
Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.)

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Treatments
2.3. Plant Growth Parameters
2.4. Physio-Biochemical Parameters
2.5. Mineral Analysis in Roots and Leaves
2.6. Proline and Lipid Peroxidation Analysis in Samples
2.7. Statistical Analysis
3. Results
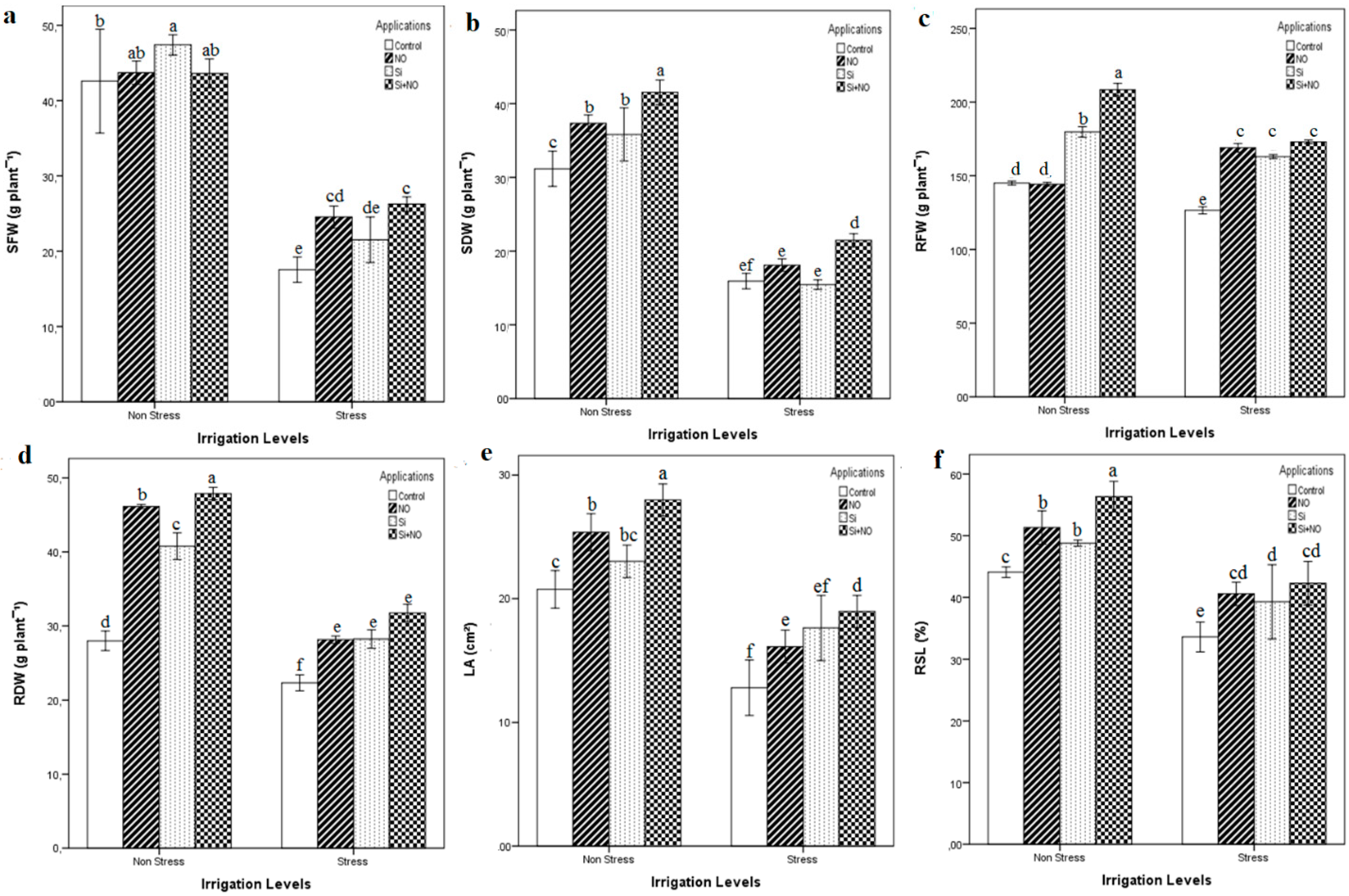
3.1. Plant Growth Parameters
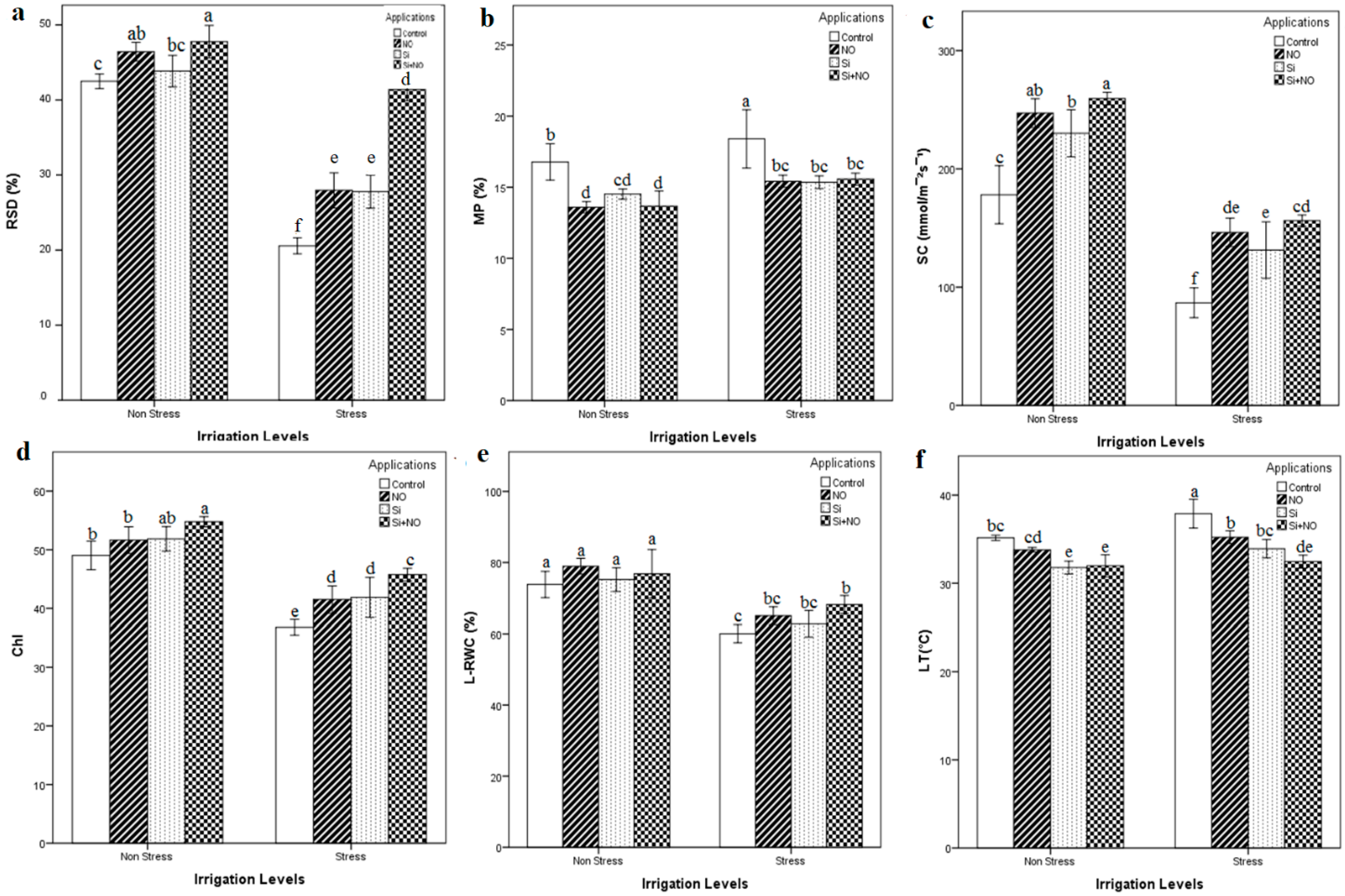
3.2. Physio-Biochemical Parameters
3.3. Minerals in Roots and Leaves
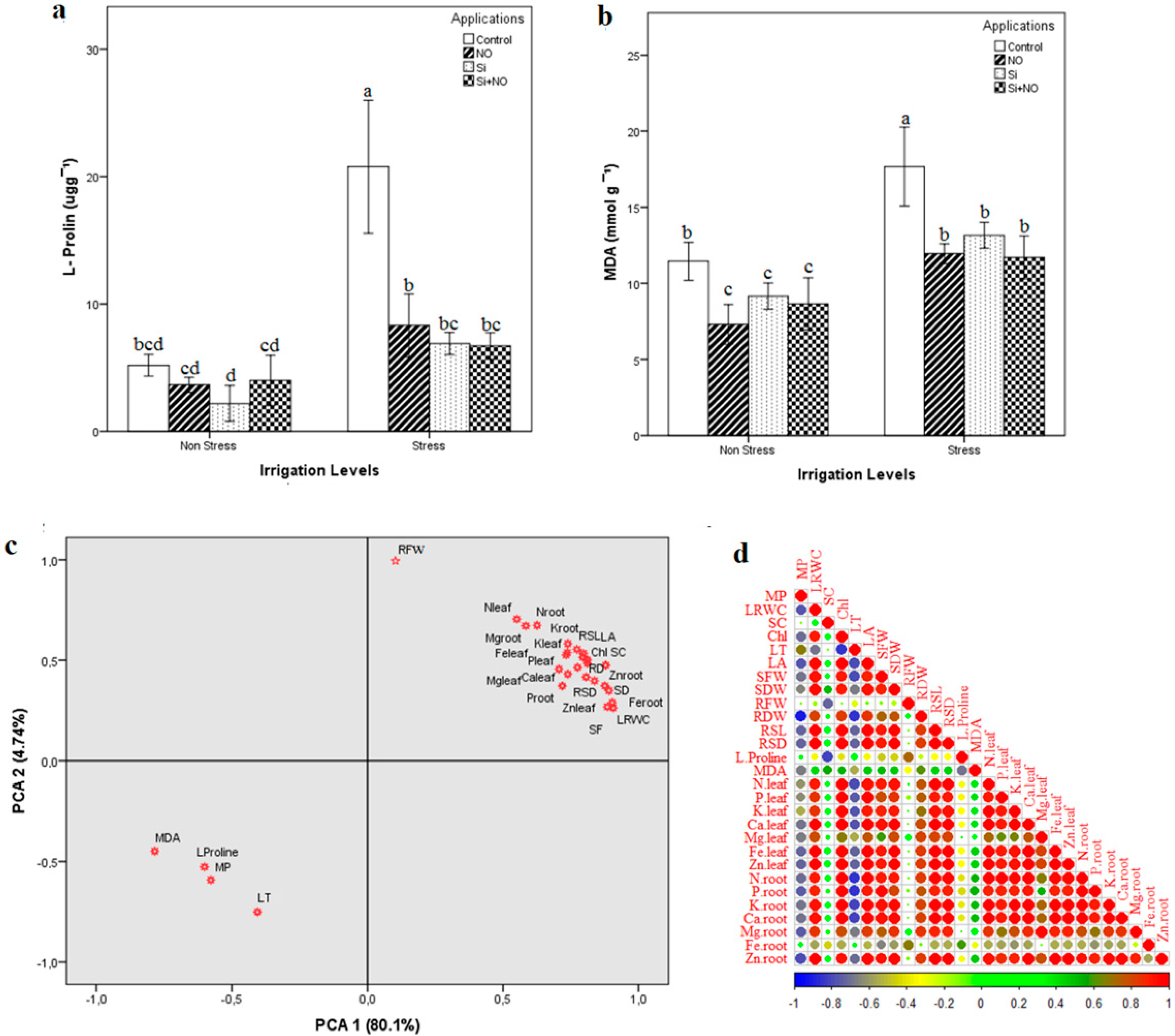
3.4. Proline and Lipid Peroxidation Levels, and Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez-Gamboa, G.; Guerrero-Méndez, M.; Araya-Alman, M.; Verdugo-Vásquez, N.; Valdés-Gómez, H.; Acevedo-Opazo, C. Sunscreen Based on Dicarboxylic Acid Salts Applications to Blueberries (Vaccinium Corymbosum L.) Plants: Effects on Water Stress Tolerance and Productivity. Horticulturae 2022, 8, 95. [Google Scholar] [CrossRef]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Shiranibidabadi, S. Effect of Silicon on Growth and Development of Strawberry under Water Deficit Conditions. Hortic. Plant J. 2018, 4, 226–232. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Zheng, W.; Martínez de Toda, F. Current Viticultural Techniques to Mitigate the Effects of Global Warming on Grape and Wine Quality: A Comprehensive Review. Food Res. Int. 2021, 139, 109946. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.Y.; Tian, S.S.; Liu, S.S.; Wang, W.Q.; Sui, N. Energy Dissipation and Antioxidant Enzyme System Protect Photosystem II of Sweet Sorghum under Drought Stress. Photosynthetica 2017, 56, 861–872. [Google Scholar] [CrossRef]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought Stress in Sunflower: Physiological Effects and Its Management through Breeding and Agronomic Alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Ramos-Artuso, F.; Galatro, A.; Buet, A.; Santa-María, G.E.; Simontacchi, M. Key Acclimation Responses to Phosphorus Deficiency in Maize Plants Are Influenced by Exogenous Nitric Oxide. J. Plant Physiol. 2018, 222, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-Induced NO Generation and Stomatal Closure in Arabidopsis Are Dependent on H2O2 Synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- Leshem, Y.Y.; Haramaty, E. The Characterization and Contrasting Effects of the Nitric Oxide Free Radical in Vegetative Stress and Senescence of Pisum Sativum Linn. Foliage. J. Plant Physiol. 1996, 148, 258–263. [Google Scholar] [CrossRef]
