
Genetic Polymorphism and mRNA Expression Studies Reveal IL6R and LEPR Gene Associations with Reproductive Traits in Chinese Holsteins

 ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genotype and Haplotype-Based Association Analyses
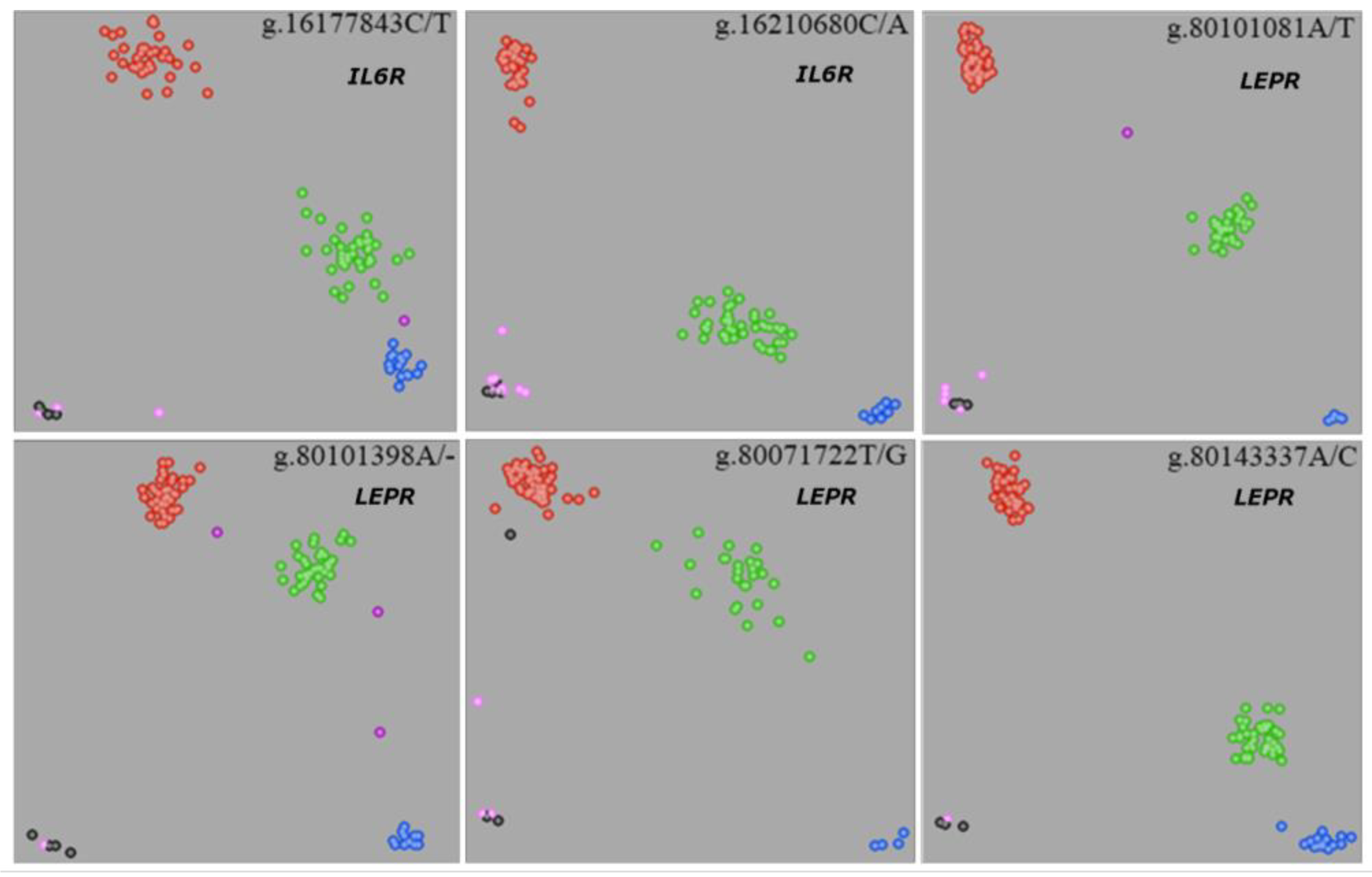
2.1.1. SNP Identification in 68 Bulls
2.1.2. Genotyping and Phenotyping of 1588 Cows
2.1.3. Association Analysis
2.2. Gene Expression Assay of IL6R and LEPR
2.2.1. Ovary Collection, Follicle Selection, and Granulosa Cell Isolation
2.2.2. Quantitative Reverse Transcription PCR (RT-qPCR)
3. Results
3.1. Screening of Polymorphisms in Association Popualtion
3.2. Association Analyses of SNPs with Reproductive Traits
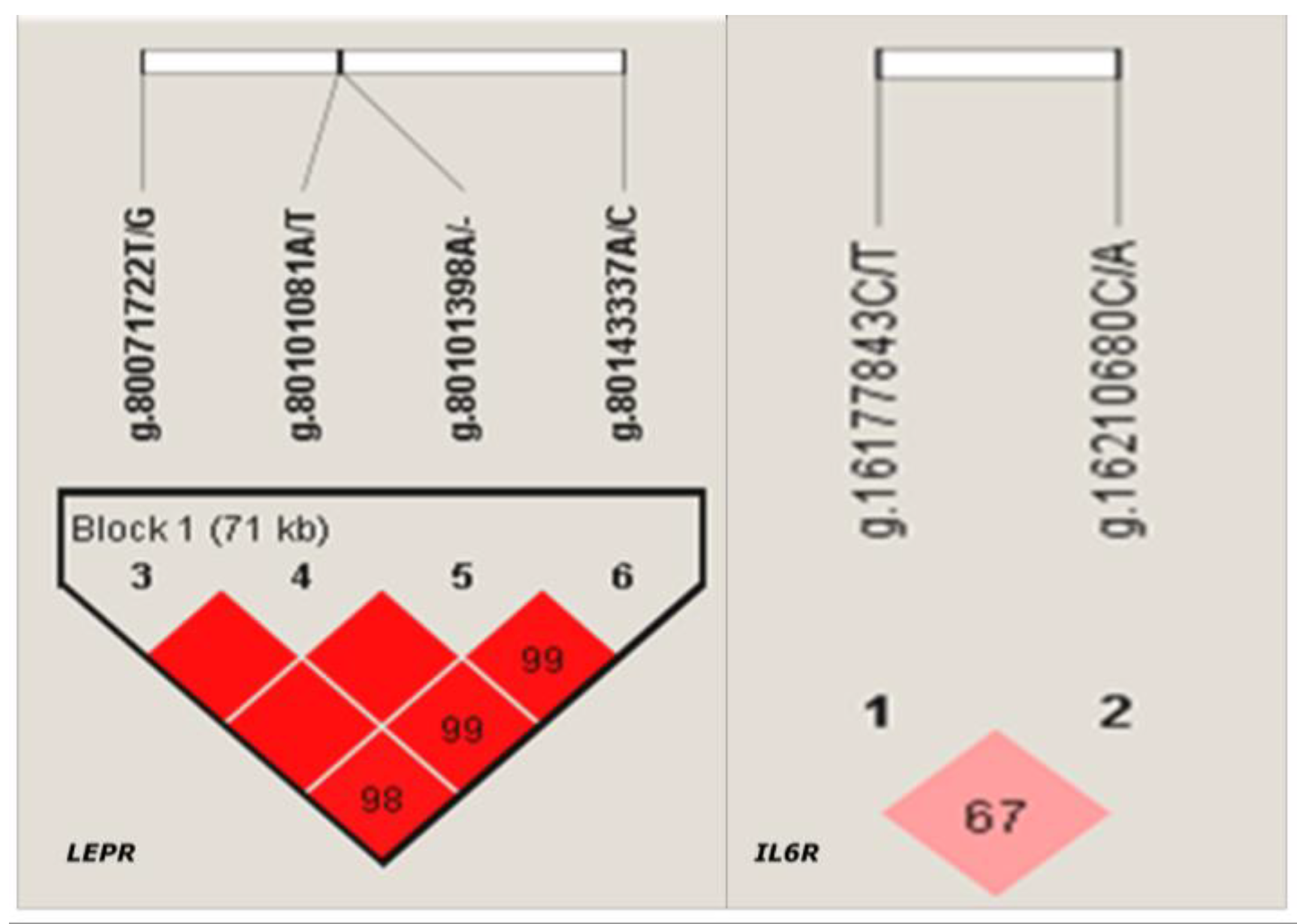
3.3. Association Analyses of Haplotype Blocks with Reproductive Traits
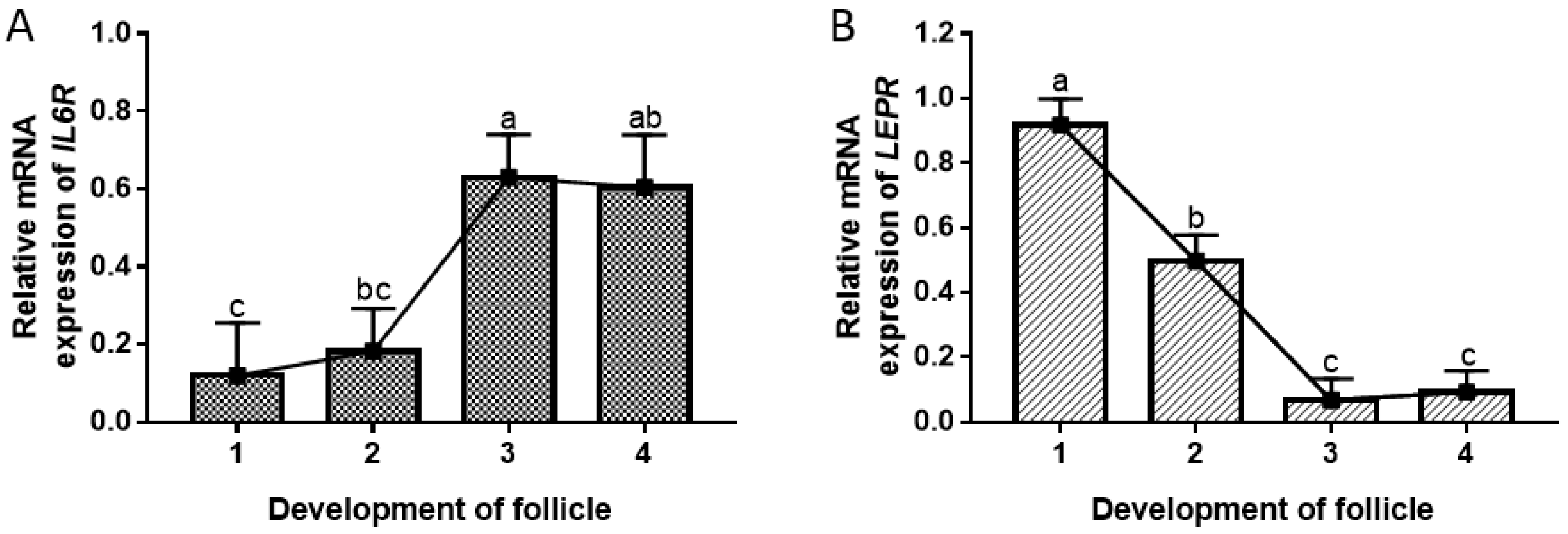
3.4. Gene Expression Analysis in GCs Isolated from Follicle of Different Developmental Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, S.W.; Williams, E.J.; Evans, A.C.O. A review of the causes of poor fertility in high milk producing dairy cows. Anim. Reprod. Sci. 2011, 123, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Jamrozik, J.; Fatehi, J.; Kistemaker, G.J.; Schaeffer, L.R. Estimates of genetic parameters for Canadian Holstein female reproduction traits. J. Dairy Sci. 2005, 88, 2199–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Guo, X.; Wang, Y.; Zhang, X.; Zhang, S.; Li, X.; Liu, L.; Shi, W.; Usman, T.; Wang, X.; et al. Estimation of genetic parameters of fertility traits in Chinese Holstein cattle. Can. J. Anim. Sci. 2014, 94, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Brondum, R.F.; Su, G.; Janss, L.; Sahana, G.; Guldbrandtsen, B.; Boichard, D.; Lund, M.S. Quantitative trait loci markers derived from whole genome sequence data increases the reliability of genomic prediction. J. Dairy Sci. 2015, 98, 4107–4116. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, A.; Cole, J.B.; VanRaden, P.M.; Wiggans, G.R.; Ruiz-López, F.J.; Van Tassell, C.P. Changes in genetic selection differentials and generation intervals in US Holstein dairy cattle as a result of genomic selection. Proc. Natl. Acad. Sci. USA 2016, 113, E3995–E4004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, C.G.; Gong, J.G.; Bramley, T.A.; Webb, R. Selection on predicted breeding value for milk production delays ovulation independently of changes in follicular development, milk production and body weight. Anim. Reprod. Sci. 2006, 95, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Sejian, V.; Bhatta, R.; Soren, N.M.; Malik, P.K.; Ravindra, J.P.; Prasad, C.S.; Lal, R. Introduction to Concepts of Climate Change Impact on Livestock and its Adaptation and Mitigation; Springer: New Delhi, India, 2015; pp. 1–23. [Google Scholar]
