Can Modification of Sowing Date and Genotype Selection Reduce the Impact of Climate Change on Sunflower Seed Production?

 ,
,  , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
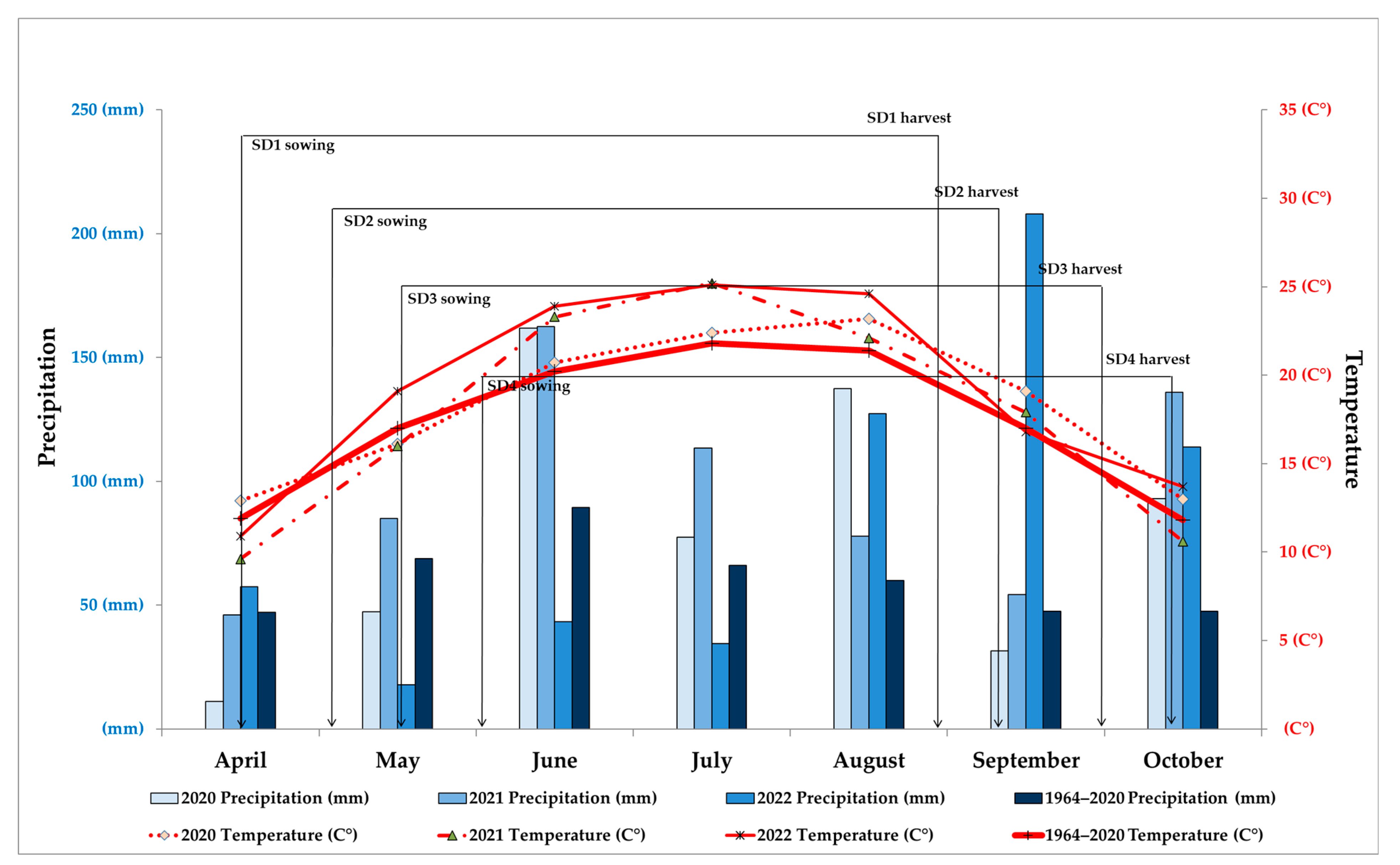
2.2. Growing Season Conditions
2.3. Observed Parameters of Sunflower Plants
2.4. Statistical Analysis
3. Results and Discussion
3.1. Factor Share in the Variation of All Examined Traits
3.2. The Influence of the Sowing Date on the Observed Traits of Sunflower Plants through Different Growing Seasons
3.3. The Behavior and Influence of Different Genotypes of Sunflower on the Observed Traits during Different Vegetation Seasons
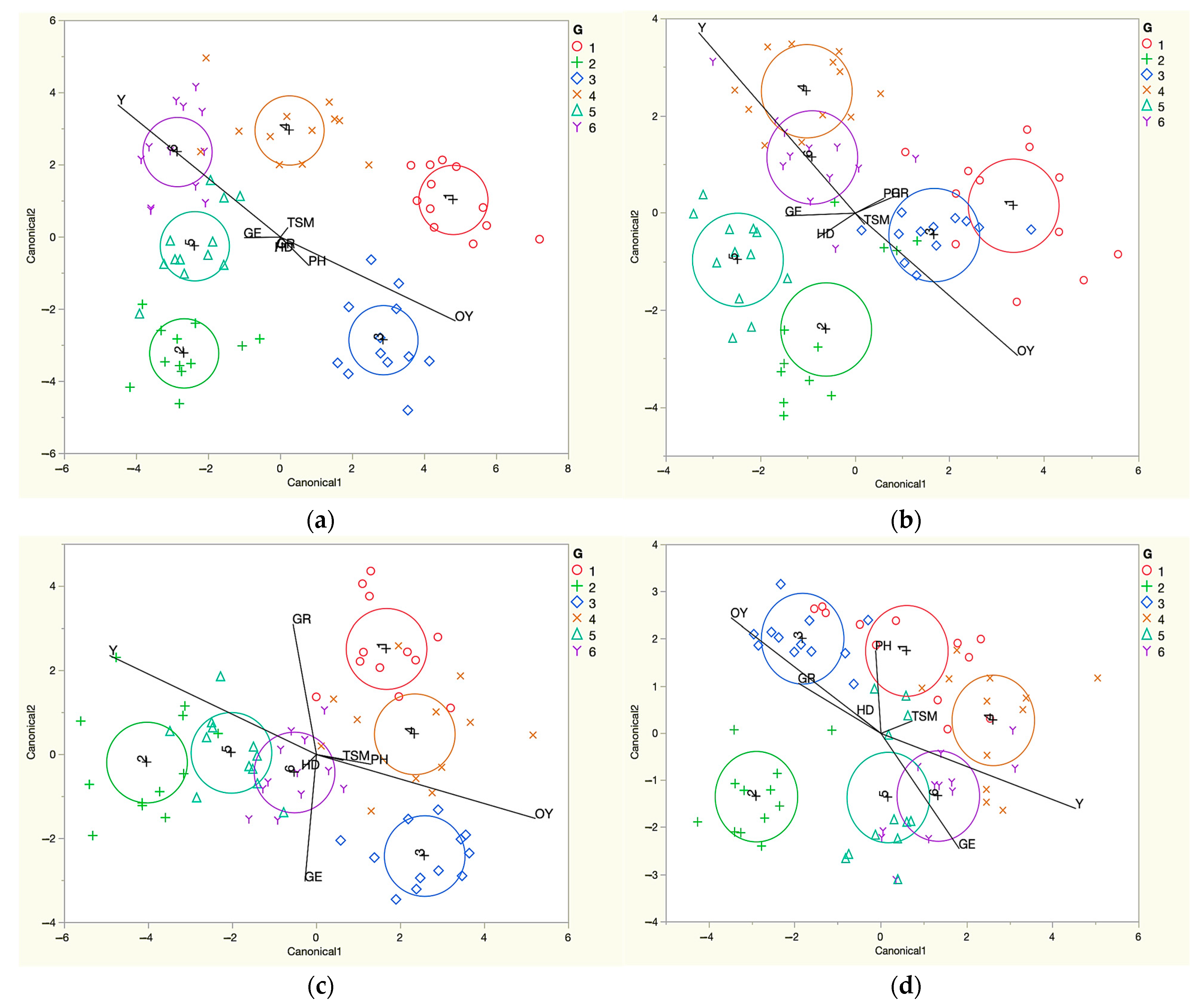
3.4. Interpretation of the Interaction between Genotype and Sowing Date through Linear Discriminant Analysis (LDA)
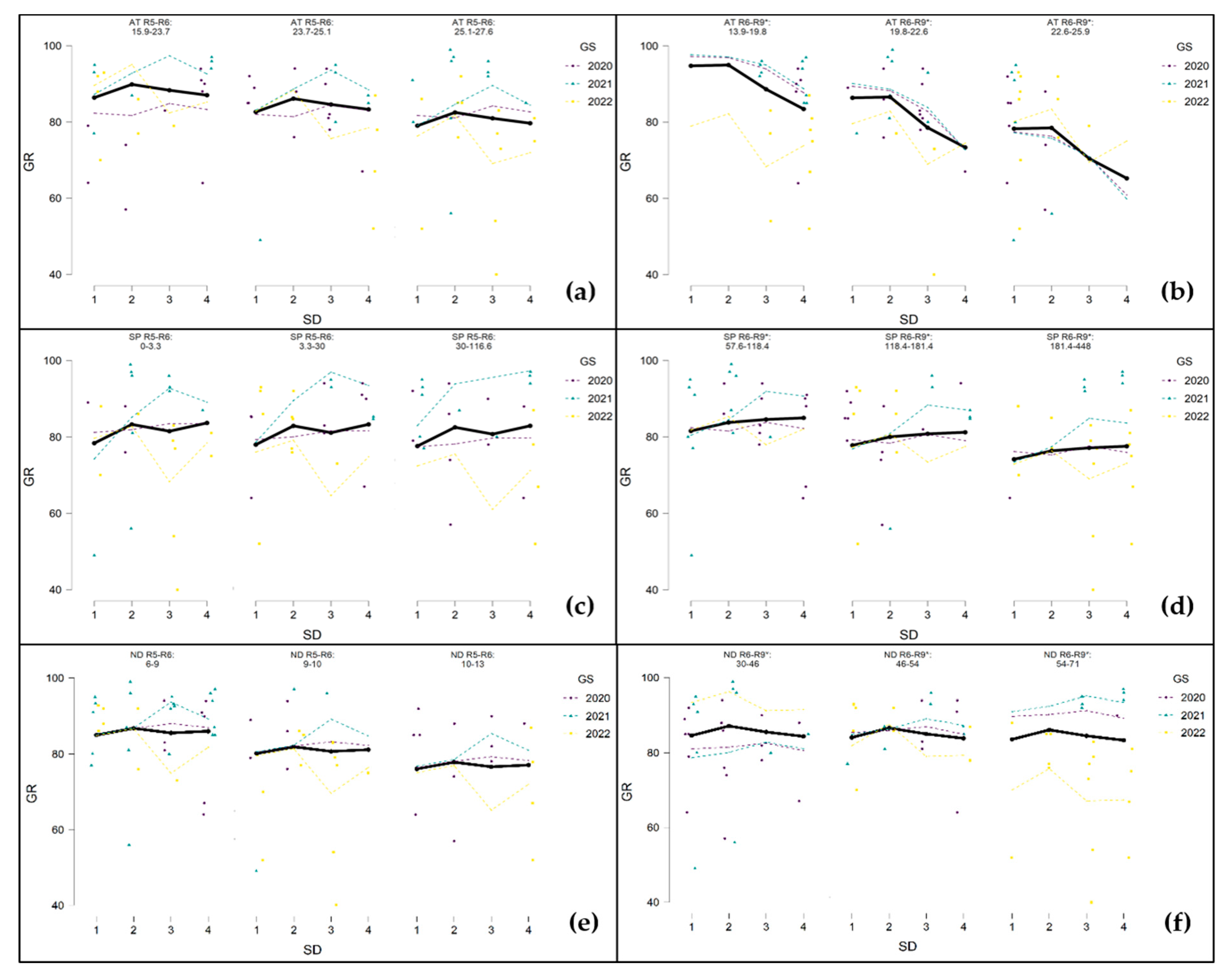
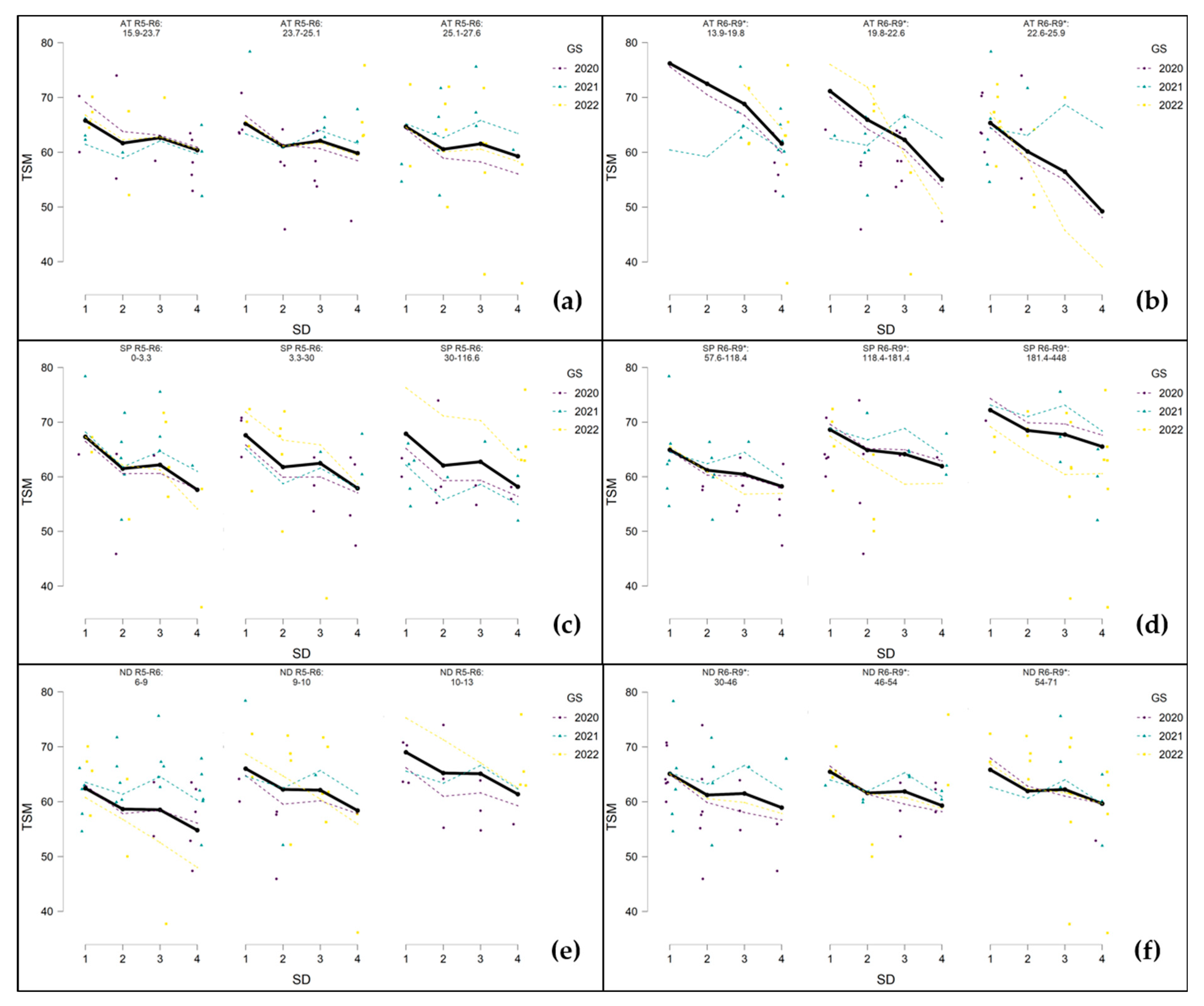
3.5. The Influence of Weather Conditions during Critical Growth Phases of Sunflowers on Measured Parameters of Seed Production Depending on Sowing Dates
3.5.1. Germination (GR)
3.5.2. 1000-Seed Mass (TSM)
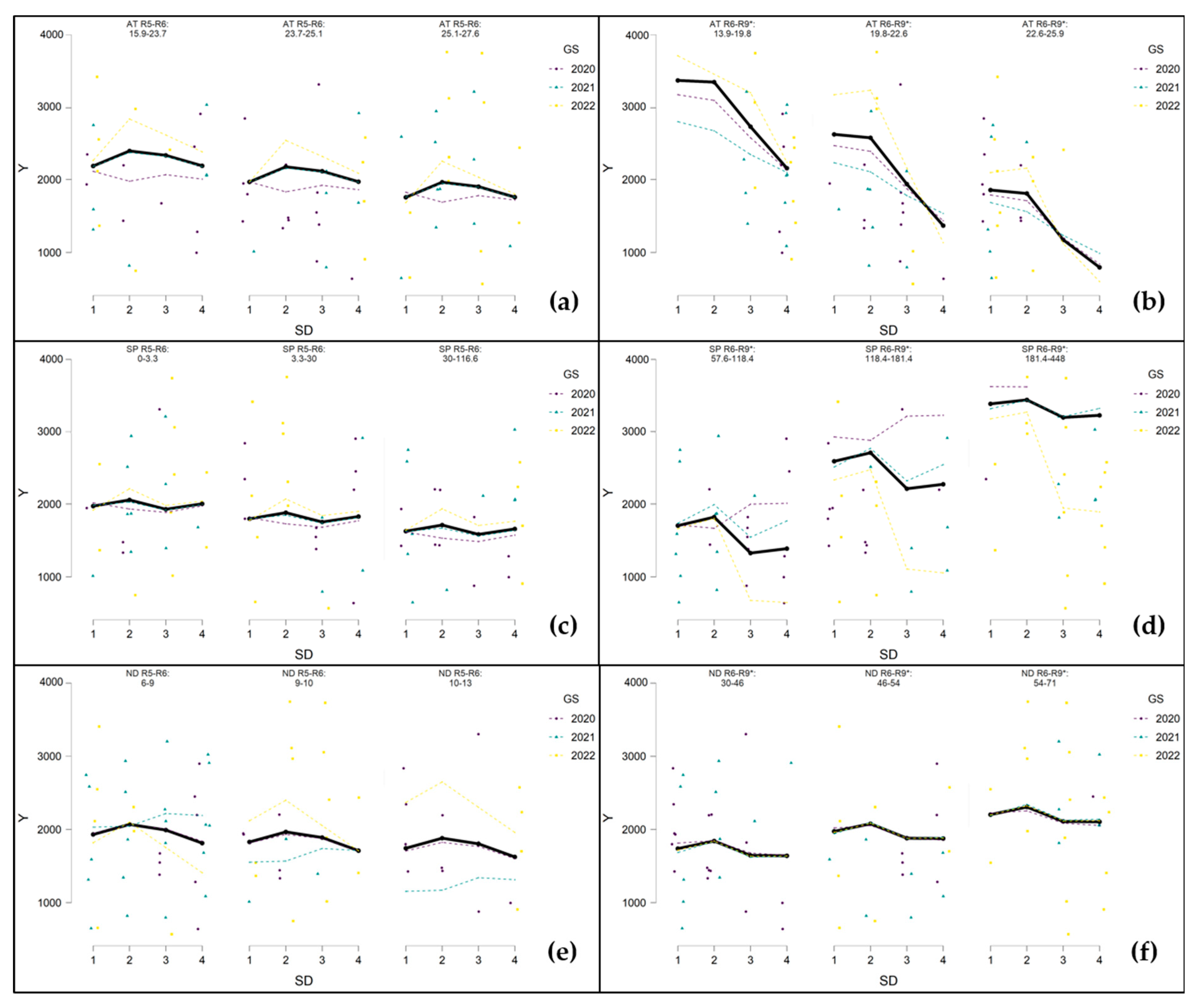
3.5.3. Seed Yield (Y)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mimić, G.; Živaljević, B.; Blagojević, D.; Pejak, B.; Brdar, S. Quantifying the effects of drought using the crop moisture stress as an indicator of maize and sunflower yield reduction in Serbia. Atmosphere 2022, 13, 1880. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Gomes, S.E.V.; Gomes, R.A.; Dantas, B.F. Climate and seed size of a dry forest species: Influence on seed production, physiological quality, and tolerance to abiotic stresses. J. Seed Sci. 2023, 45, e202345013. [Google Scholar] [CrossRef]
- Noroozi, N.; Mohammadi, G.; Ghobadi, M. Some physio-biochemical traits of sunflower (Helianthus annuus L.) as affected by arbuscular mycorrhizal fungi inoculation under different irrigation treatments. Ital. J. Agron. 2023, 18, 2033. [Google Scholar] [CrossRef]
