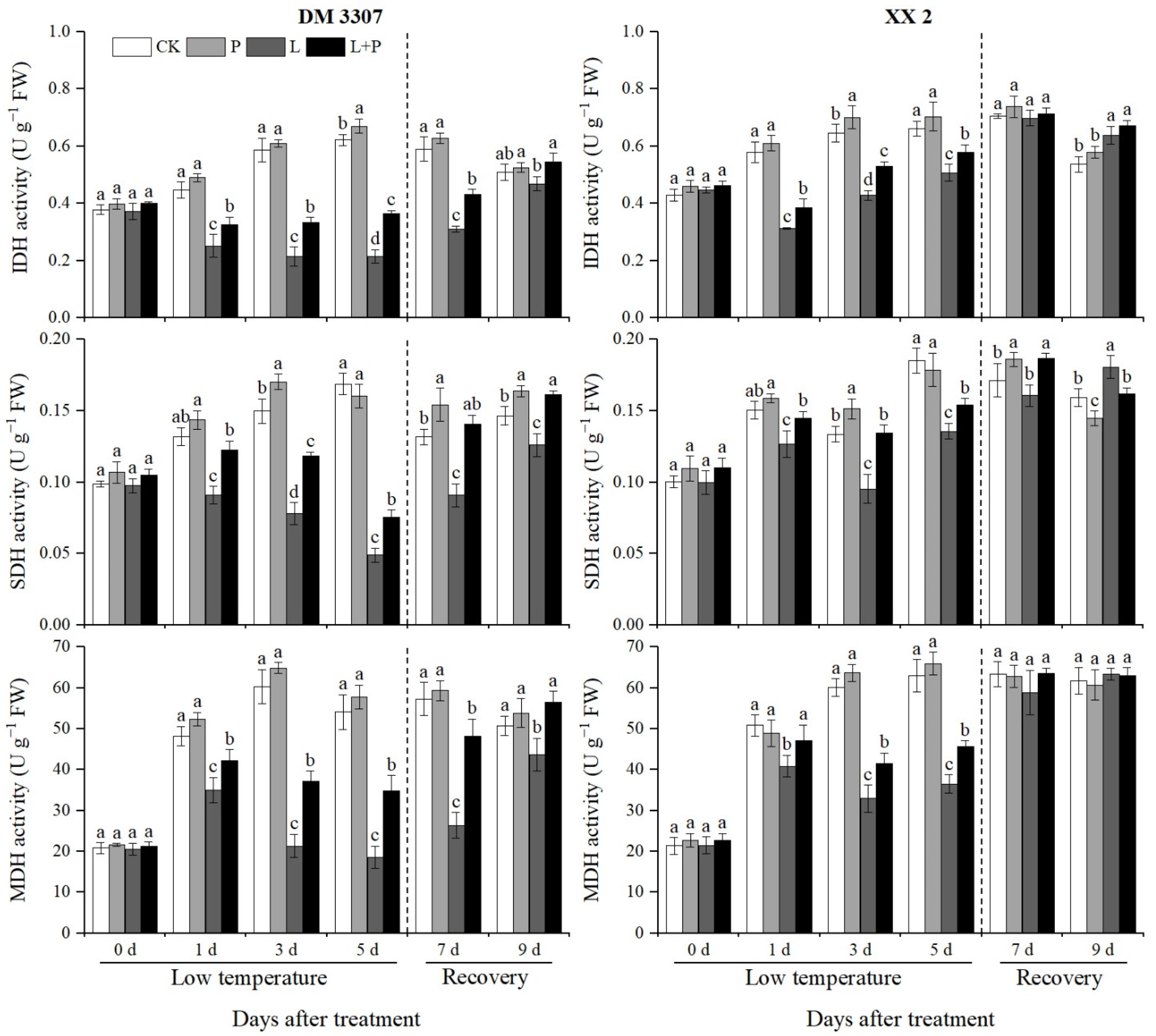
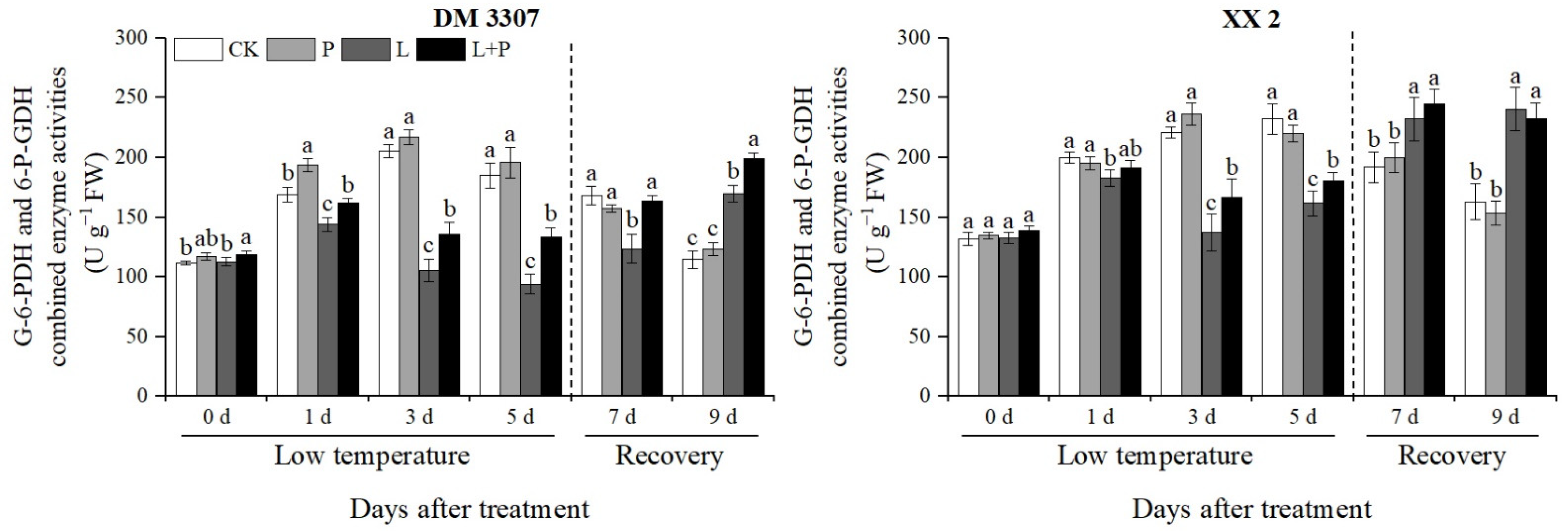