- Wathes, D.C.; Fenwick, M.; Cheng, Z.; Bourne, N.; Llewellyn, S.; Morris, D.G.; Kenny, D.; Murphy, J.; Fitzpatrick, R. Influence of negative energy balance on cyclicity and fertility in the high producing dairy cow. Theriogenology 2007, 68, S232–S241. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Umer, S.; Shi, R.; Zhu, H.; Zhao, X.; Wang, Y. Dairy cow reproduction under the influence of heat stress. J. Anim. Physiol. Anim. Nutr. 2020, 104, 978–986. [Google Scholar] [CrossRef]
- Ridker, P.M.; Pare, G.; Parker, A.; Zee, R.Y.; Danik, J.S.; Buring, J.E.; Kwiatkowski, D.; Cook, N.R.; Miletich, J.P.; Chasman, D.I. Loci related to metabolic-syndrome pathways including LEPR, HNF1A, IL6R, and GCKR associate with plasma C-reactive protein: The Women’s Genome Health Study. Am. J. Hum. Genet. 2008, 82, 1185–1192. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Wang, Y.; Sahana, G.; Zhang, Q.; Liu, L.; Lund, M.S.; Su, G. Genome-wide association studies for female fertility traits in Chinese and Nordic Holsteins. Sci. Rep. 2017, 7, 8487. [Google Scholar] [CrossRef]
- Van Snick, J. Interleukin-6: An overview. Annu. Rev. Immunol. 1990, 8, 253–278. [Google Scholar] [CrossRef]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Spicer, L.J. Leptin: A possible metabolic signal affecting reproduction. Domest. Anim. Endocrinol. 2001, 21, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Markert, U.R.; Morales-Prieto, D.M.; Fitzgerald, J.S. Understanding the link between the IL-6 cytokine family and pregnancy: Implications for future therapeutics. Expert Rev. Clin. Immunol. 2011, 7, 603–609. [Google Scholar] [CrossRef]
- Demir, B.; Guven, S.; Guven, E.S.; Atamer, Y.; Gul, T. Serum IL-6 level may have role in the pathophysiology of unexplained infertility. Am. J. Reprod. Immunol. 2009, 62, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, J.R.; Smith, S.K.; Wilson, A.; Sharkey, A.M. Soluble gp130 is up-regulated in the implantation window and shows altered secretion in patients with primary unexplained infertility. J. Clin. Endocrinol. Metab. 2002, 87, 3953–3960. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, H.; Liu, M.; Yuan, Y.; Wang, Z.; Chen, Y.; Wei, J.; Su, F.; Zhang, J. Treg/Th17 cell imbalance and IL-6 profile in patients with unexplained recurrent spontaneous abortion. Reprod. Sci. 2017, 24, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Benyo, D.F.; Smarason, A.; Redman, C.W.; Sims, C.; Conrad, K.P. Expression of inflammatory cytokines in placentas from women with preeclampsia. J. Clin. Endocrinol. Metab. 2001, 86, 2505–2512. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, M.; Rasti, Z. CTLA-4 and IL-6 gene polymorphisms: Risk factors for recurrent pregnancy loss. Hum. Immunol. 2016, 77, 1271–1274. [Google Scholar] [CrossRef]