- Tariq, M.; Ahmad, S.; Fahad, S.; Abbas, G.; Hussain, S.; Fatima, Z.; Nasim, W.; Mubeen, M.; Habib ur Rehman, M.; Khan, M.A.; et al. The impact of climate warming and crop management on phenology of sunflower-based cropping systems in Punjab, Pakistan. Agric. For. Meteorol. 2018, 256–257, 270–282. [Google Scholar] [CrossRef]
- Miladinović, D.; Hladni, N.; Radanović, A.; Jocić, S.; Cvejić, S. Sunflower and climate change: Possibilities of adaptation through breeding and genomic selection. In Genomic Designing of Climate-Smart Oilseed Crops; Kole, C., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 173–238. [Google Scholar]
- Çetin, N.; Karaman, K.; Beyzi, E.; Sağlam, C.; Demirel, B. Comparative evaluation of some quality characteristics of sunflower oilseeds (Helianthus annuus L.) through machine learning classifiers. Food Anal. Methods 2021, 14, 1666–1681. [Google Scholar] [CrossRef]
- Badouin, H.; Gouzy, J.; Grassa, C.J.; Murat, F.; Staton, S.E.; Cottret, L.; Lelandais-Brière, C.; Owens, G.L.; Carrère, S.; Mayjonade, B.; et al. The sunflower genome provides insights into oil metabolism, flowering and asterid evolution. Nature 2017, 546, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Radanović, A.; Miladinović, D.; Cvejić, S.; Jocković, M.; Jocić, S. Sunflower genetics from ancestors to modern hybrids—A review. Genes 2018, 9, 528. [Google Scholar] [CrossRef]
- Debaeke, P.; Casadebaig, P.; Langlade, N. New challenges for sunflower ideotyping in changing environments and more ecological cropping systems. OCL 2021, 28, 29. [Google Scholar] [CrossRef]
- Babec, B.; Šeremešić, S.; Hladni, N.; Ćuk, N.; Stanisavljević, D.; Rajković, M. Potential of sunflower—Legume intercropping: A way forward in sustainable production of sunflower in temperate climatic conditions. Agronomy 2021, 11, 2381. [Google Scholar] [CrossRef]
- Donatelli, M.; Srivastava, A.K.; Duveiller, G.; Niemeyer, S.; Fumagalli, D. Climate change impact and potential adaptation strategies under alternate realizations of climate scenarios for three major crops in Europe. Environ. Res. Lett. 2015, 10, 075005. [Google Scholar] [CrossRef]