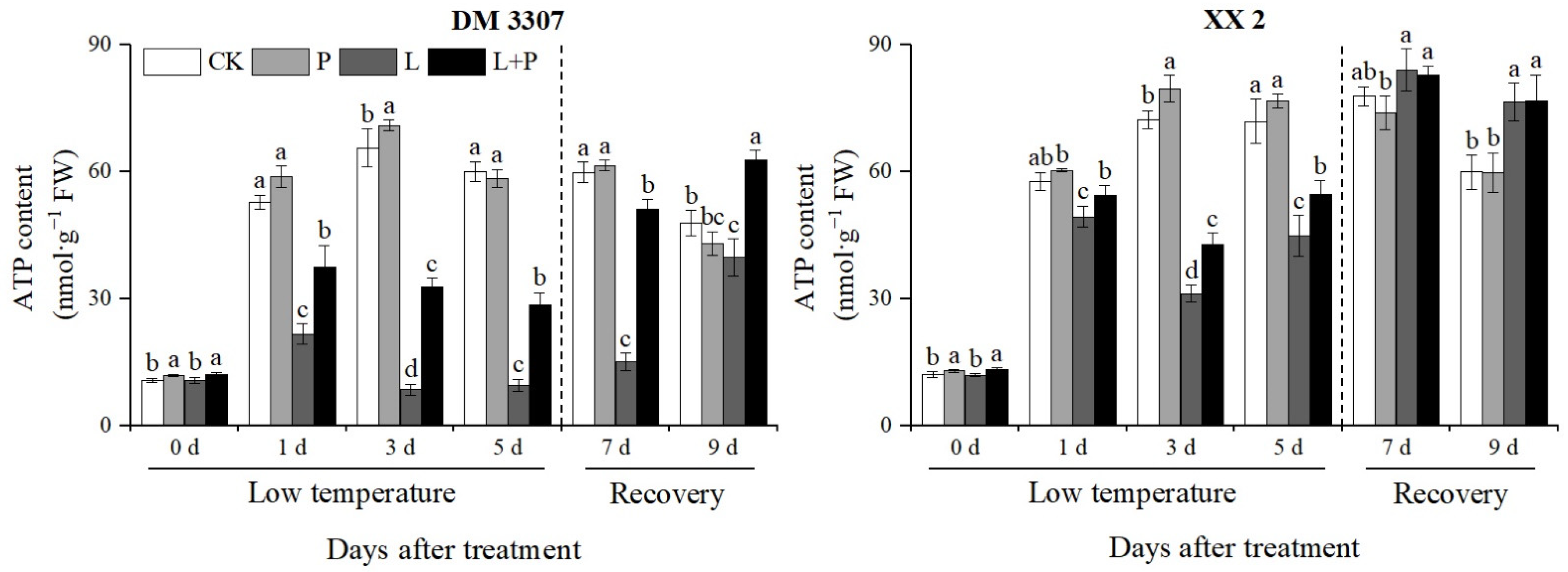