- Medica, I.; Ostojic, S.; Pereza, N.; Kastrin, A.; Peterlin, B. Association between genetic polymorphisms in cytokine genes and recurrent miscarriage—A meta-analysis. Reprod. Biomed. Online 2009, 19, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Liefers, S.C.; Veerkamp, R.F.; Te Pas, M.F.W.; Chilliard, Y.; Van der Lende, T. Genetics and physiology of leptin in periparturient dairy cows. Domest. Anim. Endocrinol. 2005, 29, 227–238. [Google Scholar] [CrossRef]
- Chin, J.R.; Heuser, C.C.; Eller, A.G.; Branch, D.W.; Nelson, L.T.; Silver, R.M. Leptin and leptin receptor polymorphisms and recurrent pregnancy loss. J. Perinatol. 2013, 33, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Clempson, A.M.; Pollott, G.E.; Brickell, J.S.; Bourne, N.E.; Munce, N.; Wathes, D.C. Evidence that leptin genotype is associated with fertility, growth, and milk production in Holstein cows. J. Dairy Sci. 2011, 94, 3618–3628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.Q.; Wu, X.; O’Brien, M.J.; Pendola, F.L.; Denegre, J.N.; Matzuk, M.M.; Eppig, J.J. Synergistic roles of BMP15 and GDF9 in the development and function of the oocyte-cumulus cell complex in mice: Genetic evidence for an oocyte-granulosa cell regulatory loop. Dev. Biol. 2004, 276, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Petro, E.M.; Leroy, J.L.; Van Cruchten, S.J.; Covaci, A.; Jorssen, E.P.; Bols, P.E. Endocrine disruptors and female fertility: Focus on (bovine) ovarian follicular physiology. Theriogenology 2012, 78, 1887–1900. [Google Scholar] [CrossRef]
- Bettaieb, A.; Averill-Bates, D.A. Thermotolerance induced at a fever temperature of 40 degrees C protects cells against hyperthermia-induced apoptosis mediated by death receptor signalling. Biochem. Cell Biol. 2008, 86, 521–538. [Google Scholar] [CrossRef]
- Eppig, J.J. Oocyte control of ovarian follicular development and function in mammals. Reproduction 2001, 122, 829–838. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Holme, J.; Anthony, J. SNP Genotyping: The KASP Assay; Springer: New York, NY, USA, 2014; pp. 75–86. [Google Scholar]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettema, J.F.; Santos, J.E.P. Impact of age at calving on lactation, reproduction, health, and income in first-parity Holsteins on commercial farms. J. Dairy Sci. 2004, 87, 2730–2742. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Sargolzaei, M.; Abo-Ismail, M.K.; Miller, S.; Schenkel, F.; Moore, S.S.; Stothard, P. Genome-wide association study for lactation persistency, female fertility, longevity, and lifetime profit index traits in Holstein dairy cattle. J. Dairy Sci. 2017, 100, 1246–1258. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Lund, M.S.; Wang, Y.; Guo, G.; Dong, G.; Madsen, P.; Su, G. Variance components and correlations of female fertility traits in Chinese Holstein population. J. Anim. Sci. Biotechnol. 2017, 8, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, W.G.; Mackay, T.F. Falconer and Introduction to quantitative genetics. Genetics 2004, 167, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Vailati-Riboni, M.; Farina, G.; Batistel, F.; Heiser, A.; Mitchell, M.D.; Crookenden, M.A.; Walker, C.G.; Kay, J.K.; Meier, S.; Roche, J.R.; et al. Far-off and close-up dry matter intake modulate indicators of immunometabolic adaptations to lactation in subcutaneous adipose tissue of pasture-based transition dairy cows. J. Dairy Sci. 2017, 100, 2334–2350. [Google Scholar] [CrossRef]
- Mandour, M.A.B.; Al-Shami, S.A.; Al-Eknah, M.M. Body condition scores at calving and their association with dairy cow performance and health in semiarid environment under two cooling systems. Ital. J. Anim. Sci. 2015, 14, 2015–3690. [Google Scholar] [CrossRef] [Green Version]
- Trakovická, A.; Moravčíková, N.; Kasarda, R. Genetic polymorphisms of leptin and leptin receptor genes in relation with production and reproduction traits in cattle. Acta Biochim. Pol. 2013, 60, 783. [Google Scholar] [CrossRef] [Green Version]
- Asadollahpour, N.H.; Ansari, M.S.; Edriss, M.A. Effect of LEPR, ABCG2 and SCD1 gene polymorphisms on reproductive traits in the Iranian Holstein cattle. Reprod. Domest. Anim. 2014, 49, 769–774. [Google Scholar] [CrossRef]
- Baghenoey, S.G.; Mahyari, S.A.; Nanaei, H.A.; Rostami, M.; Edriss, M.A. Association of LEPR gene polymorphism with milk yield and age at first calving in the Iranian Holstein dairy cows. J. Livest. Sci. Technol. 2014, 2, 39–42. [Google Scholar]
- Hill, R.; Canal, A.; Bondioli, K.; Morell, R.; Garcia, M.D. Molecular markers located on the DGAT1, CAST, and LEPR genes and their associations with milk production and fertility traits in Holstein cattle. Genet. Mol. Res. 2016, 15, gmr.15017794. [Google Scholar] [CrossRef]
- Al-Janabi, H.R.A.; Al-Rawi, A.J.; Al-Anbari, N.N. Association of leptin receptor gene polymorphism with some productive and reproductive traits in Holstein primiparous cows. J. Entomol. Zool. Stud. 2018, 6, 1353–1358. [Google Scholar]
- Pandey, V. Association analyses of single nucleotide polymorphism in the leptin receptor gene with reproduction and production traits in high yielding Indian cow breed. J. Anim. Res. 2021, 11, 241–247. [Google Scholar] [CrossRef]