- Dimov, Z.; Cukaliev, O.; Mukaetov, D.; Tanaskovic, V. Sunflower and climate changes: Adaptation and mitigation potential from case study in RN Macedonia. In Proceedings of the 20th International Sunflower Conference, Novi Sad, Serbia, 20–23 June 2022. [Google Scholar]
- De Estrada, E.; Vázquez, M.; Moreno, D.; Bravo, S.; Amores, J.; San Román, G.; Dodds, J.; Bertero de Romano, A.; Bergada, P.; Sala, C. Sunflower seed production: Past, present, and perspectives. In Proceedings of the 18th International Sunflower Conference, Mar del Plata, Argentina, 27 February–1 March 2012. [Google Scholar]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- El-Bially, M.E.; Saudy, H.S.; El-Metwally, I.M.; Shahin, M.G. Sunflower response to application of L-ascorbate under thermal stress associated with different sowing dates. Gesunde Pflanz. 2022, 74, 87–96. [Google Scholar] [CrossRef]
- Balalić, I.; Zorić, M.; Branković, G.; Terzić, S.; Crnobarac, J. Interpretation of hybrid× sowing date interaction for oil content and oil yield in sunflower. Field Crops Res. 2012, 137, 70–77. [Google Scholar] [CrossRef]
- Giannini, V.; Mula, L.; Carta, M.; Patteri, G.; Roggero, P.P. Interplay of irrigation strategies and sowing dates on sunflower yield in semi-arid Mediterranean areas. Agric. Water Manag. 2022, 260, 107287. [Google Scholar] [CrossRef]
- Paul, P.L.C.; Bell, R.W.; Barrett-Lennard, E.G.; Kabir, E.; Gaydon, D.S. Opportunities and risks with early sowing of sunflower in a salt-affected coastal region of the Ganges Delta. Agron. Sustain. Dev. 2021, 41, 39. [Google Scholar] [CrossRef]
- Hussain, F.; Rafiq, M.; Ghias, M.; Qamar, R.; Razzaq, M.K.; Hameed, A.; Habib, S.; Mustafa, H.S. Genetic diversity for seed yield and its components using principal component and cluster analysis in sunflower r (Helianthus annuus L.). Life Sci. J. 2017, 14, 71–78. [Google Scholar] [CrossRef]