- Merwad, A.R.M.A.; Desoky, E.S.M.; Rady, M.M. Response of Water Deficit-Stressed Vigna Unguiculata Performances to Silicon, Proline or Methionine Foliar Application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Mata, C.G.; Lamattina, L. Nitric Oxide Induces Stomatal Closure and Enhances the Adaptive Plant Responses against Drought Stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous Silicon (Si) Increases Antioxidant Enzyme Activity and Reduces Lipid Peroxidation in Roots of Salt-Stressed Barley (Hordeum Vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Ahire, M.L.; Mundada, P.S.; Nikam, T.D.; Bapat, V.A.; Penna, S. Multifaceted Roles of Silicon in Mitigating Environmental Stresses in Plants. Plant Physiol. Biochem. 2021, 169, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, R.; Mur, L.A.J.; Ruan, J.; Shen, Q.; Guo, S. Functions of Silicon in Plant Drought Stress Responses. Hortic. Res. 2021, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.E.; Hamdan, M.F.; Pua, T.L.; Saidi, N.B.; Tan, B.C. Plant Nitric Oxide Signaling under Drought Stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef]
- Farnese, F.S.; Menezes-Silva, P.E.; Gusman, G.S.; Oliveira, J.A. When Bad Guys Become Good Ones: The Key Role of Reactive Oxygen Species and Nitric Oxide in the Plant Responses to Abiotic Stress. Front. Plant Sci. 2016, 7, 471. [Google Scholar] [CrossRef] [Green Version]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-María, G.E. Plant Survival in a Changing Environment: The Role of Nitric Oxide in Plant Responses to Abiotic Stress. Front. Plant Sci. 2015, 6, 977. [Google Scholar] [CrossRef]
- Pirooz, P.; Amooaghaie, R.; Ahadi, A.; Sharififar, F.; Torkzadeh-Mahani, M. Silicon and Nitric Oxide Synergistically Modulate the Production of Essential Oil and Rosmarinic Acid in Salvia Officinalis under Cu Stress. Protoplasma 2022, 259, 905–916. [Google Scholar] [CrossRef]