- Banos, G.; Woolliams, J.A.; Woodward, B.W.; Forbes, A.B.; Coffey, M.P. Impact of single nucleotide polymorphisms in leptin, leptin receptor, growth hormone receptor, and diacylglycerol acyltransferase (DGAT1) gene loci on milk Production, feed, and body energy traits of UK dairy cows. J. Dairy Sci. 2008, 91, 3190–3200. [Google Scholar] [CrossRef] [Green Version]
- Komisarek, J. Impact of LEP and LEPR gene polymorphisms on functional traits in Polish Holstein-Friesian cattle. Anim. Sci. Pap. Rep. 2010, 10, 133–141. [Google Scholar]
- Giblin, L.; Butler, S.T.; Kearney, B.M.; Waters, S.M.; Callanan, M.J.; Berry, D.P. Association of bovine leptin polymorphisms with energy output and energy storage traits in progeny tested Holstein-Friesian dairy cattle sires. BMC Genet. 2010, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Chen, H.; Lan, X.; Zhang, B.; Pan, C.; Zhang, L.; Zhang, C.; Zhao, M. Novel SNPs of the bovine LEPR gene and their association with growth traits. Biochem. Genet. 2008, 46, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.; Liu, G.Y.; Zhou, L.; Gui, L.S.; Khan, R.; Jinmeng, Y.; Chugang, M.; Schreurs, N.M.; Ji, R.; Zan, L. Detection of polymorphisms in the bovine leptin receptor gene affects fat deposition in two Chinese beef cattle breeds. Gene 2020, 758, 144957. [Google Scholar] [CrossRef] [PubMed]
- Samir, M.; Glister, C.; Mattar, D.; Laird, M.; Knight, P.G. Follicular expression of pro-inflammatory cytokines tumour necrosis factor-α (TNFα), interleukin 6 (IL6) and their receptors in cattle: TNFα, IL6 and macrophages suppress thecal androgen production in vitro. Reproduction 2017, 154, 35–49. [Google Scholar] [CrossRef]
- Baskind, N.E.; Orsi, N.M.; Sharma, V. Follicular-phase ovarian follicular fluid and plasma cytokine profiling of natural cycle in vitro fertilization patients. Fertil. Steril. 2014, 102, 410–418. [Google Scholar] [CrossRef]
- Salmassi, A.; Lu, S.; Hedderich, J.; Oettinghaus, C.; Jonat, W.; Mettler, L. Interaction of interleukin-6 on human granulosa cell steroid secretion. J. Endocrinol. 2001, 170, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Liu, Y.; Yao, W.; Li, Q.; Liu, H.; Pan, Z. Initiation of follicular atresia: Gene networks during early atresia in pig ovaries. Reproduction 2018, 156, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, M.; Schilffarth, S.; Schams, D.; Meyer, H.H.; Berisha, B. The expression of leptin and its receptor during different physiological stages in the bovine ovary. Mol. Reprod. Dev. 2010, 77, 174–181. [Google Scholar] [CrossRef]