- Markulj-Kulundžić, A.; Sudarić, A.; Matoša-Kočar, M.; Mijić, A.; Liović, I.; Viljevac-Vuletić, M.; Varga, I.; Cesar, V.; Lepeduš, H. Sunflower agronomic traits in field irrigation conditions. Genetika 2022, 54, 473–489. [Google Scholar] [CrossRef]
- Mrđa, J.; Crnobarac, J.; Radić, V.; Miklič, V. Sunflower seed quality and yield in relation to environmental conditions of production region. Helia 2012, 35, 123–134. [Google Scholar] [CrossRef]
- Kalenska, S.; Ryzhenko, A.; Novytska, N.; Garbar, L.; Stolyarchuk, T.; Kalenskyi, V.; Shytiy, O. Morphological features of plants and yield of sunflower hybrids cultivated in the northern part of the forest-steppe of Ukraine. Am. J. Plant Sci. 2020, 11, 1331–1344. [Google Scholar] [CrossRef]
- Duca, M.; Port, A.; Burcovschi, I.; Joiţa-Păcureanu, M.; Dan, M. Environmental response in sunflower hybrids: A multivariate approach. Rom. Agric. Res. 2022, 39, 1–14. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, M.; Zaman, J.; Paul, S.K.; Rahman, M.M.; Islam, M.; Majumdar, F. Effect of sowing dates on the yield of sunflower. Bangladesh Agron. J. 2015, 18, 1–5. [Google Scholar] [CrossRef]
- Debaeke, P.; Casadebaig, P.; Flenet, F.; Langlade, N. Sunflower crop and climate change: Vulnerability, adaptation, and mitigation potential from case-studies in Europe. OCL 2017, 24, D102. [Google Scholar] [CrossRef]
- Ćuk, N.; Cvejić, S.; Mladenov, V.; Jocković, M.; Babec, B.; Miklič, V.; Jocić, S. Variability of agronomic traits in sunflower inbred lines. Sel. I Semen. 2020, 26, 29–37. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015 International soil classification system for naming soils and creating legends for soil maps. In World Soil Resources Reports; FAO: Rome, Italy, 2015; Volume 106, pp. 1–203. [Google Scholar]
- Republic Hydrometeorological Service of Serbia. Available online: https://www.hidmet.gov.rs/ciril/meteorologija/agro.php (accessed on 10 May 2023).
- Schneiter, A.A.; Miller, J.F. Description of sunflower growth stages. Crop Sci. 1981, 21, 901–903. [Google Scholar] [CrossRef]
- ISTA Rules; International Seed Testing Association: Zurich, Switzerland, 2018.