- Badem, A.; Söylemez, S. Effects of Nitric Oxide and Silicon Application on Growth and Productivity of Pepper under Salinity Stress. J. King Saud Univ.—Sci. 2022, 34, 102189. [Google Scholar] [CrossRef]
- Liu, X.; Yin, L.; Deng, X.; Gong, D.; Du, S.; Wang, S.; Zhang, Z. Combined Application of Silicon and Nitric Oxide Jointly Alleviated Cadmium Accumulation and Toxicity in Maize. J. Hazard. Mater. 2020, 395, 122679. [Google Scholar] [CrossRef]
- Pirooz, P.; Amooaghaie, R.; Ahadi, A.; Sharififar, F. Silicon-Induced Nitric Oxide Burst Modulates Systemic Defensive Responses of Salvia Officinalis under Copper Toxicity. Plant Physiol. Biochem. 2021, 162, 752–761. [Google Scholar] [CrossRef]
- Liyanage, D.K.; Chathuranga, I.; Mori, B.A.; Thilakarathna, M.S. A Simple, Semi-Automated, Gravimetric Method to Simulate Drought Stress on Plants. Agronomy 2022, 12, 349. [Google Scholar] [CrossRef]
- Patil, A.A.; Durgude, A.G.; Pharande, A.L.; Kadlag, A.D.; Nimbalkar, C.A. Effect of Calcium Silicate as a Silicon Source on Growth and Yield of Rice Plants. Int. J. Chem. Stud. 2017, 5, 545–549. [Google Scholar]
- Habibi, G. Effects of Soil- and Foliar-Applied Silicon on the Resistance of Grapevine Plants to Freezing Stress. Acta Biol. Szeged. 2015, 59, 109–117. [Google Scholar]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric Oxide Regulates Plant Responses to Drought, Salinity, and Heavy Metal Stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Calif. Agric. Exp. Stn. 1990, 347, 32. [Google Scholar]
- Abràmoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Klamkowski, K.; Treder, W. Response to Drought Stress of Three Strawberry Cultivars Grown under Greenhouse Condition. J. Fruit Ornam. Plant Res. 2008, 16, 179–188. [Google Scholar]
- Bolat, I.; Dikilitas, M.; Ikinci, A.; Ercisli, S.; Tonkaz, T. Morphological, Physiological, Biochemical Characteristics and Bud Success Responses of Myrobolan 29 c Plum Rootstock Subjected to Water Stress. Can. J. Plant Sci. 2016, 96, 485–493. [Google Scholar] [CrossRef]