- Boelhauve, M.; Sinowatz, F.; Wolf, E.; Paula-Lopes, F.F. Maturation of bovine oocytes in the presence of leptin improves development and reduces apoptosis of in vitro-produced blastocysts. Biol. Reprod. 2005, 73, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | SNP | Genotype | Number of Individuals | Genotypic Frequency | Allele | Allelic Frequency | H-W-E Test, χ2 Value (df = 1) |
|---|---|---|---|---|---|---|---|
| G:G | 155 | 0.125 | G | 0.373 | |||
| IL6R | g.16210680C/A | T:G | 614 | 0.496 | T | 0.627 | 0.034 |
| T:T | 469 | 0.379 | |||||
| A:A | 455 | 0.325 | A | 0.570 | |||
| IL6R | g.16177843C/T | A:G | 684 | 0.489 | G | 0.430 | 0.945 |
| G:G | 259 | 0.185 | |||||
| G:G | 439 | 0.299 | G | 0.550 | |||
| LEPR | g.80143337A/C | G:T | 736 | 0.502 | T | 0.450 | 0.581 |
| T:T | 291 | 0.198 | |||||
| -:- | 125 | 0.085 | - | 0.297 | |||
| LEPR | g.80101398A/- | A:- | 621 | 0.423 | A | 0.703 | 0.600 |
| A:A | 722 | 0.492 | |||||
| A:A | 53 | 0.037 | A | 0.177 | |||
| LEPR | g.80071722T/G | C:A | 403 | 0.280 | C | 0.823 | 0.145 |
| C:C | 985 | 0.684 | |||||
| A:A | 721 | 0.494 | A | 0.706 | |||
| LEPR | g.80101081A/T | A:T | 619 | 0.424 | T | 0.294 | 0.387 |
| T:T | 119 | 0.082 |
| SNP | Genotype | AFS | AFC | IFL_H | IFL_C | ICF | SB_H | SB_C | CE_H | CE_C |
|---|---|---|---|---|---|---|---|---|---|---|
| IL6R | GG (155) | −16.718 ± 0.631 | −10.085 ± 0.506 | 5.983 ± 0.499 | 2.365 ± 0.410 | 0.544 ± 0.090 | 0.025 ± 0.003 | 0.016 ± 0.001 | 0.009 ± 0.001 | 0.001 ± 0.000 |
| g.16210680C/A | TG (614) | −17.431 ± 0.317 | −9.317 ± 0.254 | 6.609 ± 0.251 | 2.585 ± 0.206 | 0.425 ± 0.045 | 0.029 ± 0.001 | 0.015 ± 0.001 | 0.009 ± 0.001 | 0.001 ± 0.000 |
| TT (469) | −17.359 ± 0.363 | −9.020 ± 0.291 | 6.673 ± 0.287 | 2.475 ± 0.236 | 0.356 ± 0.052 | 0.028 ± 0.001 | 0.014 ± 0.001 | 0.008 ± 0.001 | 0.001 ± 0.000 | |
| p value | 0.593 | 0.189 | 0.466 | 0.870 | 0.182 | 0.450 | 0.089 | 0.331 | 0.162 | |
| IL6R | AA (456) | −17.750 ± 0.354 | −9.726 ± 0.299 | 6.464 ± 0.295 | 2.505 ± 0.237 | 0.403 ± 0.052 | 0.027 ± 0.001 | 0.014 ± 0.001 | 0.007 ± 0.001 b | 0.001 ± 0.000 |
| g.16177843C/T | AG (684) | −17.392 ± 0.289 | −9.129 ± 0.244 | 6.921 ± 0.241 | 2.465 ± 0.193 | 0.355 ± 0.043 | 0.029 ± 0.001 | 0.015 ± 0.000 | 0.009 ± 0.000 a | 0.001 ± 0.000 |
| GG (260) | −17.199 ± 0.469 | −8.731 ± 0.397 | 6.739 ± 0.391 | 2.344 ± 0.314 | 0.469 ± 0.070 | 0.027 ± 0.002 | 0.014 ± 0.001 | 0.009 ± 0.001 ab | 0.000 ± 0.000 | |
| p value | 0.598 | 0.107 | 0.486 | 0.917 | 0.367 | 0.495 | 0.690 | 0.018 | 0.310 | |
| LEPR | GG (440) | −16.878 ± 0.370 | −8.753 ± 0.306a | 7.114 ± 0.299 | 2.666 ± 0.243 | 0.571 ± 0.053a | 0.028 ± 0.001 | 0.015 ± 0.001 | 0.007 ± 0.001 b | 0.001 ± 0.000 a |