- Granlund, M.; Zimmerman, D.C. Effect of drying conditions on oil contents of sunflower (Helianthus annuus L.) seed determined by wide-line Nuclear Magnetic Resonance (NMR). Proc. N. Dak. Acad. Sci. 1975, 27, 128–132. [Google Scholar]
- Milligan, G.W.; Cooper, M.C. An examination of procedures for determining the number of clusters in a data set. Psychometrika 1985, 50, 159–179. [Google Scholar] [CrossRef]
- Tharwat, A.; Gaber, T.; Ibrahim, A.; Hassanien, A.E. Linear discriminant analysis: A detailed tutorial. AI Commun. 2017, 30, 169–190. [Google Scholar] [CrossRef]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Jaunet, T.; Chi-Ham, C.; Cantu, D.; Bradford, K.J.; Van Deynze, A. Texture diversity in melon (Cucumis melo L.): Sensory and physical assessments. Postharvest Biol. Technol. 2020, 159, 111024. [Google Scholar] [CrossRef]
- IBM Corp. Released. IBM SPSS Statistics for Windows; Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- SAS Institute Inc. JMP® 13 Predictive and Specialized Modeling, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Love, J.; Selker, R.; Marsman, M.; Jamil, T.; Dropmann, D.; Verhagen, J.; Ly, A.; Gronau, Q.F.; Šmíra, M.; Epskamp, S.; et al. JASP: Graphical statistical software for common statistical designs. J. Stat. Softw. 2019, 88, 1–17. [Google Scholar] [CrossRef]
- Faramarzi, A.; Khorshidi, M.B. Planting date effect on yield and yield components of sunflower in Mizaneh region. In Proceedings of the 17th International Sunflower Conference, Cordoba, Spain, 8–12 June 2008. [Google Scholar]
- Partal, E. Sunflower yield and quality under the influence of sowing date, plant population and the hybrid. Rom. Agric. Res. 2022, 39, 463–470. [Google Scholar] [CrossRef]
- Babec, B. Intercropping In the Service of Improving Soil Quality and Biological and Production Characteristics of Sunflower. Ph.D. Thesis, Faculty of Agriculture, University of Novi Sad, Novi Sad, Serbia, 2023. Available online: https://fiver.ifvcns.rs/handle/123456789/3454 (accessed on 11 September 2023).
- Krstić, M.; Ovuka, J.; Mladenov, V.; Radić, V.; Krstić, J.; Ćuk, N.; Miklič, V. Interdependence of the oil content in pure seed and other tested traits of inbred sunflower lines. Uljarstvo 2021, 52, 5–12. [Google Scholar]
- Ion, V.; Dicu, G.; Basa, A.G.; Dumbrava, M.; Temocico, G.; Epure, L.I.; State, D. Sunflower yield and yield components under different sowing conditions. Agric. Agric. Sci. Procedia 2015, 6, 44–51. [Google Scholar] [CrossRef]
- Kvashin, A.A.; Neshchadim, N.N.; Yablonskay, E.K.; Gorpinchenko, K.N. Crop yield and the quality of sunflower seeds in the use of fertilizers and growth regulation substances. Helia 2018, 41, 227–239. [Google Scholar] [CrossRef]
- Radić, V.; Balalić, I.; Miladinov, Z.; Ćirić, M.; Vasiljević, M.; Jocić, S.; Marjanović-Jeromela, A. Genotype x environment interaction of some traits in sunflower (Helianthus annuus L.) lines. Appl. Ecol. Environ. Res. 2020, 18, 1707–1719. [Google Scholar] [CrossRef]
- Crnobarac, J.; Balalić, I.; Latković, D.; Jaćimović, G. The influence of sowing date on yield and quality of NS sunflower hybrids. In Proceedings of the 20th International Sunflower Conference, Novi Sad, Serbia, 20–23 June 2022. [Google Scholar]
- Chambo, E.D.; Escocard de Oliveira, N.T.; Garcia, R.C.; RuvoloTakasusuki, M.C.C.; Arnaut de Toledo, V.A. Phenotypic correlation and path analysis in sunflower genotypes and pollination influence on estimates. Open Biol. Sci. J. 2017, 3, 9–15. [Google Scholar] [CrossRef]
- Ćuk, N.; Cvejić, S.; Mladenov, V.; Babec, B.; Miklič, V.; Miladinović, D.; Jocić, S. Variability of morphological traits in sunflower inbred lines. Genetika 2020, 52, 911–923. [Google Scholar] [CrossRef]
- Radić, V.; Balalić, I.; Jaćimović, G.; Krstić, M.; Jocković, M.; Jocić, S. A study of correlations and path analyses of some parameters in sunflower parental lines. Ratar. I Povrt. 2021, 58, 7–13. [Google Scholar] [CrossRef]
- Creux, N.; Marshall, C.; Memela, U.; Mapfumo, P.; Harmer, S. The delicate balancing act of climate control during flowering, pollination and seed development in sunflower. In Proceedings of the 20th International Sunflower Conference, Novi Sad, Serbia, 20–23 June 2022. [Google Scholar]
- Oshundiya, F.; Olowe, V.I.; Sowemimo, F.; Odedina, J.N. Seed yield and quality of sunflower (Helianthus annuus L.) as influenced by staggered sowing and organic fertilizer application in the humid tropics. Helia 2014, 37, 237–255. [Google Scholar] [CrossRef]
- Ozturk, E.; Polat, T.; Sezek, M. The effect of sowing date and nitrogen fertilizer form on growth, yield and yield components in sunflower. Turk. J. Field Crops 2017, 22, 143–151. [Google Scholar] [CrossRef]
- Balalić, I. Multivariate Analysis of Interaction between Hybrids and Planting Dates for Oil Content, Yield and Yield Components in Sunflower. Ph.D. Thesis, Faculty of Agriculture, University of Novi Sad, Novi Sad, Serbia, 2010. [Google Scholar]
- Abbas, G.; Ahmad, S.; Ahmad, A.; Nasim, W.; Fatima, Z.; Hussain, S.; Habib ur Rehman, M.; Khan, M.A.; Hasanuzzaman, M.; Fahad, S.; et al. Quantification the impacts of climate change and crop management on phenology of maize-based cropping system in Punjab, Pakistan. Agric. For. Meteorol. 2017, 247, 42–55. [Google Scholar] [CrossRef]
- Rezadust, S.; Mokhtarzadeh, S. Effect of planting date on germination indices of Erflor cultivar of sunflower seeds. Turk. J. Agric. Nat. Sci. 2019, 6, 792–797. [Google Scholar] [CrossRef]