- Yamasaki, S.; Dillenburg, L.R. Measurements of Leaf Relative Water Content in Araucaria Angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Jara-Rojas, F.; Ortega-Farías, S.; Valdés-Gómez, H.; Acevedo-Opazo, C. Gas Exchange Relations of Ungrafted Grapevines (Cv. Carménère) Growing under Irrigated Field Conditions. S. Afr. J. Enol. Vitic. 2015, 36, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Opazo, C.; Valdés-Gómez, H.; Taylor, J.A.; Avalo, A.; Verdugo-Vásquez, N.; Araya, M.; Jara-Rojas, F.; Tisseyre, B. Assessment of an Empirical Spatial Prediction Model of Vine Water Status for Irrigation Management in a Grapevine Field. Agric. Water Manag. 2013, 124, 58–68. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Pérez-Donoso, A.G.; Pou-Mir, A.; Acevedo-Opazo, C.; Valdés-Gómez, H. Hydric Behaviour and Gas Exchange in Different Grapevine Varieties (Vitis Vinifera L.) from the Maule Valley (Chile). S. Afr. J. Enol. Vitic. 2019, 40, 181–191. [Google Scholar] [CrossRef]
- Yuan, Z.; Cao, Q.; Zhang, K.; Ata-Ul-Karim, S.T.; Tan, Y.; Zhu, Y.; Cao, W.; Liu, X. Optimal Leaf Positions for SPAD Meter Measurement in Rice. Front. Plant Sci. 2016, 7, 719. [Google Scholar] [CrossRef] [Green Version]
- Chen, C. Determining the Leaf Emissivity of Three Crops by Infrared Thermometry. Sensors 2015, 15, 11401. [Google Scholar] [CrossRef] [Green Version]
- Kacar, B.; Inal, A. Plant Analysis; Nobel Press: Ankara, Turkey, 2008. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Panneerselvam, R. Calcium Chloride Effects on Salinity-Induced Oxidative Stress, Proline Metabolism and Indole Alkaloid Accumulation in Catharanthus Roseus. Comptes Rendus Biol. 2007, 330, 674–683. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, W.U.; Ali Shah, A.; Yasin, N.A.; Naz, S.; Ali, A.; Tahir, A.; Iram Batool, A. Synergistic Effects of Nitric Oxide and Silicon on Promoting Plant Growth, Oxidative Stress Tolerance and Reduction of Arsenic Uptake in Brassica Juncea. Chemosphere 2021, 262, 128384. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Rai, P.; Guerriero, G.; Sharma, S.; Corpas, F.J.; Singh, V.P. Silicon Induces Adventitious Root Formation in Rice under Arsenate Stress with Involvement of Nitric Oxide and Indole-3-Acetic Acid. J. Exp. Bot. 2021, 72, 4457–4471. [Google Scholar] [CrossRef]