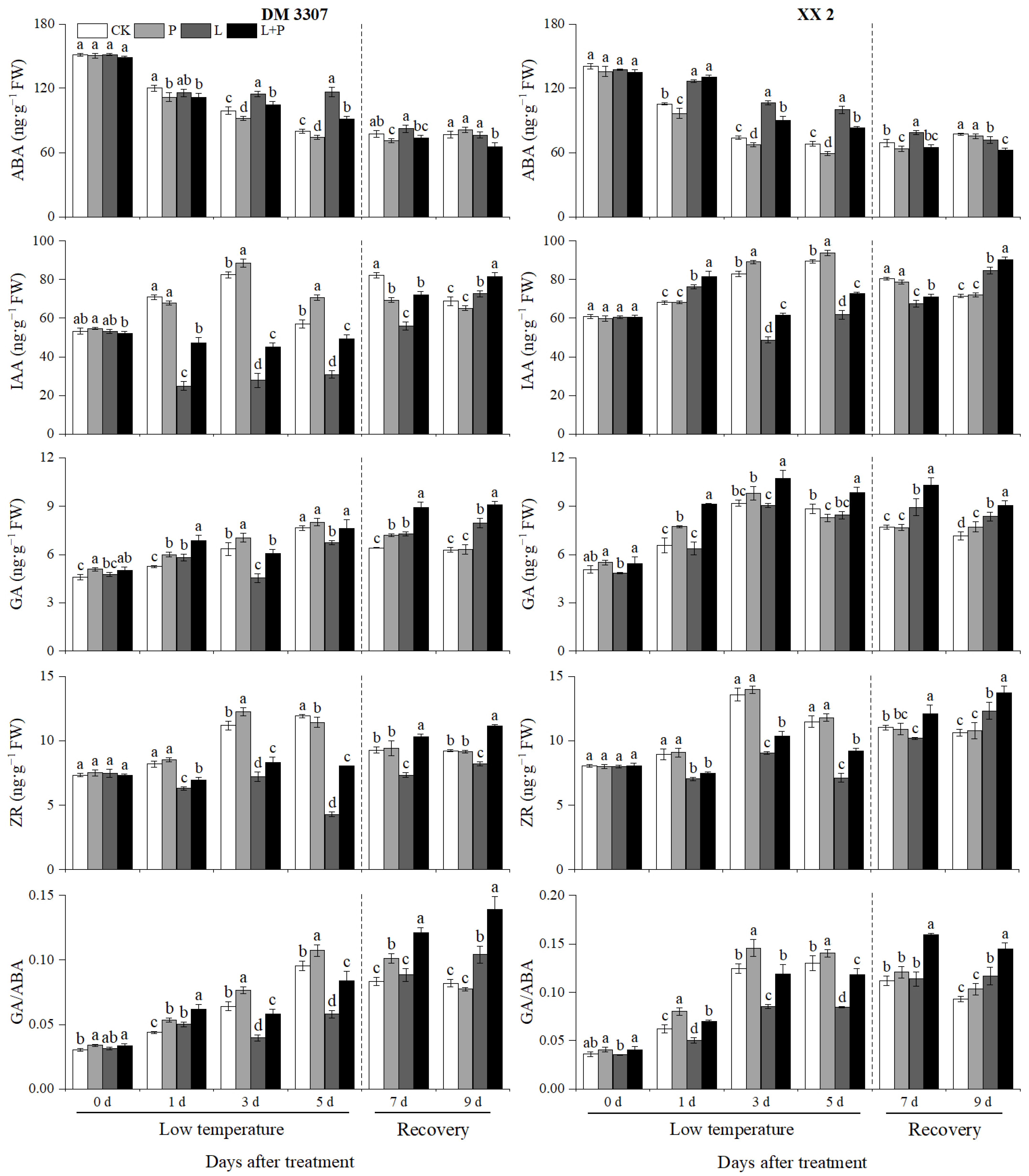
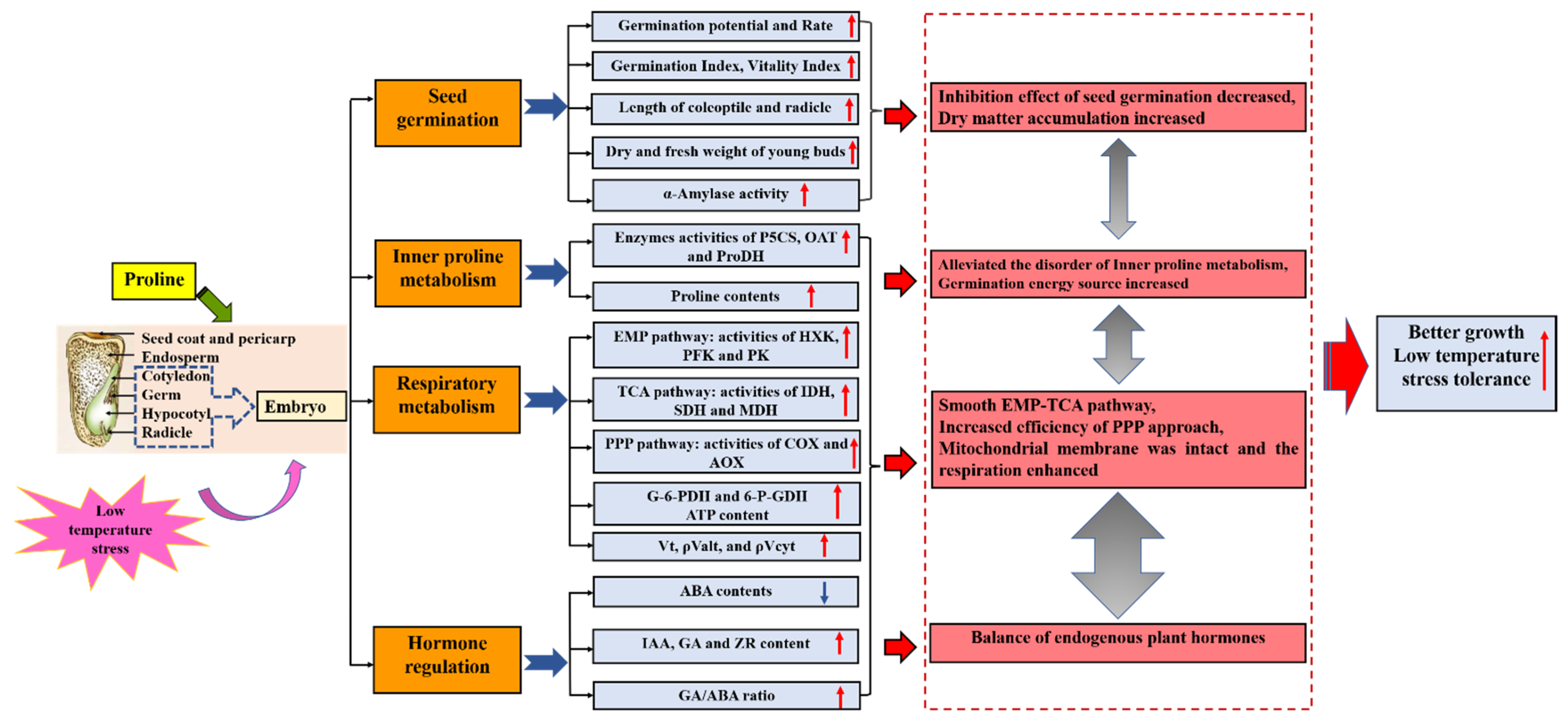
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).