- Škorić, D. Sunflower breeding for resistance to abiotic and biotic stresses. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A., Shanker, C., Eds.; InTech: Rijeka, Croatia, 2016; Volume 25. [Google Scholar]
- Krstić, M.; Babec, B.; Hladni, N.; Klisurić, N.; Radeka, I.; Ostojić, B.; Butaš, D.; Jokić, G.; Ovuka, J.; Miklič, V.; et al. Potencijal NS hibrida suncokreta u uslovima promenjene klime. In Proceedings of the 57. Savetovanje Agronoma i Poljoprivrednika Srbije (SAPS) i 3. Savetovanje Agronoma Republike Srbije i Republike Srpske, Zlatibor, Serbia, 30 January–3 February 2023. [Google Scholar]
- Alqudah, A.M.; Samarah, N.H.; Mullen, R.E. Drought stress effect on crop pollination, seed set, yield and quality. In Alternative Farming Systems, Biotechnology, Drought Stress and Ecological Fertilisation. Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2011; Volume 6, pp. 193–213. [Google Scholar] [CrossRef]
- Soriano, M.A.; Orgaz, F.; Villalobos, F.J.; Fereres, E. Efficiency of water use of early plantings in sunflower. Eur. J. Agron. 2004, 21, 465–476. [Google Scholar] [CrossRef]
- Šućur, R.; Mladenov, V.; Banjac, B.; Trkulja, D.; Mikić, S.; Šumaruna, M.; Börner, A. Phenotypic marker study of worldwide wheat germplasm. Ital. J. Agron. 2023; early access. [Google Scholar] [CrossRef]
- Ćirić, M.; Jocić, S.; Cvejić, S.; Čanak, P.; Jocković, M.; Marinković, R.; Mirosavljević, M. Evaluation of combining abilities of new sunflower inbred lines. Ratar. I Povrt. 2013, 50, 8–15. [Google Scholar] [CrossRef]
- Babec, B.; Šeremešić, S.; Hladni, N.; Terzić, S.; Vojnov, B.; Ćuk, N.; Gvozdenac, S. Effect of intercropping sunflower with legumes on some sunflower morphological traits. Ratar. I Povrt. 2020, 57, 61–67. [Google Scholar] [CrossRef]
- Dar, J.S.; Cheema, M.A.; Rehmani, M.I.A.; Khuhro, S.; Rajput, S.; Virk, A.L.; Hussain, S.; Bashir, M.A.; Alghanem, S.M.; Al-Zuaibr, F.M.; et al. Potassium fertilization improves growth, yield and seed quality of sunflower (Helianthus annuus L.) under drought stress at different growth stages. PLoS ONE 2021, 16, e0256075. [Google Scholar] [CrossRef] [PubMed]
- Mrđa, J.; Crnobarac, J.; Dušanić, N.; Jocić, S.; Miklič, V. Germination energy as a parameter of seed quality in different sunflower genotypes. Genetika 2011, 43, 427–436. [Google Scholar] [CrossRef]
- Krstić, M.; Ovuka, J.; Gvozdenac, S.; Butaš, D.; Ćuk, N.; Babec, B.; Cvejić, S. Seed vitality of sunflower inbred lines as influenced by climatic factors and seed size. J. Process. Energy Agric. 2022, 26, 106–110. [Google Scholar] [CrossRef]
- Baraiya, V.K.; Jagtap, P.K.; Sangani, J.L.; Malviya, A.V. Correlation and path analysis in sunflower (Helianthus annus L.). J. Pharmacogn. Phytochem. 2018, 7, 2730–2732. [Google Scholar]
- Babić, V.; Babić, M. Phenotypic characterization and multivariate analyses. Proc. XXII Conf. Agron. Vet. Technol. 2008, 14, 71–79. [Google Scholar]
- Krstić, M.; Mladenov, V.; Ćuk, N.; Ovuka, J.; Gvozdenac, S.; Krstić, J.; Miklič, V. Agro-morphological traits of inbred sunflower lines and their genetic assessment. Contemp. Agric. 2022, 71, 87–95. [Google Scholar] [CrossRef]
- Bushnev, A.S.; Demurin, Y.N.; Orekhov, G.I. Productivity of sunflower hybrids with erectoid leaves at various plant densities. OCL 2021, 28, 39. [Google Scholar] [CrossRef]
- Gul, R.M.S.; Sajid, M.; Rauf, S.; Munir, H.; Shehzad, M.; Haider, W. Evaluation of drought-tolerant sunflower (Helianthus annuus L.) hybrids in autumn and spring planting under semi-arid rainfed conditions. OCL 2021, 28, 24. [Google Scholar] [CrossRef]
- Rezaie, B.; Hosseinpanahi, F.; Siosemardeh, A.; Darand, M.; Bannayan, M. Shifting the sowing date of winter wheat as a strategy for adaptation to climate change in a Mediterranean-type environment. Int. J. Plant Prod. 2022, 16, 595–610. [Google Scholar] [CrossRef]
- Hussain, M.M.; Rauf, S.; Noor, M.; Bibi, A.; Ortiz, R.; Dahlberg, J. Evaluation of introgressed lines of sunflower (Helianthus annuus L.) under contrasting water treatments. Agriculture 2023, 13, 1250. [Google Scholar] [CrossRef]
- Riveira Rubin, M.; Arata, G.J.; López, E.D.; Rodriguez, M.V.; Batlla, D. Dormancy attributes in sunflower achenes (Helianthus annuus L.): II. Sowing date effects. Crop Sci. 2021, 61, 4309–4324. [Google Scholar] [CrossRef]
- Berton, T.; Bernillon, S.; Fernandez, O. Leaf metabolomic data of eight sunflower lines and their sixteen hybrids under water deficit. OCL 2021, 28, 42. [Google Scholar] [CrossRef]
- Aguirrezábal, L.A.N.; Lavaud, Y.; Dossio, G.A.A.; Izquierdo, N.G.; Andrade, F.H.; Gonzales, L.M. Intercepted solar radiation during seed filling determines sunflower weight per seed and oil concentration. Crop Sci. 2003, 43, 152–161. [Google Scholar] [CrossRef]
- Nazarli, H.; Zardashti, M.R.; Darvishzadeh, R.; Najafi, S. The effect of water stress and polymer on water use efficiency, yield and several morphological traits of sunflower under greenhouse condition. Not. Sci. Biol. 2010, 2, 53–58. [Google Scholar] [CrossRef]
- Farzad, B.A.; Mahmoud, T.; Majid, N.; Mohamad-Reza, S. Effect of drought stress on yield and yield components of some sunflower recombinant inbred lines. Int. J. Biosci. 2013, 3, 50–56. [Google Scholar] [CrossRef]
- Angeloni, P.; Aguirrezábal, L.; Echarte, M.M. Assessing the mechanisms underlying sunflower grain weight and oil content responses to temperature during grain filling. Field Crops Res. 2021, 262, 108040. [Google Scholar] [CrossRef]