- Hamidou, F.; Zombre, G.; Braconnier, S. Physiological and Biochemical Responses of Cowpea Genotypes to Water Stress under Glasshouse and Field Conditions. J. Agron. Crop Sci. 2007, 193, 229–237. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Giannakoula, A.; Ilias, I.; Zamanidis, P. Alleviation of Drought and Salinity Stresses on Growth, Physiology, Biochemistry and Quality of Two Cucumis Sativus L. Cultivars by Si Application. Rev. Bras. Bot. 2016, 39, 531–539. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K. The Regulatory Role of Silicon on Water Relations, Photosynthetic Gas Exchange, and Carboxylation Activities of Wheat Leaves in Field Drought Conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A Review of the Use of Biostimulants in the Vineyard for Improved Grape and Wine Quality: Effects on Prevention of Grapevine Diseases. J. Sci. Food Agric. 2019, 99, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Alañón-Sánchez, N.; Mateluna-Cuadra, R.; Verdugo-Vásquez, N. An Overview about the Impacts of Agricultural Practices on Grape Nitrogen Composition: Current Research Approaches. Food Res. Int. 2020, 136, 109477. [Google Scholar] [CrossRef]
- Ismail, G.S.M. Protective Role of Nitric Oxide against Arsenic-Induced Damages in Germinating Mung Bean Seeds. Acta Physiol. Plant. 2012, 34, 1303–1311. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Sharma, V.P.; Sharma, N.; Kohli, R.K. Nitric Oxide Alleviates Arsenic Toxicity by Reducing Oxidative Damage in the Roots of Oryza Sativa (Rice). Nitric Oxide 2009, 20, 289–297. [Google Scholar] [CrossRef]
- Moussa, H.R. Influence of Exogenous Application of Silicon on Physiological Response of Salt-Stressed Maize (Zea Mays L.). Int. J. Agric. Biol. 2006, 8, 293–297. [Google Scholar]
- Agarie, S. Physiological Roles of Silicon in Photosynthesis and Dry Matter Production in Rice Plants I. Effects of Silicon and Shading Treatments. Jpn. J. Crop Sci. 1992, 61, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Verma, K.K.; Singh, P.; Song, X.P.; Malviya, M.K.; Singh, R.K.; Chen, G.L.; Solomon, S.; Li, Y.R. Mitigating Climate Change for Sugarcane Improvement: Role of Silicon in Alleviating Abiotic Stresses. Sugar Tech 2020, 22, 741–749. [Google Scholar] [CrossRef]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Kobayashi, S.; Kawamura, Y.; Tanaka, K.; Inanaga, S. Effect of Silicon Application on Sorghum Root Responses to Water Stress. J. Plant Nutr. 2010, 34, 71–82. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial Effects of Silicon on Salt and Drought Tolerance in Plants. Agron. Sustain. Dev. 2013, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Ju, S.; Wang, L.; Yin, N.; Li, D.; Wang, Y.; Zhang, C. Silicon Alleviates Simulated Acid Rain Stress of Oryza Sativa L. Seedlings by Adjusting Physiology Activity and Mineral Nutrients. Protoplasma 2017, 254, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.; Kapoor, D. Fascinating Regulatory Mechanism of Silicon for Alleviating Drought Stress in Plants. Plant Physiol. Biochem. 2021, 166, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Devasagayam, T.P.A.; Bhushan, B.; Verma, N.C. Scavenging of Reactive Oxygen Species by Chlorophyllin: An ESR Study. Free Radic. Res. 2009, 35, 563–574. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Zhang, J.; He, Y.; Zhu, X.; Zhou, X.; Gong, H.; Yin, J.; Liu, Y. Silicon Confers Cucumber Resistance to Salinity Stress through Regulation of Proline and Cytokinins. Plant Physiol. Biochem. 2020, 156, 209–220. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of Leaf Stomatal Density to Water Status and Its Relationship with Photosynthesis in a Grass. J. Exp. Bot. 2008, 59, 3325. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.X.; Xu, X.B.; Hu, Y.H.; Han, W.H.; Yin, J.L.; Li, H.L.; Gong, H.J. Silicon Improves Salt Tolerance by Increasing Root Water Uptake in Cucumis Sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Irrigation Regime | Chemicals Applications | N (%) | P (%) | K (%) | Ca (%) | Mg (%) | Fe (ppm) | Zn (ppm) |
|---|---|---|---|---|---|---|---|---|
| Non-water-stress | Control | 2.80 bc | 0.19 b | 2.8 | 1.5 c | 0.41 c | 75.5 d | 27.4 c |
| Si | 2.89 b | 0.19 b | 2.9 | 1.6 b | 0.41 c | 77.2 c | 28.4 b | |
| NO | 2.86 b | 0.19 b | 2.7 | 1.6 b | 0.79 a | 83.2 b | 28.5 b | |
| Si+NO | 3.54 a | 0.22 a | 3.2 | 1.7 a | 0.84 a | 94.7 a | 29.4 a | |
| Water stress | Control | 2.50 d | 0.15 c | 2.1 | 1.1 g | 0.20 d | 65.2 h | 22.5 g |
| Si | 2.59 cd | 0.16 c | 2.2 | 1.2 f | 0.25 d | 65.3 g | 23.3 f | |