| g.80143337A/C | GT (737) | −17.978 ± 0.285 | −9.694 ± 0.236b | 6.414 ± 0.231 | 2.605 ± 0.188 | 0.394 ± 0.041b | 0.029 ± 0.001 | 0.015 ± 0.000 | 0.009 ± 0.000 a | 0.001 ± 0.000 a |
| TT (291) | −17.232 ± 0.454 | −8.942 ± 0.375ab | 6.609 ± 0.367 | 2.260 ± 0.299 | 0.076 ± 0.065c | 0.027 ± 0.002 | 0.015 ± 0.001 | 0.010 ± 0.001 a | 0.000 ± 0.000 b | |
| p value | 0.050 | 0.033 | 0.179 | 0.534 | <0.0001 | 0.794 | 0.897 | 0.002 | <0.0001 | |
| LEPR | 119 | −16.770 ± 0.707 | −8.300 ± 0.590 | 7.485 ± 0.575 | 2.950 ± 0.469 | 0.641 ± 0.103a | 0.030 ± 0.003 | 0.014 ± 0.001 | 0.007 ± 0.001 | 0.001 ± 0.001 |
| g.80101398A/- | A- (621) | −17.610 ± 0.308 | −9.344 ± 0.257 | 6.690 ± 0.250 | 2.701 ± 0.204 | 0.437 ± 0.045 ab | 0.028 ± 0.001 | 0.014 ± 0.001 | 0.008 ± 0.001 | 0.001 ± 0.000 |
| AA (723) | −17.570 ± 0.286 | −9.305 ± 0.239 | 6.547 ± 0.232 | 2.281 ± 0.189 | 0.294 ± 0.041 b | 0.028 ± 0.001 | 0.015 ± 0.000 | 0.009 ± 0.000 | 0.001 ± 0.000 | |
| p value | 0.538 | 0.250 | 0.318 | 0.201 | 0.002 | 0.850 | 0.354 | 0.388 | 0.510 | |
| LEPR | AA (722) | −17.649 ± 0.286 | −9.332 ± 0.239 | 6.522 ± 0.233 | 2.277 ± 0.190 | 0.290 ± 0.042 b | 0.028 ± 0.001 | 0.015 ± 0.000 | 0.009 ± 0.000 | 0.001 ± 0.000 |
| g.80101081A/T | AT (619) | −17.598 ± 0.308 | −9.328 ± 0.258 | 6.723 ± 0.251 | 2.739 ± 0.205 | 0.434 ± 0.045 ab | 0.028 ± 0.001 | 0.014 ± 0.001 | 0.008 ± 0.001 | 0.001 ± 0.000 |
| TT (120) | −16.613 ± 0.703 | −8.270 ± 0.588 | 7.291 ± 0.574 | 2.854 ± 0.467 | 0.633 ± 0.103 a | 0.030 ± 0.003 | 0.015 ± 0.001 | 0.007 ± 0.001 | 0.001 ± 0.001 | |
| p value | 0.383 | 0.225 | 0.445 | 0.192 | 0.002 | 0.741 | 0.334 | 0.366 | 0.470 | |
| LEPR | AA (53) | −18.391 ± 1.060 | −9.591 ± 0.872 | 5.843 ± 0.852 | 2.571 ± 0.702 | 0.520 ± 0.154 ab | 0.029 ± 0.004 | 0.017 ± 0.002 | 0.006 ± 0.002 | 0.002 ± 0.001 |
| g.80071722T/G | CA (404) | −16.865 ± 0.385 | −8.618 ± 0.316 | 6.833 ± 0.309 | 2.911 ± 0.254 | 0.517 ± 0.056 a | 0.028 ± 0.002 | 0.015 ± 0.001 | 0.008 ± 0.001 | 0.001 ± 0.000 |
| CC (986) | −17.799 ± 0.246 | −9.515 ± 0.202 | 6.641 ± 0.198 | 2.375 ± 0.163 | 0.335 ± 0.036 b | 0.028 ± 0.001 | 0.015 ± 0.000 | 0.009 ± 0.000 | 0.001 ± 0.000 | |
| p value | 0.090 | 0.054 | 0.537 | 0.207 | 0.017 | 0.969 | 0.448 | 0.168 | 0.336 |
| Haplotype | AFS | AFC | IFL_H | IFL_C | ICF | SB_H | SB_C | CE_H | CE_C |
|---|---|---|---|---|---|---|---|---|---|
| H1H1 (274) | −17.304 ± 0.462 | −9.194 ± 0.382 | 6.493 ± 0.376 | 2.178 ± 0.308 | 0.091 ± 0.067 b | 0.027 ± 0.002 | 0.015 ± 0.001 | 0.010 ± 0.001 | 0.000 ± 0.000 b |
| H1H2 (379) | −18.234 ± 0.393 | −9.720 ± 0.325 | 6.522 ± 0.320 | 2.886 ± 0.262 | 0.396 ± 0.057 a | 0.028 ± 0.002 | 0.014 ± 0.001 | 0.009 ± 0.001 | 0.000 ± 0.000 ab |