- Forrest, J.; Miller-Rushing, A.J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos. Trans. R. Soc. B 2010, 365, 3101–3112. [Google Scholar] [CrossRef] [PubMed]
- Nasim, W.; Ahmad, A.; Wajid, A.; Akhtar, J.; Muhammad, D. Nitrogen effects on growth and development of sunflower hybrids under agro-climatic conditions of Multan. Pak. J. Bot. 2011, 43, 2083–2092. [Google Scholar]
- Kaya, Y. Sunflower. In Breeding Oil Seed Crops for Sustainable Production. Opportunities and Constraints; Gupta, S.K., Ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 4, pp. 55–88. [Google Scholar]
- Miklič, V.; Balalić, I.; Jocić, S.; Cvejić, S.; Jocković, M.; Miladinović, D.; Terzić, S. NS hibridi suncokreta u ogledima u Bačkoj i Banatu 2019. godine. In Proceedings of the 61. Savetovanje industrije ulja, Herceg Novi, Montenegro, 28 June–3 July 2020. [Google Scholar]
- Kalarani, M.K.; Senthil, A.; Thangaraj, M. Effect of water stress on morpho-physiological traits of sunflower (Helianthus annuus L.) genotypes. Madras Agric. J. 2004, 91, 239–248. [Google Scholar]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Totsky, I.V.; Lyakh, V.A. Pollen selection for drought tolerance in sunflower. Helia 2015, 38, 211–220. [Google Scholar] [CrossRef]
- Škorić, D. Sunflower breeding. In Sunflower Genetics and Breeding; Škorić, D., Seiler, G., Zhao, L., Jan, C.C., Miller, J.F., Charlet, L., Eds.; SASA-Branch: Novi Sad, Serbia, 2012; pp. 165–354. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Genotype | Vegetation Duration | Experimental Description |
|---|---|---|---|
| G1 | HA-267 (A × B) | Early | Production of Basic Seed Category The experiment consisted of two isolations to prevent the overlap of flowering between different B-analogs on earlier and later dates. In one isolation, SD1 and SD3 were sown, while in the other isolation, SD2 and SD4 were sown. All inbred lines had varying vegetation durations to avoid overlapping pollination of different B-analogous groups on the same SD. Across all SDs, the row ratio was 6:2 (A:B). |
| G2 | BG N 2 (PR) (A × B) | Medium-early | |
| G3 | IMI-AB-12 (PR) (A × B) | Late | |
| G4 | BG N 1 × SU RF 49 (A × Rf) | Medium-early | Production of First-Generation Certified Seed Category The experiment consisted of a single isolation encompassing all SDs. The male component (Rf) was the same across all three hybrid combinations. Across all SDs, the row ratio was 8:2 (A:Rf). |
| G5 | BG N 2 (PR) × SU RF 49 (A × Rf) | Medium-early | |
| G6 | BG N 4 × SU RF 49 (A × Rf) | Medium-early |
| Source of Variation | Traits | 2020 | 2021 | 2022 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| df | F | SS% | df | F | SS% | df | F | SS% | ||
| Sowing date (SD) Genotype (G) SD × G | HD | 3 | 49.32 ** | 42 | 3 | 45.25 ** | 27 | 3 | 10.60 ** | 10 |
| 5 | 12.66 ** | 19 | 5 | 41.26 ** | 41 | 5 | 27.13 ** | 40 | ||
| 15 | 9.23 ** | 39 | 15 | 10.65 ** | 32 | 15 | 11.33 ** | 50 | ||
| Sowing date (SD) Genotype (G) SD × G | PH | 3 | 29.61 ** | 7 | 3 | 110.48 ** | 32 | 3 | 29.33 ** | 12 |
| 5 | 204.52 ** | 75 | 5 | 89.26 ** | 43 | 5 | 110.20 ** | 74 | ||
| 15 | 16.44 ** | 18 | 15 | 17.361 ** | 25 | 15 | 6.89 ** | 14 | ||
| Sowing date (SD) Genotype (G) SD × G | GE | 3 | 5.61 ** | 6 | 3 | 24.71 ** | 7 | 3 | 116.94 ** | 17 |
| 5 | 42.34 ** | 67 | 5 | 155.05 ** | 76 | 5 | 172.45 ** | 42 | ||
| 15 | 5.71 ** | 27 | 15 | 11.59 ** | 17 | 15 | 55.10 ** | 41 | ||
| Sowing date (SD) Genotype (G) SD × G | GR | 3 | 3.22 * | 5 | 3 | 50.73 ** | 13 | 3 | 151.44 ** | 19 |
| 5 | 31.88 ** | 65 | 5 | 158.53 ** | 64 | 5 | 169.28 ** | 36 | ||
| 15 | 4.97 ** | 31 | 15 | 19.2 ** | 23 | 15 | 70.59 ** | 45 | ||
| Sowing date (SD) Genotype (G) SD × G | TSM | 3 | 22.14 ** | 24 | 3 | 20.17 ** | 11 | 3 | 38.02 ** | 7 |
| 5 | 25.96 ** | 47 | 5 | 64.71 ** | 63 | 5 | 228.61 ** | 66 | ||
| 15 | 5.35 ** | 29 | 15 | 8.83 ** | 26 | 15 | 31.12 ** | 27 | ||
| Sowing date (SD) Genotype (G) SD × G | Y | 3 | 10.04 ** | 5 | 3 | 11.62 ** | 6 | 3 | 36.24 ** | 6 |
| 5 | 53.13 ** | 42 | 5 | 95.89 ** | 74 | 5 | 261.40 ** | 75 | ||
| 15 | 22.05 ** | 53 | 15 | 8.72 ** | 20 | 15 | 21.55 ** | 19 | ||
| Sowing date (SD) Genotype (G) SD × G | OY | 3 | 13.66 ** | 7 | 3 | 5.44 ** | 3 | 3 | 52.92 ** | 8 |
| 5 | 45.18 ** | 39 | 5 | 86.83 ** | 76 | 5 | 269.29 ** | 71 | ||
| 15 | 20.73 ** | 54 | 15 | 7.96 ** | 21 | 15 | 27.08 ** | 21 | ||
| Factor | HD (cm) | PH (cm) | GE (%) | GR (%) | TSM (g) | Y (kg ha−1) | OY (kg ha−1) | |
|---|---|---|---|---|---|---|---|---|
| 2020 | ||||||||
| Sowing date (SD) | SD1 | 18.4 a | 128 b | 78 a | 82 ab | 65.4 a | 2045 a | 734 a |
| SD2 | 17.8 b | 128 b | 74 b | 79 b | 59.2 b | 1679 b | 565 b | |
| SD3 | 16.9 c | 129 b | 80 a | 85 a | 58.8 bc | 1766 b | 606 b | |
| SD4 | 16.0 d | 139 a | 79 a | 82 ab | 56.7 c | 1743 b | 605 b | |
| Genotype (G) | G1 | 18.2 a | 134 b | 60 e | 66 d | 66.5 a | 2279 a | 810 a |
| G2 | 17.5 bc | 118 cd | 73 d | 78 c | 55.2 c | 1329 c | 424 d | |
| G3 | 17.6 b | 163 a | 84 b | 89 ab | 60.6 b | 1323 c | 500 c | |
| G4 | 16.5 d | 136 b | 83 b | 86 b | 62.4 b | 2401 a | 827 a | |
| G5 | 16.8 d | 116 d | 89 a | 92 a | 62.0 b | 1745 b | 636 b | |
| G6 | 17.0 cd | 120 c | 78 c | 81 c | 53.1 c | 1773 b | 568 bc | |
| 2021 | ||||||||
| Sowing date (SD) | SD1 | 15.7 c | 92 d | 77 c | 81 c | 63.7 b | 1648 c | 637 b |
| SD2 | 15.4 c | 103 c | 82 b | 86 b | 62.3 bc | 1890 b | 705 a | |