| NO | 2.65 bcd | 0.15 c | 2.3 | 1.3 e | 0.43 c | 70.5 f | 24.4 e | |
| Si+NO | 2.72 bcd | 0.16 c | 2.4 | 1.4 d | 0.54 b | 72.1 e | 25.1 d | |
| Irrigation regime | ** | ** | ** | ** | ** | ** | ** | |
| Chemicals applications | ** | ** | ** | ** | ** | ** | ** | |
| Interaction | ** | ** | ns | ** | ** | ** | ** | |
| Irrigation Regime | Chemicals Applications | N (%) | P (%) | K (%) | Ca (%) | Mg (%) | Fe (ppm) | Zn (ppm) |
|---|---|---|---|---|---|---|---|---|
| Non-water-stress | Control | 1.63 c | 0.19 c | 0.91 c | 1.7 bc | 0.16 c | 12.3 | 1.0 c |
| Si | 1.83 b | 0.22 a | 1.05 ab | 1.9 ab | 0.17 bc | 14.0 | 1.3 b | |
| NO | 1.79 b | 0.20 b | 1.01 b | 1.8 b | 0.19 b | 14.2 | 1.4 a | |
| Si+NO | 2.38 a | 0.20 b | 1.08 a | 2.0 a | 0.22 a | 15.2 | 1.4 a | |
| Water stress | Control | 1.30 f | 0.16 e | 0.46 e | 1.1 f | 0.13 d | 5.0 | 0.6 f |
| Si | 1.46 de | 0.18 cd | 0.75 d | 1.4 e | 0.15 cd | 5.8 | 0.7 e | |
| NO | 1.40 ef | 0.17 d | 0.76 d | 1.5 de | 0.17 bc | 5.4 | 0.8 d | |
| Si+NO | 1.60 cd | 0.18 cd | 0.78 d | 1.6 d | 0.17 bc | 6.7 | 0.8 d | |
| Irrigation regime | ** | ** | ** | ** | ** | * | ** | |
| Chemical applications | ** | ** | ** | ** | ** | * | ** | |
| Interaction | ** | ** | ** | ** | ** | ns | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolat, I.; Bakır, A.G.; Korkmaz, K.; Gutiérrez-Gamboa, G.; Kaya, O. Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.). Agriculture 2022, 12, 1273. https://doi.org/10.3390/agriculture12081273
Bolat I, Bakır AG, Korkmaz K, Gutiérrez-Gamboa G, Kaya O. Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.). Agriculture. 2022; 12(8):1273. https://doi.org/10.3390/agriculture12081273
Chicago/Turabian StyleBolat, Ibrahim, Asuman Gundogdu Bakır, Kubra Korkmaz, Gastón Gutiérrez-Gamboa, and Ozkan Kaya. 2022. "Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.)" Agriculture 12, no. 8: 1273. https://doi.org/10.3390/agriculture12081273
APA StyleBolat, I., Bakır, A. G., Korkmaz, K., Gutiérrez-Gamboa, G., & Kaya, O. (2022). Silicon and Nitric Oxide Applications Allow Mitigation of Water Stress in Myrobalan 29C Rootstocks (Prunus cerasifera Ehrh.). Agriculture, 12(8), 1273. https://doi.org/10.3390/agriculture12081273