| H1H3 (215) | −17.282 ± 0.522 | −8.786 ± 0.431 | 6.588 ± 0.425 | 2.773 ± 0.348 | 0.449 ± 0.076 a | 0.029 ± 0.002 | 0.015 ± 0.001 | 0.008 ± 0.001 | 0.000 ± 0.000 ab |
| H1H4 (108) | −18.748 ± 0.736 | −10.791 ± 0.609 | 6.223 ± 0.600 | 1.189 ± 0.491 | 0.270 ± 0.107 ab | 0.029 ± 0.003 | 0.016 ± 0.001 | 0.010 ± 0.001 | 0.002 ± 0.001 a |
| H2H2 (115) | −16.655 ± 0.713 | −8.191 ± 0.590 | 7.465 ± 0.581 | 2.923 ± 0.475 | 0.652 ± 0.103 a | 0.030 ± 0.003 | 0.015 ± 0.001 | 0.007 ± 0.001 | 0.001 ± 0.001 ab |
| H2H3 (144) | −16.422 ± 0.637 | −8.239 ± 0.527 | 7.306 ± 0.519 | 3.264 ± 0.425 | 0.578 ± 0.092 a | 0.028 ± 0.003 | 0.014 ± 0.001 | 0.007 ± 0.001 | 0.001 ± 0.000 ab |
| H2H4 (62) | −16.877 ± 0.971 | −9.634 ± 0.803 | 7.058 ± 0.791 | 1.188 ± 0.648 | 0.474 ± 0.141 ab | 0.027 ± 0.004 | 0.014 ± 0.002 | 0.006 ± 0.002 | 0.002 ± 0.001 ab |
| H3H3 (52) | −18.382 ± 1.061 | −9.591 ± 0.877 | 5.872 ± 0.864 | 2.644 ± 0.707 | 0.526 ± 0.154 ab | 0.029 ± 0.004 | 0.017 ± 0.002 | 0.006 ± 0.002 | 0.002 ± 0.001 ab |
| H3H4 (29) | −17.430 ± 1.420 | −9.808 ± 1.175 | 6.159 ± 1.157 | 3.277 ± 0.947 | 0.457 ± 0.206 ab | 0.028 ± 0.006 | 0.015 ± 0.002 | 0.008 ± 0.002 | 0.002 ± 0.001 ab |
| H4H4 (9) | −23.454 ± 2.550 | −12.237 ± 2.108 | 9.755 ± 2.077 | 0.389 ± 1.700 | 0.568 ± 0.370 ab | 0.013 ± 0.010 | 0.010 ± 0.004 | 0.001 ± 0.004 | 0.001 ± 0.002 ab |
| p value | 0.044 | 0.025 | 0.545 | 0.012 | <0.001 | 0.942 | 0.850 | 0.087 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Sammad, A.; Shi, R.; Dong, Y.; Zhao, S.; Liu, L.; Guo, G.; Xu, Q.; Liu, A.; Wang, Y. Genetic Polymorphism and mRNA Expression Studies Reveal IL6R and LEPR Gene Associations with Reproductive Traits in Chinese Holsteins. Agriculture 2023, 13, 321. https://doi.org/10.3390/agriculture13020321
Zhang H, Sammad A, Shi R, Dong Y, Zhao S, Liu L, Guo G, Xu Q, Liu A, Wang Y. Genetic Polymorphism and mRNA Expression Studies Reveal IL6R and LEPR Gene Associations with Reproductive Traits in Chinese Holsteins. Agriculture. 2023; 13(2):321. https://doi.org/10.3390/agriculture13020321
Chicago/Turabian StyleZhang, Hailiang, Abdul Sammad, Rui Shi, Yixin Dong, Shanjiang Zhao, Lin Liu, Gang Guo, Qing Xu, Aoxing Liu, and Yachun Wang. 2023. "Genetic Polymorphism and mRNA Expression Studies Reveal IL6R and LEPR Gene Associations with Reproductive Traits in Chinese Holsteins" Agriculture 13, no. 2: 321. https://doi.org/10.3390/agriculture13020321
APA StyleZhang, H., Sammad, A., Shi, R., Dong, Y., Zhao, S., Liu, L., Guo, G., Xu, Q., Liu, A., & Wang, Y. (2023). Genetic Polymorphism and mRNA Expression Studies Reveal IL6R and LEPR Gene Associations with Reproductive Traits in Chinese Holsteins. Agriculture, 13(2), 321. https://doi.org/10.3390/agriculture13020321