| SD3 | 17.2 b | 110 b | 86 a | 91 a | 66.9 a | 1931 b | 710 a | |
| SD4 | 17.8 a | 116 a | 87 a | 91 a | 61.2 c | 2135 a | 767 a | |
| Genotype (G) | G1 | 14.6 d | 109 b | 58 c | 67 c | 71.1 a | 2136 b | 808 b |
| G2 | 17.9 a | 93 c | 81 b | 86 b | 60.6 d | 837 d | 302 e | |
| G3 | 15.7 c | 122 a | 82 b | 85 b | 62.5 c | 1631 c | 654 cd | |
| G4 | 16.7 b | 107 b | 93 a | 96 a | 65.4 b | 2976 a | 1106 a | |
| G5 | 18.1 a | 93 c | 91 a | 94 a | 66.2 b | 1745 c | 632 d | |
| G6 | 16.3 b | 109 b | 93 a | 95 a | 55.4 e | 2083 b | 725 c | |
| 2022 | ||||||||
| Sowing date (SD) | SD1 | 17.4 c | 115 a | 75 b | 83 b | 66.2 a | 1938 c | 713 c |
| SD2 | 18.4 ab | 114 a | 77 a | 80 a | 62.4 b | 2475 a | 930 a | |
| SD3 | 18.1 b | 108 b | 62 c | 68 d | 59.8 c | 2110 b | 783 b | |
| SD4 | 18.8 a | 103 c | 69 c | 73 c | 60.2 c | 1876 c | 653 d | |
| Genotype (G) | G1 | 16.2 d | 117 b | 60 e | 70 c | 66.0 bc | 2090 c | 744 d |
| G2 | 18.1 c | 94 d | 75 c | 81 b | 45.8 e | 844 e | 302 f | |
| G3 | 19.0 ab | 123 a | 60 e | 63 d | 59.1 d | 1398 d | 568 e | |
| G4 | 17.8 c | 123 a | 80 b | 83 b | 70.5 a | 3363 a | 1278 a | |
| G5 | 19.6 a | 93 d | 84 a | 87 a | 66.7 b | 2494 b | 916 b | |
| G6 | 18.4 bc | 111 c | 65 d | 71 c | 64.9 c | 2409 b | 810 c | |
| Average (GS) | 2020 | 17.3 | 131 | 78 | 82 | 60.0 | 1808 | 628 |
| 2021 | 16.5 | 105 | 83 | 87 | 63.5 | 1901 | 705 | |
| 2022 | 18.2 | 110 | 71 | 76 | 62.2 | 2100 | 770 | |
| CV% | 10.4 | 17.3 | 18.7 | 16.1 | 12.8 | 41.1 | 7.6 | |
| Standard deviation | 1.80 | 19.98 | 14.43 | 13.11 | 7.93 | 796.70 | 2.74 | |
| SD1 | SD2 | SD3 | SD4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DF | Eigenvalue | Cum Percent | Prob > F | Eigenvalue | Cum Percent | Prob > F | Eigenvalue | Cum Percent | Prob > F | Eigenvalue | Cum Percent | Prob > F |
| DF1 | 9.51 | 51.02 | <0.0001 * | 4.08 | 43.33 | <0.0001 * | 6.46 | 64.85 | <0.0001 * | 3.77 | 51.95 | <0.0001 * |
| DF2 | 6.18 | 84.17 | <0.0001 * | 2.64 | 71.33 | <0.0001 * | 2.29 | 87.84 | <0.0001 * | 2.29 | 83.52 | <0.0001 * |
| DF3 | 2.01 | 94.97 | <0.0001 * | 2.01 | 92.68 | <0.0001 * | 0.88 | 96.63 | <0.0001 * | 0.80 | 94.60 | <0.0001 * |
| DF4 | 0.91 | 99.85 | <0.0001 * | 0.68 | 99.93 | <0.0001 * | 0.32 | 99.88 | 0.0155 * | 0.35 | 99.46 | 0.005 * |
| DF5 | 0.03 | 100.00 | 0.63 | 0.01 | 100.00 | 0.94 | 0.01 | 100.00 | 0.8563 | 0.04 | 100.00 | 0.4829 |
| Test | <0.0001 * | <0.0001 * | <0.0001 * | <0.0001 * | ||||||||
| WL | <0.0001 * | <0.0001 * | <0.0001 * | <0.0001 * | ||||||||
| PT | <0.0001 * | <0.0001 * | <0.0001 * | <0.0001 * | ||||||||
| HL | <0.0001 * | <0.0001 * | <0.0001 * | <0.0001 * | ||||||||
| RMR | <0.0001 * | <0.0001 * | <0.0001 * | <0.0001 * |
| SD1 | HD | PH | GE | GR | TSM | Y | OY |
|---|---|---|---|---|---|---|---|
| DF1 | 0.14 | 0.79 | 1.01 | 0.10 | 0.21 | 4.52 | 4.86 |
| DF2 | 0.41 | 0.79 | 0.02 | 0.32 | 0.27 | 3.67 | 2.32 |
| DF3 | 0.17 | 0.50 | 0.28 | 0.11 | 0.17 | 3.19 | 3.44 |
| DF4 | 0.63 | 0.80 | 0.48 | 0.16 | 0.87 | 0.86 | 0.71 |
| DF5 | 0.46 | 0.32 | 1.89 | 1.91 | 0.11 | 1.06 | 1.09 |
| SD2 | HD | PH | GE | GR | TSM | Y | OY |
| DF1 | 1.00 | 0.76 | 1.84 | 0.97 | 0.25 | 4.14 | 4.30 |
| DF2 | 0.64 | 0.38 | 0.07 | 0.40 | 0.29 | 4.65 | 3.66 |
| DF3 | 0.03 | 0.82 | 0.41 | 0.05 | 0.09 | 4.89 | 5.13 |
| DF4 | 0.05 | 0.14 | 1.03 | 0.89 | 1.04 | 1.01 | 0.91 |
| DF5 | 0.29 | 0.01 | 2.15 | 2.33 | 0.04 | 0.61 | 0.93 |
| SD3 | HD | PH | GE | GR | TSM | Y | OY |
| DF1 | 0.35 | 1.30 | 0.27 | 0.56 | 0.64 | 4.93 | 5.21 |
| DF2 | 0.35 | 0.23 | 3.02 | 3.10 | 0.14 | 2.36 | 1.52 |
| DF3 | 0.23 | 0.04 | 2.34 | 1.99 | 0.24 | 2.46 | 2.08 |
| DF4 | 0.44 | 0.22 | 0.43 | 0.86 | 0.09 | 4.31 | 4.43 |
| DF5 | 0.24 | 0.18 | 0.41 | 0.00 | 0.94 | 0.74 | 1.30 |
| SD4 | HD | PH | GE | GR | TSM | Y | OY |
| DF1 | 0.37 | 0.08 | 1.21 | 1.28 | 0.49 | 3.03 | 2.33 |
| DF2 | 0.26 | 1.17 | 1.63 | 0.71 | 0.18 | 1.06 | 1.63 |
| DF3 | 0.16 | 0.26 | 1.66 | 0.96 | 0.77 | 2.31 | 1.94 |
| DF4 | 0.53 | 0.62 | 2.85 | 2.43 | 0.22 | 2.01 | 2.50 |
| DF5 | 0.24 | 0.07 | 2.84 | 3.04 | 0.47 | 2.16 | 2.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krstić, M.; Mladenov, V.; Banjac, B.; Babec, B.; Dunđerski, D.; Ćuk, N.; Gvozdenac, S.; Cvejić, S.; Jocić, S.; Miklič, V.; et al. Can Modification of Sowing Date and Genotype Selection Reduce the Impact of Climate Change on Sunflower Seed Production? Agriculture 2023, 13, 2149. https://doi.org/10.3390/agriculture13112149
Krstić M, Mladenov V, Banjac B, Babec B, Dunđerski D, Ćuk N, Gvozdenac S, Cvejić S, Jocić S, Miklič V, et al. Can Modification of Sowing Date and Genotype Selection Reduce the Impact of Climate Change on Sunflower Seed Production? Agriculture. 2023; 13(11):2149. https://doi.org/10.3390/agriculture13112149
Chicago/Turabian StyleKrstić, Miloš, Velimir Mladenov, Borislav Banjac, Brankica Babec, Dušan Dunđerski, Nemanja Ćuk, Sonja Gvozdenac, Sandra Cvejić, Siniša Jocić, Vladimir Miklič, and et al. 2023. "Can Modification of Sowing Date and Genotype Selection Reduce the Impact of Climate Change on Sunflower Seed Production?" Agriculture 13, no. 11: 2149. https://doi.org/10.3390/agriculture13112149
APA StyleKrstić, M., Mladenov, V., Banjac, B., Babec, B., Dunđerski, D., Ćuk, N., Gvozdenac, S., Cvejić, S., Jocić, S., Miklič, V., & Ovuka, J. (2023). Can Modification of Sowing Date and Genotype Selection Reduce the Impact of Climate Change on Sunflower Seed Production? Agriculture, 13(11), 2149. https://doi.org/10.3390/agriculture13112149